
Mineral Paragenesis Precipitating in Salt Flat Pools of Continental Environments Replicated in Microbial Mat Microcosms without Evaporation



Abstract
:1. Introduction
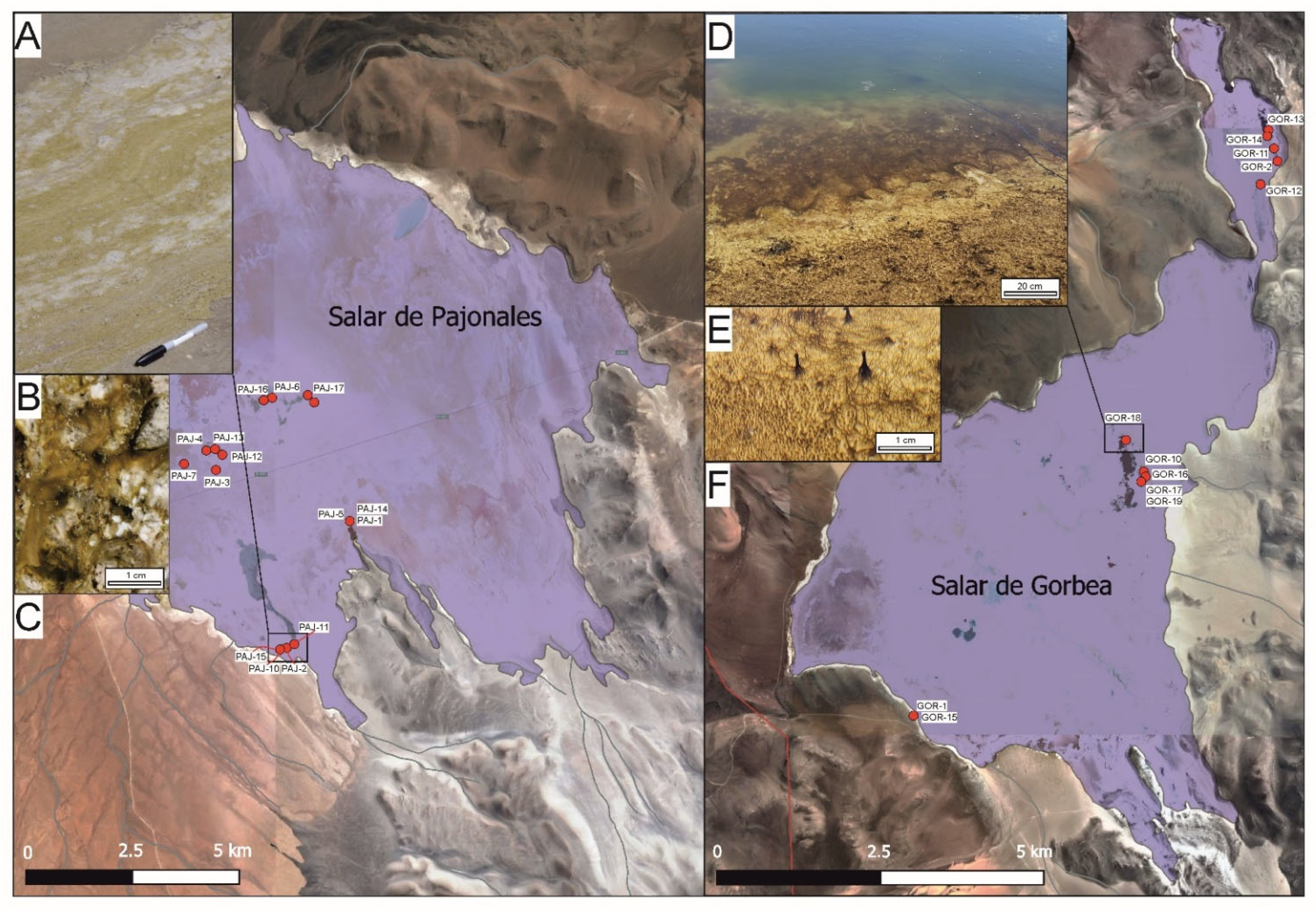
Overview of Sites
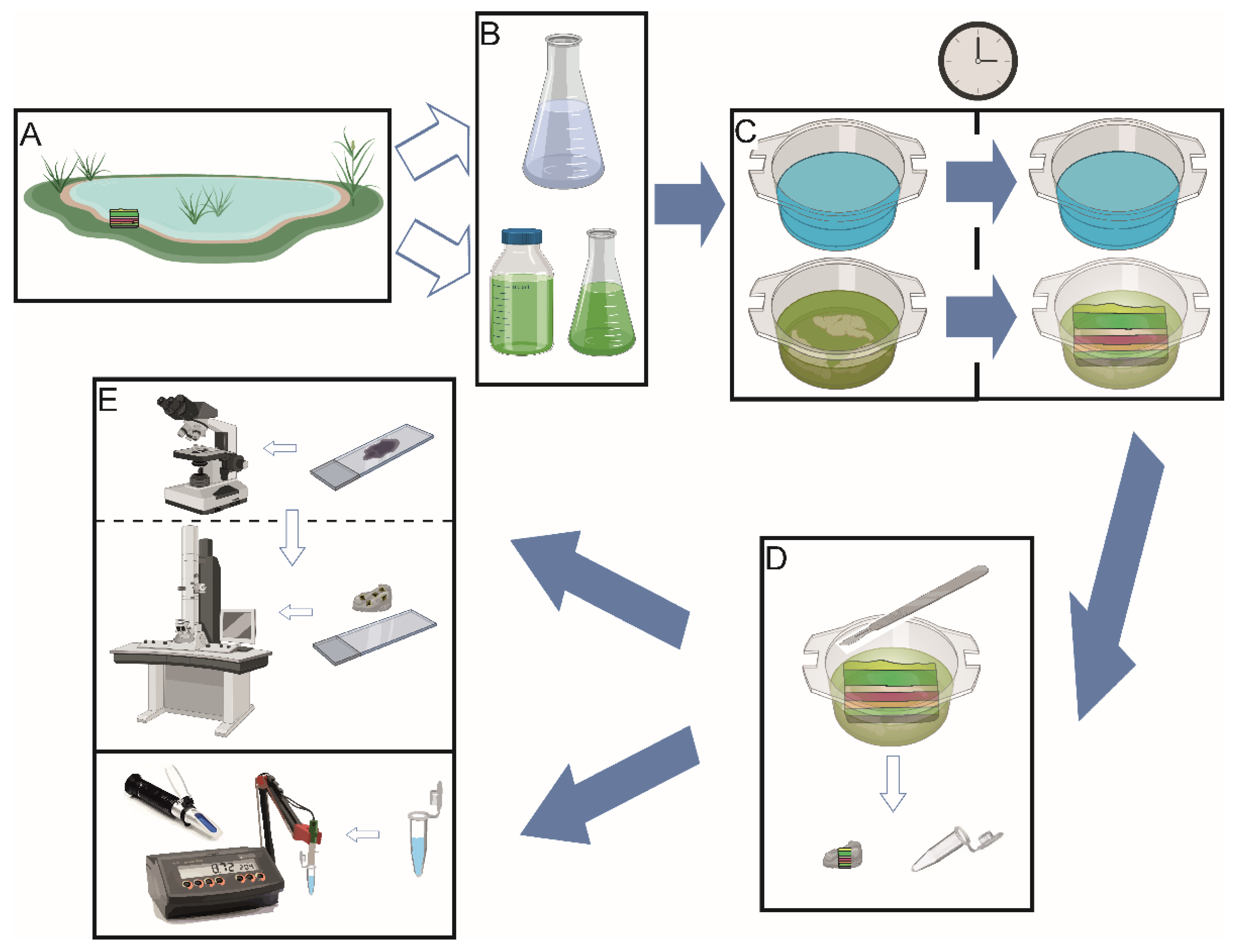
2. Methodology
2.1. Field Work, Sample Collection, and Water Analysis
2.2. Experimental Setup: Intact Microbial Mat Manipulations
2.3. Optical and Electron Microscopy
2.4. X-ray Diffraction
2.5. Data Processing
3. Results
3.1. Field Measurements and Geochemical Modeling
3.2. Physicochemical Monitoring of Laboratory Incubations
3.3. Microbial Mat Dynamics
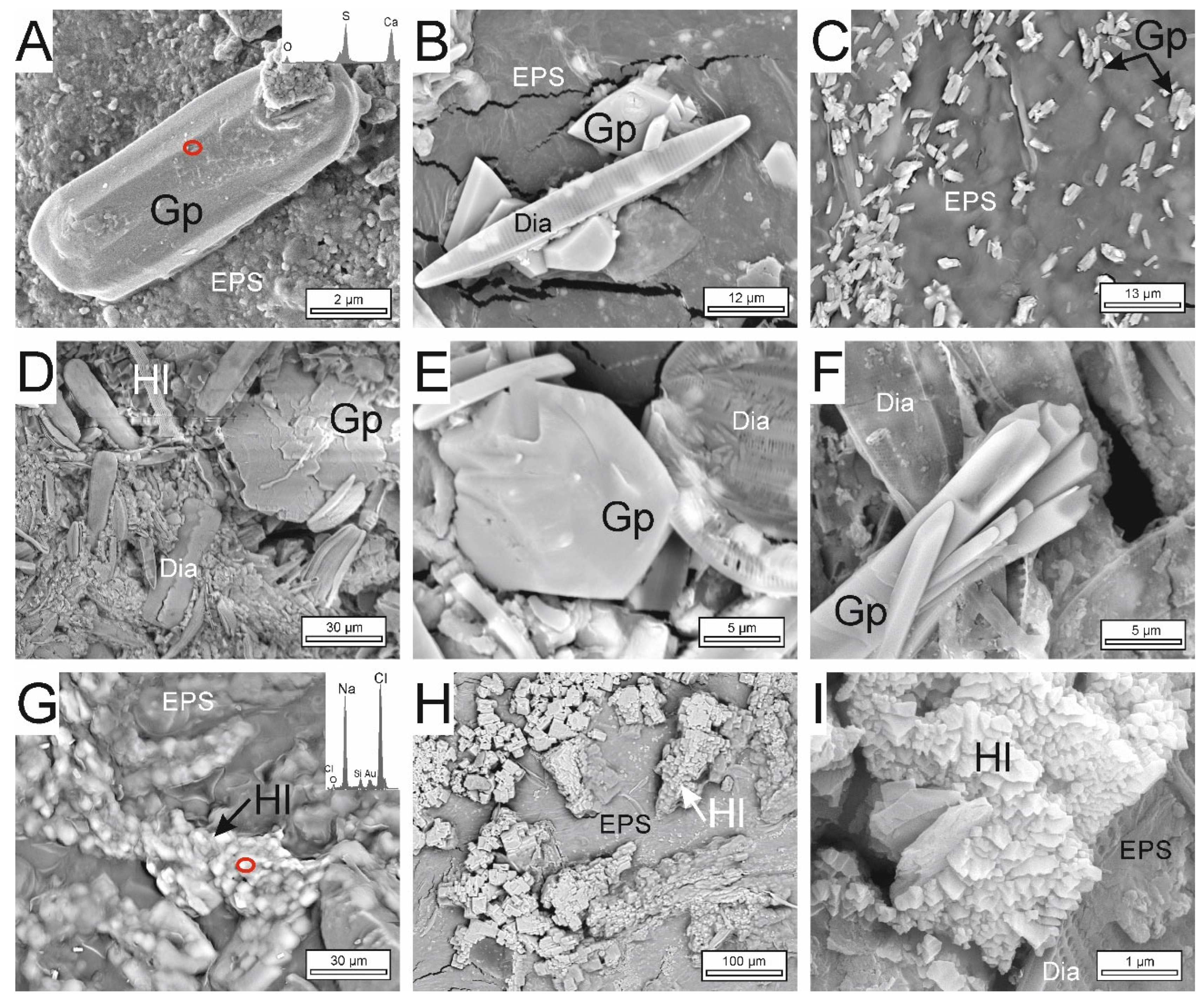
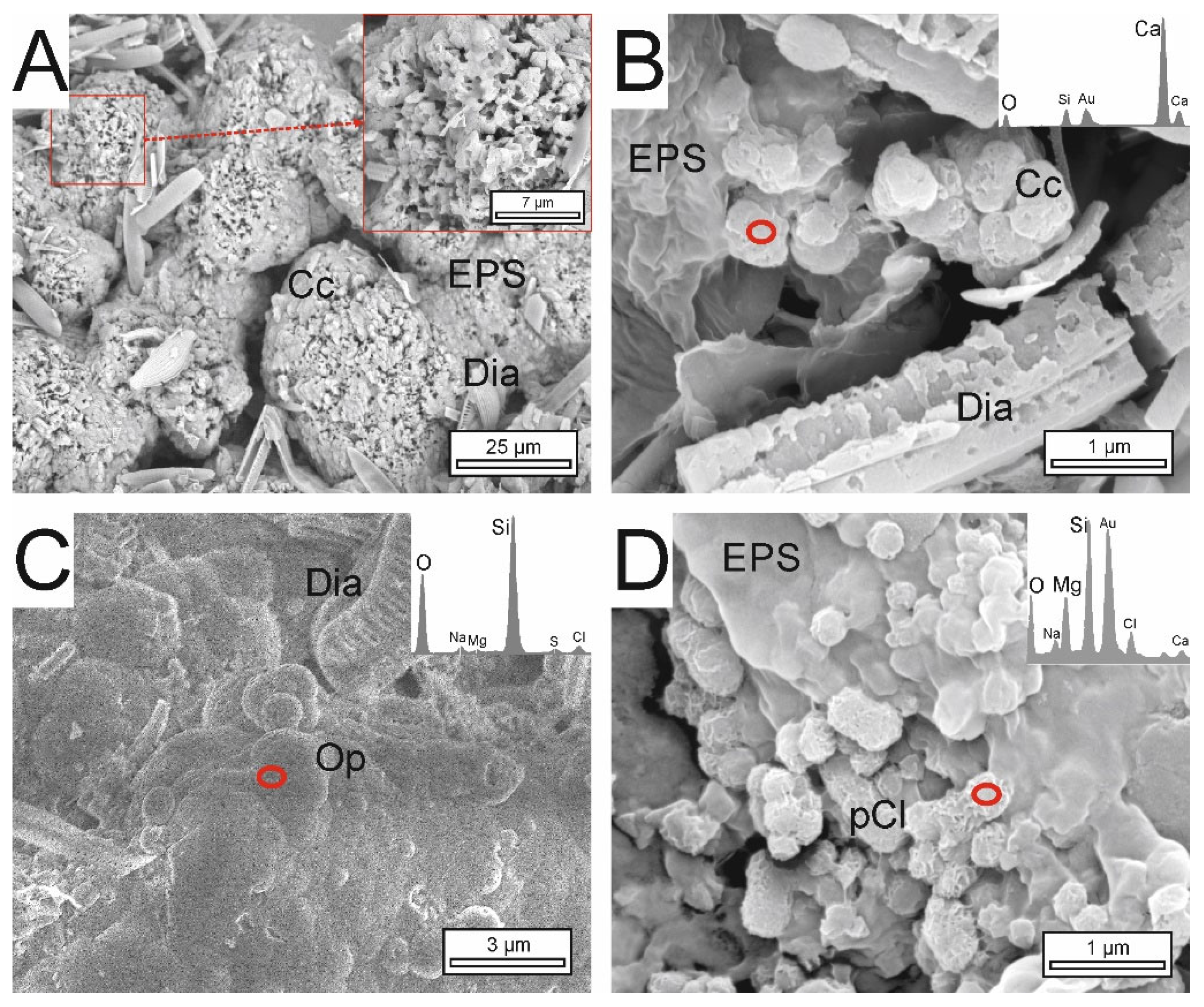
3.4. Textural and Mineralogical Composition of Precipitates
4. Discussion
4.1. Substrates Used in Microcosms
4.2. Monitoring Communities: Short-Term and Long-Term Mineral Formation Implications
4.3. Textures for the Interpretation of Recent and Preserved Microbialites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eugster, H.P.; Hardie, L.A. Saline Lakes. In Physics and Chemistry of Lakes; Lerman, A., Ed.; Springer: Berlin/Heidelberg, Germany, 1978; pp. 237–293. [Google Scholar]
- Warren, J.K. Evaporites: Sediments, Sources and Hydrocarbons; Springer: Berlin, Germany, 2006; 1035p. [Google Scholar]
- Van Driessche AE, S.; Stawski, T.M.; Kellermeier, M. Calcium sulfate precipitation pathways in natural and engineered environments. Chem. Geol. 2019, 530, 119274. [Google Scholar] [CrossRef]
- Cabrol, N.A.; McKay, C.P.; Grin, E.A.; Kiss, K.T.; Ács, É.; Tóth, B.; Grigorszky, I.; Szabó-Taylor, K.; Fike, D.A.; Hock, A.N.; et al. Signatures of habitats and life in Earth’s high-altitude lakes: Clues to Noachian aqueous environments on Mars. In The Geology of Mars: Evidence from Earth-Based Analogs; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Newman, D.K.; Banfield, J.F. Geomicrobiology: How molecular-scale interactions underpin biogeochemical systems. Science 2002, 296, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Montague, C.L. Influence of biota on erodibility of sediments. In Estuarine Cohesive Sediment Dynamics; Springer: New York, NY, USA, 1986; pp. 251–269. [Google Scholar]
- Dupraz, C.; Visscher, P.T. Microbial lithification in marine stromatolites and hypersaline mats. Trends Microbiol. 2005, 13, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.E.; Grant, W.D.; Duckworth, A.W.; Owenson, G.G. Microbial diversity of soda lakes. Extremophiles 1998, 2, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Burne, R.V.; Moore, L.S. Microbialites; organosedimentary deposits of benthic microbial communities. Palaios 1987, 2, 241–254. [Google Scholar] [CrossRef]
- Kobluk, D.R.; Crawford, D.R. A modern hypersaline organic mud-and gypsum-dominated basin and associated microbialites. Palaios 1990, 5, 134–148. [Google Scholar] [CrossRef]
- Vogel, M.B.; Des Marais, D.J.; Parenteau, M.N.; Jahnke, L.L.; Turk, K.A.; Kubo, M.D. Biological influences on modern sulfates: Textures and composition of gypsum deposits from Guerrero Negro, Baja California Sur, Mexico. Sediment. Geol. 2010, 223, 265–280. [Google Scholar] [CrossRef]
- Petrash, D.A.; Gingras, M.K.; Lalonde, S.V.; Orange, F.; Pecoits, E.; Konhauser, K.O. Dynamic controls on accretion and lithification of modern gypsum-dominated thrombolites, Los Roques, Venezuela. Sediment. Geol. 2012, 245, 29–47. [Google Scholar] [CrossRef]
- Taher, A.G. Microbially induced sedimentary structures in evaporite–siliciclastic sediments of Ras Gemsa sabkha, Red Sea Coast, Egypt. J. Adv. Res. 2014, 5, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Strohmenger, C.J.; Jameson, J. Gypsum stromatolites from Sawda Nathil: Relicts from a southern coastline of Qatar. Carbonates Evaporites 2018, 33, 169–186. [Google Scholar] [CrossRef]
- Cabestrero, Ó.; Sanz-Montero, M.E. Brine evolution in two inland evaporative environments: Influence of microbial mats in mineral precipitation. J. Paleolimnol. 2016, 59, 139–157. [Google Scholar] [CrossRef]
- Cabestrero, Ó.; del Buey, P.; Sanz-Montero, M.E. Biosedimentary and geochemical constraints on the precipitation of mineral crusts in shallow sulphate lakes. Sediment. Geol. 2018, 366, 32–46. [Google Scholar] [CrossRef]
- Cabestrero, Ó.; Sanz-Montero, M.E.; Arregui, L.; Serrano, S.; Visscher, P.T. Seasonal variability of mineral formation in microbial mats subjected to drying and wetting cycles in alkaline and hypersaline sedimentary environments. Aquat. Geochem. 2018, 24, 79–105. [Google Scholar] [CrossRef]
- del Buey, P.; Cabestrero, Ó.; Arroyo, X.; Sanz-Montero, M.E. Microbially induced palygorskite-sepiolite authigenesis in modern hypersaline lakes (Central Spain). Appl. Clay Sci. 2018, 160, 9–21. [Google Scholar] [CrossRef]
- del Buey, P.; Sanz-Montero, M.E.; Braissant, O.; Cabestrero, Ó.; Visscher, P.T. The role of microbial extracellular polymeric substances on formation of sulfate minerals and fibrous Mg-clays. Chem. Geol. 2021, 581, 120403. [Google Scholar] [CrossRef]
- Demergasso, C.; Chong, G.; Galleguillos, P.; Escudero, L.; Martínez-Alonso, M.; Esteve, I. Tapetes microbianos del Salar de Llamará, norte de Chile. Rev. Chil. Hist. Nat. 2003, 76, 485–499. [Google Scholar] [CrossRef] [Green Version]
- Farias, M.E.; Contreras, M.; Rasuk, M.C.; Kurth, D.; Flores, M.R.; Poire, D.G.; Novoa, F.; Visscher, P.T. Characterization of bacterial diversity associated with microbial mats, gypsum evaporites and carbonate microbialites in thalassic wetlands: Tebenquiche and La Brava, Salar de Atacama, Chile. Extremophiles 2014, 18, 311–329. [Google Scholar] [CrossRef]
- Rasuk, M.C.; Kurth, D.; Flores, M.R.; Contreras, M.; Novoa, F.; Poire, D.; Farias, M.E. Microbial characterization of microbial ecosystems associated to evaporites domes of gypsum in Salar de Llamara in Atacama Desert. Microb. Ecol. 2014, 68, 483–494. [Google Scholar] [CrossRef]
- Ercilla Herrero, O. Origen y evolución de estromatolitos de yeso en salares del altiplano andino, norte de Chile. Andean Geol. 2019, 46, 211–222. [Google Scholar]
- Madigan, M.; Bender, K.; Buckley, D.; Sattley, W.M.; Stahl, D. Brock Biology of Microorganisms, 15th ed.; Pearson: New York, NY, USA, 2018; 1064p. [Google Scholar]
- Thompson, J.B.; Ferris, F.G. Cyanobacterial precipitation of gypsum, calcite, and magnesite from natural alkaline lake water. Geology 1990, 18, 995–998. [Google Scholar] [CrossRef]
- Demergasso, C.; Chong, G.; Escudero, L.; Pueyo, J.J.; Pedrós-Alió, C. Microbial Precipitation of Arsenic Sulfides in Andean Salt Flats. Geomicrobiol. J. 2007, 24, 111–123. [Google Scholar] [CrossRef]
- Sánchez-Román, M.; Vasconcelos, C.; Warthmann, R.; Rivadeneyra, M.; McKenzie, J.A. Microbial dolomite precipitation under aerobic conditions: Results from Brejo do Espinho Lagoon (Brazil) and culture experiments. Int. Assoc. Sedimentol. Spec. Publ. 2009, 41, 167–178. [Google Scholar]
- Glunk, C.; Dupraz, C.; Braissant, O.; Gallagher, K.L.; Verrecchia, E.P.; Visscher, P.T. Microbially mediated carbonate precipitation in a hypersaline lake, Big Pond (Eleuthera, Bahamas). Sedimentology 2011, 58, 720–736. [Google Scholar] [CrossRef]
- Gallagher, K.L.; Kading, T.J.; Braissant, O.; Dupraz, C.; Visscher, P.T. Inside the alkalinity engine: The role of electron donors in the organomineralization potential of sulfate-reducing bacteria. Geobiology 2012, 10, 518–530. [Google Scholar] [CrossRef]
- Sanz-Montero, M.E.; Cabestrero, Ó.; Sánchez-Román, M. Microbial Mg-rich carbonates in an extreme alkaline lake (Las Eras, Central Spain). Front. Microbiol. 2019, 10, 148. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.R.; Gong, J.; Pajusalu, M.; Skoog, E.J.; Xu, M.; Feliz Soto, T.; Sojo, V.; Matreux, T.; Baldes, M.J.; Braun, D.; et al. A new model for silicification of cyanobacteria in Proterozoic tidal flats. Geobiology 2021, 19, 438–449. [Google Scholar] [CrossRef]
- Henke, J.M.; Bassler, B.L. Bacterial social engagements. Trends Cell Biol. 2004, 14, 648–656. [Google Scholar] [CrossRef]
- Bassler, B.L.; Losick, R. Bacterially speaking. Cell 2006, 125, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Balows, A.; Trüper, H.G.; Dworkin, M.; Harder, W.; Schleifer, K.H. (Eds.) The Prokaryotes: A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications; Springer Science + Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Risacher, F.; Alonso, H.; Salazar, C. The origin of brines and salts in Chilean salars: A hydrochemical review. Earth-Sci. Rev. 2003, 63, 249–293. [Google Scholar] [CrossRef]
- Hinman, N.W.; Cabrol, N.A.; Gulick, V.; Warren-Rhodes, K.; Tebes, C.; Chong, G.; Demergasso, C. Initial Investigations of Endoevaporitic Gypsum Habitats of Salar de Pajonales, Chile. In Astrobiology Science Conference, Mesa, AZ (Lunar and Planetary Institute, 2017), Arizona. Abstract; 2017; Volume 3568, Available online: https://www.hou.usra.edu/meetings/abscicon2017/eposter/3568.pdf (accessed on 14 May 2022).
- Chong Díaz, G.; Demergasso, C.; Urrutia Meza, J.; Vargas, M. El Dominio Salino del Norte de Chile y sus Yacimientos de Minerales Industriales. Bol. Soc. Geol. Mex. 2020, 72, 00018. [Google Scholar] [CrossRef]
- Pueyo, J.; Demergasso, C.; Escudero, L.; Chong, G.; Cortéz-Rivera, P.; Sanjurjo-Sánchez, J.; Carmona, V.; Giralt, S. On the origin of saline compounds in acidic salt flats (Central Andean Altiplano). Chem. Geol. 2021, 574, 120155. [Google Scholar] [CrossRef]
- Naranjo, J.A.; Villa, V.; Venegas, C. Geología de las áreas Salar de Pajonales y Cerro Moño, regiones de Antofagasta y Atacama. Serv. Nac. Geol. Min. Carta Geol. Chile Ser. Geol. Básica 2013, 153–154, 1. [Google Scholar]
- Rodríguez, C. Geología del Salar Pajonales (7209000-7226500N 510000-530 000E) y Antecedentes de su Microbiota, Antofagasta, Norte de Chile; Memoria Para Optar al Título de Geólogo; Universidad Católica del Norte: Antofagasta, Chile, 2018; 104p. [Google Scholar]
- Benison, K.C. The physical and chemical sedimentology of two high-altitude acid salars in Chile: Sedimentary processes in an extreme environment. J. Sediment. Res. 2019, 89, 147–167. [Google Scholar] [CrossRef] [Green Version]
- Dunai, T.J.; López, G.A.G.; Juez-Larré, J. Oligocene–Miocene age of aridity in the Atacama Desert revealed by exposure dating of erosion-sensitive landforms. Geology 2005, 33, 321–324. [Google Scholar] [CrossRef]
- Kampf, S.K.; Tyler, S.W. Spatial characterization of land surface energy fluxes and uncertainty estimation at the Salar de Atacama, Northern Chile. Adv. Water Resour. 2006, 29, 336–354. [Google Scholar] [CrossRef]
- Schween, J.H.; Hoffmeister, D.; Löhnert, U. Filling the observational gap in the Atacama Desert with a new network of climate stations. Glob. Planet. Change 2020, 184, 103034. [Google Scholar] [CrossRef]
- Duffie, J.A.; Beckman, W.A. Solar Radiation. In Solar Engineering of Thermal Processes, 4th ed.; Wiley: Hoboken, NJ, USA, 2013; pp. 12–20. [Google Scholar]
- Bozkurt, D.; Rondanelli, R.; Garreaud, R.; Arriagada, A. Impact of Warmer Eastern Tropical Pacific SST on the March 2015 Atacama Floods. Mon. Weather Rev. 2016, 144, 4441–4460. [Google Scholar] [CrossRef] [Green Version]
- Cabré, A.; Remy, D.; Aguilar, G.; Carretier, S.; Riquelme, R. Mapping rainstorm erosion associated with an individual storm from InSAR coherence loss validated by field evidence for the Atacama Desert. Earth Surf. Processes Landf. 2020, 45, 2091–2106. [Google Scholar] [CrossRef]
- Hinman, N.W.; Hofmann, M.H.; Warren-Rhodes, K.; Phillips, M.; Nofke, N.; Cabrol, N.A.; Chong-Diaz, G.; Demergasso, C.; Tebes Cayo, C.; Cabestrero, Ó.; et al. Surface Morphologies in a Mars-analog Ca-sulfate Salar, High Andes, Northern Chile. Front. Astron. Space Sci. 2022, 8, 235. [Google Scholar] [CrossRef]
- Phillips, M.S.; Moersch, J.E.; Cabrol, N.A.; Candela, A.; Wettergreen, D.; Warren-Rhodes, K.; Hinman, N.W.; Rhodes, R.; Team, T.S.I.N. Mapping planetary features from ground to orbit with Deep Learning: Application to habitats in a Mars-analog environment. Astrobiology 2022. Submitted. [Google Scholar]
- Escudero, L.W.; Bijman, J.; Chong, G.; Pueyo, J.J.; Demergasso, C. Geochemistry and microbiology in an acidic high altitude (4000 m) salt flat, high Andes, Northern Chile. Adv. Mater. Res. 2013, 825, 28–32. [Google Scholar] [CrossRef]
- Escudero, L.W.; Oetiker, N.; Gallardo, K.; Tebes-Cayo, C.; Guajardo, M.; Núñez, C.; Davis-Belmar, C.; Pueyo, J.J.; Chong, G.; Demergasso, C. A thiotrophic microbial comunity in an acidic brine lake in Northern Chile. Antonie Leeuwenhoek 2018, 111, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Tebes-Cayo, C.; Rodriguez, C.; Demergasso, C.; Chong, G.; Parro, V.; Sánchez-García, L.; Carrizo, D.; Hinman, N.W.; Warren-Rhodes, K.; Cabrol, N.A. Microbial Participation on the Formation and the Preservation of Gypsum Structures from Salar de Pajonales, Northern of Chile. In 2019 Astrobiology Science Conference; AGU: Washington, DC, USA, 2019; Available online: https://agu.confex.com/data/extendedabstract/agu/abscicon19/Paper_482718_extendedabstract_31733_0.pdf (accessed on 14 May 2022).
- Tebes-Cayo, C.; Demergasso, C.; Chong, G.; Cabestrero, Ó.; Sanz-Montero, M.E.; Castro-Nallar, E.; Cabrol, N. Geochemistry and microbial communities in Salar de Pajonales and Salar de Gorbea, Northern Chile: Influence on the gypsum microbialites formation. Geotemas 2021, 18, 458–462. [Google Scholar]
- Otálora, F.; Criado-Reyes, J.; Baselga, M.; Canals, A.; Verdugo-Escamilla, C.; García Ruiz, J.M. Hydrochemical and mineralogical evolution through evaporitic processes in Salar de Llamara Brines (Atacama, Chile). ACS Earth Space Chem. 2020, 4, 882–896. [Google Scholar] [CrossRef]
- Reid, R.P.; Oehlert, A.M.; Suosaari, E.P.; Demergasso, C.; Chong, G.; Escudero, L.V.; Piggot, A.M.; Lascu, I.; Palma, A.T. Electrical conductivity as a driver of biological and geological spatial heterogeneity in the Puquios, Salar de Llamara, Atacama Desert, Chile. Sci. Rep. 2021, 11, 12769. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, J.; Jodar, J.; Medina, A.; Herrera, C.; Chong, G.; Urqueta, H.; Luque, J.A. Hydrogeology and sustainable future groundwater abstraction from the Agua Verde aquifer in the Atacama Desert, northern Chile. Hydrogeol. J. 2018, 26, 1989–2007. [Google Scholar] [CrossRef]
- Benison, K.C.; Karmanocky, F.J., III. Could microorganisms be preserved in Mars gypsum? Insights from terrestrial examples. Geology 2014, 42, 615–618. [Google Scholar] [CrossRef] [Green Version]
- Parkhurst, D.L.; Appelo, C.A.J. Description of Input and Examples for PHREEQC Version 3: A Computer Program for Speciation, Batch-Reaction, One-Dimensional Transport, and Inverse Geochemical Calculations; U.S. Geological Survey Techniques and Methods, No. 6-A43. 2013. Available online: http://pubs.usgs.gov/tm/06/a43 (accessed on 14 May 2022).
- Pitzer, K.S. Thermodynamics of electrolytes. I. Theoretical basis and general equations. J. Phys. Chem. 1973, 77, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Castenholz, R.W. Phylum BX Cyanobacteria. Oxigenic photosynthetic bacteria. In Bergey’s Manual of Systematic Bacteriology; Boone, D.R., Castenholz, R.W., Garrity, G.M., Eds.; The Archea and Depply Branching and Photrophic Bacteria; Springer: New York, NY, USA, 2001; Volume 1, pp. 473–599. [Google Scholar]
- Díaz, C.; Maidana, N. Diatomeas de los Salares de Atacama y Punta Negra II Región; Centro de Ecología Aplicada: La Reina, Chile, 2005. [Google Scholar]
- Silverstone, S.; Nelson, M.; Alling, A.; Allen, J.P. Soil and crop management experiments in the Laboratory Biosphere: An analogue system for the Mars on Earth® facility. Adv. Space Res. 2005, 35, 1544–1551. [Google Scholar] [CrossRef]
- Cockell, C.S.; Santomartino, R.; Finster, K.; Waajen, A.C.; Eades, L.J.; Moeller, R.; Rettberg, P.; Fuchs, F.M.; Van Houdt, R.; Leys, N.; et al. Space station biomining experiment demonstrates rare earth element extraction in microgravity and Mars gravity. Nat. Commun. 2020, 11, 5523. [Google Scholar] [CrossRef]
- Graue, J.; Engelen, B.; Cypionka, H. Degradation of cyanobacterial biomass in anoxic tidal-flat sediments: A microcosm study of metabolic processes and community changes. ISME J. 2012, 6, 660–669. [Google Scholar] [CrossRef] [Green Version]
- García-Maldonado, J.Q.; Escobar-Zepeda, A.; Raggi, L.; Bebout, B.M.; Sanchez-Flores, A.; López-Cortés, A. Bacterial and archaeal profiling of hypersaline microbial mats and endoevaporites, under natural conditions and methanogenic microcosm experiments. Extremophiles 2018, 22, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Llirós, M.; Gaju, N.; de Oteyza, T.G.; Grimalt, J.O.; Esteve, I.; Martínez-Alonso, M. Microcosm experiments of oil degradation by microbial mats. II. The changes in microbial species. Sci. Total Environ. 2008, 393, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Bebout, B.M.; Carpenter, S.P.; Des Marais, D.J.; Discipulo, M.; Embaye, T.; Garcia-Pichel, F.; Hoehler, T.M.; Hogan, M.; Jahnke, L.L.; Keller, R.M.; et al. Long-term manipulations of intact microbial mat communities in a greenhouse collaboratory: Simulating earth’s present and past field environments. Astrobiology 2002, 2, 383–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, T.; Decho, A.W. Isolation and biochemical characterization of extracellular polymeric secretions (EPS) from modern soft marine stromatolites (Bahamas) and its inhibitory effect on CaCO3 precipitation. Prep. Biochem. Biotechnol. 2002, 32, 51–63. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, N.; Wilson, L.A.; Brasier, M.D. Growth of synthetic stromatolites and wrinkle structures in the absence of microbes–implications for the early fossil record. Geobiology 2008, 6, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Couradeau, E.; Benzerara, K.; Moreira, D.; Gerard, E.; Kaźmierczak, J.; Tavera, R.; López-García, P. Prokaryotic and eukaryotic community structure in field and cultured microbialites from the alkaline Lake Alchichica (Mexico). PLoS ONE 2011, 6, e28767. [Google Scholar] [CrossRef]
- Newman, S.A.; Mariotti, G.; Pruss, S.; Bosak, T. Insights into cyanobacterial fossilization in Ediacaran siliciclastic environments. Geology 2016, 44, 579–582. [Google Scholar] [CrossRef]
- Newman, S.A.; Klepac-Ceraj, V.; Mariotti, G.; Pruss, S.B.; Watson, N.; Bosak, T. Experimental fossilization of mat-forming cyanobacteria in coarse-grained siliciclastic sediments. Geobiology 2017, 15, 484–498. [Google Scholar] [CrossRef]
- Osman, J.R.; Viedma, P.; Mendoza, J.; Cotoras, D. Bacterial and Geochemical Composition of Thrombolites from Lake Sarmiento, Torres del Paine National Park of Chilean Patagonia. Geomicrobiol. J. 2020, 37, 376–388. [Google Scholar] [CrossRef]
- Osman, J.R.; Viedma, P.; Mendoza, J.; Fernandes, G.; DuBow, M.S.; Cotoras, D. Prokaryotic diversity and biogeochemical characteristics of field living and laboratory cultured stromatolites from the hypersaline Laguna Interna, Salar de Atacama (Chile). Extremophiles 2021, 25, 327–342. [Google Scholar] [CrossRef]
- Baker, M. 1500 scientists lift the lid on reproducibility. Nat. News 2016, 533, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabestrero, Ó.; Sanz-Montero, M.E.; del Buey, P.; Tebes-Cayo, C.; Demergasso, C.; Visscher, P.T. Experiments with microbial mats from shallow lakes: Metabolic influence of the microorganisms in the precipitation of minerals. Geotemas 2021, 18, 144–146. [Google Scholar]
- Marín, S.; Cabestrero, O.; Demergasso, C.; Olivares, S.; Zetola, V.; Vera, M. An indigenous bacterium with enhanced performance of microbially-induced Ca-carbonate biomineralization under extreme alkaline conditions for concrete and soil-improvement industries. Acta Biomater. 2021, 120, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Popall, R.M.; Bolhuis, H.; Muyzer, G.; Sánchez-Román, M. Stromatolites as biosignatures of atmospheric oxygenation: Carbonate biomineralization and UV-C resilience in a Geitlerinema sp.-dominated culture. Front. Microbiol. 2020, 11, 948. [Google Scholar] [CrossRef]
- Mellage, A.; Smeaton, C.M.; Furman, A.; Atekwana, E.A.; Rezanezhad, F.; Van Cappellen, P. Linking spectral induced polarization (SIP) and subsurface microbial processes: Results from sand column incubation experiments. Environ. Sci. Technol. 2018, 52, 2081–2090. [Google Scholar] [CrossRef]
- Duteil, T.; Bourillot, R.; Grégoire, B.; Virolle, M.; Brigaud, B.; Nouet, J.; Braissant, O.; Portier, E.; Féniès, H.; Patrier, P.; et al. Experimental formation of clay-coated sand grains using diatom biofilm exopolymers. Geology 2020, 48, 1012–1017. [Google Scholar] [CrossRef]
- Moore, K.R.; Pajusalu, M.; Gong, J.; Sojo, V.; Matreux, T.; Braun, D.; Bosak, T. Biologically mediated silicification of marine cyanobacteria and implications for the Proterozoic fossil record. Geology 2020, 48, 862–866. [Google Scholar] [CrossRef]
- Brehm, U.; Gorbushina, A.; Mottershead, D. The role of microorganisms and biofilms in the breakdown and dissolution of quartz and glass. In Geobiology: Objectives, Concepts, Perspectives; Elsevier: Amsterdam, The Netherlands, 2005; pp. 117–129. [Google Scholar]
- Palm, A. The Dissolution of Diatoms in Marine Microbial Mats. Master’s Thesis, University of Connecticut, Storrs, CT, USA, 2018; 92p. Available online: https://opencommons.uconn.edu/gs_theses/1219 (accessed on 14 May 2022).
- Fecskeová, L.K.; Piwosz, K.; Hanusová, M.; Nedoma, J.; Znachor, P.; Koblížek, M. Diel changes and diversity of pufM expression in freshwater communities of anoxygenic phototrophic bacteria. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Guerrero, M.C.; Tadeo, A.B.; de Wit, R. Environmental factors controlling the development of microbial mats in inland saline lakes; the granulometric composition of the sediment. In Microbial Mats; Springer: Berlin/Heidelberg, Germany, 1994; pp. 85–90. [Google Scholar]
- Prisciandaro, M.; Lancia, A.; Musmarra, D. The retarding effect of citric acid on calcium sulfate nucleation kinetics. Ind. Eng. Chem. Res. 2003, 42, 6647–6652. [Google Scholar] [CrossRef]
- Rabizadeh, T.; Peacock, C.L.; Benning, L.G. Carboxylic acids: Effective inhibitors for calcium sulfate precipitation? Mineral. Mag. 2014, 78, 1465–1472. [Google Scholar] [CrossRef] [Green Version]
- Ossorio, M.; Stawski, T.M.; Rodríguez-Blanco, J.D.; Sleutel, M.; García-Ruiz, J.M.; Benning, L.G.; Van Driessche, A.E. Physicochemical and additive controls on the multistep precipitation pathway of gypsum. Minerals 2017, 7, 140. [Google Scholar] [CrossRef] [Green Version]
- Nicoleau, L.; Van Driessche, A.E.; Kellermeier, M. A kinetic analysis of the role of polymers in mineral nucleation. The example of gypsum. Cem. Concr. Res. 2019, 124, 105837. [Google Scholar] [CrossRef]
- Sangwal, K. Additives and Crystallization Processes: From Fundamentals to Applications; John Wiley & Sons: Hoboken, NJ, USA, 2007. [Google Scholar]
- Amjad, Z.; Demadis, K.D. (Eds.) Mineral Scales and Deposits: Scientific and Technological Approaches; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Dupraz, C.; Reid, R.P.; Braissant, O.; Decho, A.W.; Norman, R.S.; Visscher, P.T. Processes of carbonate precipitation in modern microbial mats. Earth-Sci. Rev. 2009, 96, 141–162. [Google Scholar] [CrossRef]
- Douglas, T.; Mann, S. Oriented nucleation of gypsum (CaSO4·2H2O) under compressed Langmuir monolayers. Mater. Sci. Eng. C 1994, 1, 193–199. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Miao, R.; Lv, Y.; Wang, X.; Meng, X.; Yang, R.; Zhang, X. Enhanced gypsum scaling by organic fouling layer on nanofiltration membrane: Characteristics and mechanisms. Water Res. 2016, 91, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Wierzchos, J.; Casero, M.C.; Artieda, O.; Ascaso, C. Endolithic microbial habitats as refuges for life in polyextreme environment of the Atacama Desert. Curr. Opin. Microbiol. 2018, 43, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, D.B. Precipitation Pathways of Amorphous Silica. Ph.D. Thesis, University of Leeds, Leeds, UK, 2016; 241p. [Google Scholar]
- Daza Brunet, R. Distribución, Composición y Génesis (Biótica/Abiótica) de los Espeleotemas de las Cuevas Volcánicas de la Isla Terceira (Azores, Portugal). Ph.D. Thesis, Complutense University of Madrid, Madrid, Spain, 2015; 178p. [Google Scholar]
- van den Heuvel, D.B.; Stawski, T.M.; Tobler, D.J.; Wirth, R.; Peacock, C.L.; Benning, L.G. Formation of silica-lysozyme composites through co-Precipitation and adsorption. Front. Mater. 2018, 5, 19. [Google Scholar] [CrossRef]
- Konhauser, K.O. Introduction to Geomicrobiology; Blackwell Science Ltd.: Oxford, UK, 2007; 452p. [Google Scholar]
- Paerl, H.W.; Steppe, T.F.; Reid, R.P. Bacterially mediated precipitation in marine stromatolites. Environ. Microbiol. 2001, 3, 123–130. [Google Scholar] [CrossRef]
- Pozo, M.; Calvo, J.P. An overview of authigenic magnesian clays. Minerals 2018, 8, 520. [Google Scholar] [CrossRef] [Green Version]
- Martín-Pérez, A.; La Iglesia, Á.; Almendros, G.; González-Pérez, J.A.; Alonso-Zarza, A.M. Precipitation of kerolite and sepiolite associated with Mg-rich carbonates in a cave environment. Sediment. Geol. 2021, 411, 105793. [Google Scholar] [CrossRef]
- Cortez, P. Caracterización Geológica de los Salares Gorbea e Ignorado y su Asociación con su Microbiota; Universidad Católica del Norte: Antofagasta, Chile, 2014; 200p. [Google Scholar]
- Rouchy, J.M.; Monty, C. Gypsum microbial sediments: Neogene and modern examples. In Microbial Sediments; Springer: Berlin/Heidelberg, Germany, 2000; pp. 209–216. [Google Scholar]
- Sanz-Montero, M.E.; Rodriguez-Aranda, J.P.; Calvo, J.P. Mediation of endoevaporitic microbial communities in early replacement of gypsum by dolomite: A case study from Miocene Lake deposits of the Madrid Basin, Spain. J. Sediment. Res. 2006, 76, 1257–1266. [Google Scholar] [CrossRef]
- Sanz-Montero, M.E.; Rodríguez-Aranda, J.P.; del Cura, M.A.G. Bioinduced precipitation of barite and celestite in dolomite microbialites: Examples from Miocene lacustrine sequences in the Madrid and Duero Basins, Spain. Sediment. Geol. 2009, 222, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Allwood, A.C.; Burch, I.W.; Rouchy, J.M.; Coleman, M. Morphological biosignatures in gypsum: Diverse formation processes of Messinian (∼6.0 Ma) gypsum stromatolites. Astrobiology 2013, 13, 870–886. [Google Scholar] [CrossRef] [PubMed]
- Manning-Berg, A.R.; Kah, L.C. Proterozoic microbial mats and their constraints on environments of silicification. Geobiology 2017, 15, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Manning-Berg, A.R.; Wood, R.S.; Williford, K.H.; Czaja, A.D.; Kah, L.C. The taphonomy of proterozoic microbial mats and implications for early diagenetic silicification. Geosciences 2019, 9, 40. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salar | Pajonales | Gorbea | |||||
|---|---|---|---|---|---|---|---|
| Sample | PAJ-10 | PAJ-1 | PAJ-7 | PAJ-13 | GOR-2 | GOR-1 | |
| Salinity | % w/v | 1.9 | 4.0 | 17.3 | 24.2 | 1.2 | 10.6 |
| Temperature | °C | 11.1 | 13.6 | 14.3 | 19.8 | 11 | 9 |
| pH | 7.7 | 8.0 | 7.3 | 6.6 | 4.6 | 3.3 | |
| Na+ | mg/L | 3680 ± 368 | 107,85 ± 1078 | 51,760 ± 5176 | 100,220 ± 10,022 | 2250 ± 225 | 9511 ± 951 |
| Mg2+ | mg/L | 223 ± 22 | 561 ± 56 | 3090 ± 309 | 5840 ± 584 | 16 ± 2 | 8760 ± 876 |
| Ca2+ | mg/L | 678 ± 68 | 2326 ± 233 | 6895 ± 690 | 9985 ± 999 | 370 ± 37 | 600 ± 60 |
| K+ | mg/L | 210 ± 21 | 340 ± 34 | 2150 ± 215 | 2670 ± 267 | 250 +25 | 1224 ± 122 |
| SiO2 | mg/L | 67 ± 7 | 69.5 ± 7 | 72.5 ± 7 | 78.9 ± 7 | 134.1 ± 13 | 103.1 ± 10 |
| Cl− | mg/L | 7397 ± 740 | 20,460 ± 2046 | 104,000 ± 10,400 | 198,700 ± 19,870 | 3700 ± 370 | 37,970 ± 3797 |
| SO42− | mg/L | 470 ± 47 | 1320 ± 132 | 2240 ± 224 | 760 ± 76 | 1090 ± 109 | 12,000 ± 1200 |
| HCO3− | mg/L | 63.7 ± 6 | 95 ± 10 | 49 ± 5 | 92 ± 10 | 0 | 0 |
| Calcite SI | log Q/K | 0.3 | 1.2 | 1.1 | 1.9 | - | - |
| Gypsum SI | log Q/K | –0.8 | –0.2 | 0.2 | 0.4 | –0.5 | –0.2 |
| Halite SI | log Q/K | –3.3 | –2.5 | –0.8 | 0.4 | –3.8 | –2.2 |
| Opal SI | log Q/K | –0.3 | –0.2 | 0.1 | 0.8 | 0.0 | 0.1 |
| Sepiolite SI | log Q/K | 1.1 | 3.1 | 3.8 | 6.2 | –12.6 | –12.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabestrero, Ó.; Tebes-Cayo, C.; Hinman, N.W.; Demergasso, C. Mineral Paragenesis Precipitating in Salt Flat Pools of Continental Environments Replicated in Microbial Mat Microcosms without Evaporation. Minerals 2022, 12, 646. https://0-doi-org.brum.beds.ac.uk/10.3390/min12050646
Cabestrero Ó, Tebes-Cayo C, Hinman NW, Demergasso C. Mineral Paragenesis Precipitating in Salt Flat Pools of Continental Environments Replicated in Microbial Mat Microcosms without Evaporation. Minerals. 2022; 12(5):646. https://0-doi-org.brum.beds.ac.uk/10.3390/min12050646
Chicago/Turabian StyleCabestrero, Óscar, Cinthya Tebes-Cayo, Nancy W. Hinman, and Cecilia Demergasso. 2022. "Mineral Paragenesis Precipitating in Salt Flat Pools of Continental Environments Replicated in Microbial Mat Microcosms without Evaporation" Minerals 12, no. 5: 646. https://0-doi-org.brum.beds.ac.uk/10.3390/min12050646