miR-196B-5P and miR-200B-3P Are Differentially Expressed in Medulloblastomas of Adults and Children

 , , , , , , and
, , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Patient Samples
2.3. Nucleic Acid Extraction
2.4. Immunostaining
2.5. CTNNB1 and TP53 Mutational Screening
2.6. Mirnome Expression Analysis
2.7. MicroRNAs Validation
2.8. miRNAs Target Prediction and Pathway Enrichment
2.9. Target Differentially Expressed in Adult vs. Childhood MBs
2.10. Statistical Analysis
2.11. Construction of Protein–Protein Interaction (PPI) Network
2.12. Data Availability
3. Results
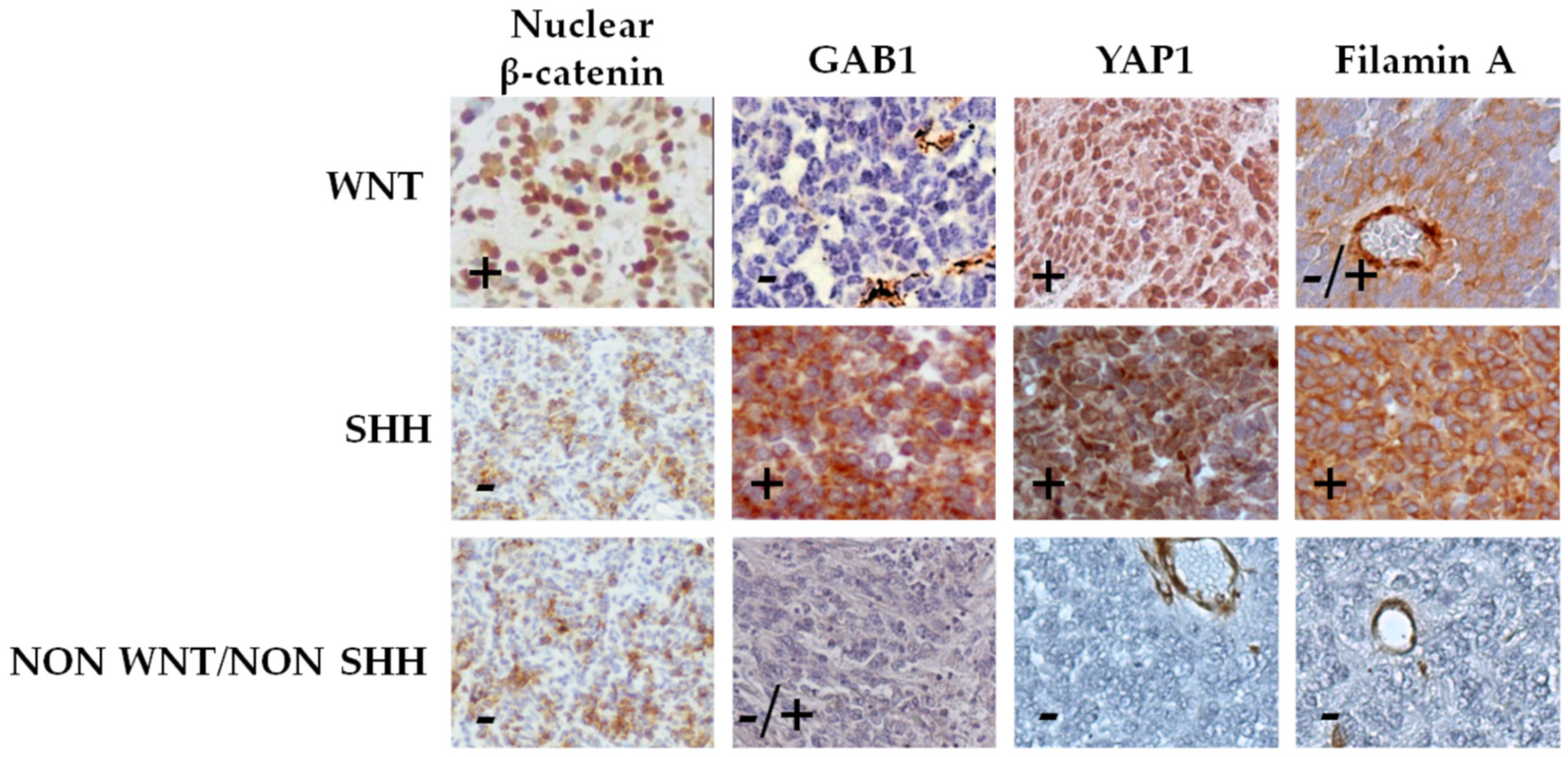
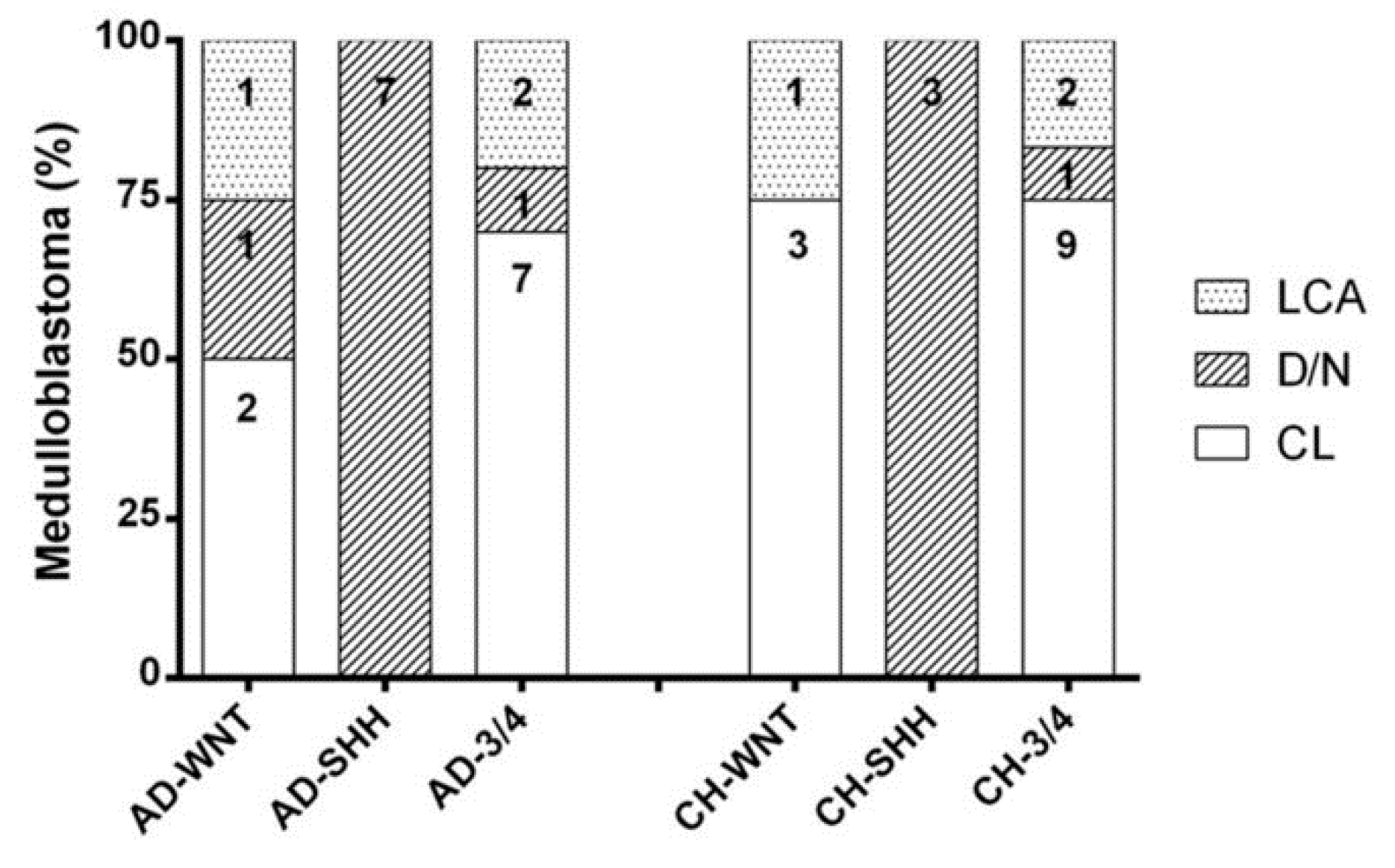
3.1. Patients Subgrouping
3.2. miRNome Expression Analysis in the Adult and Childhood Medulloblastoma
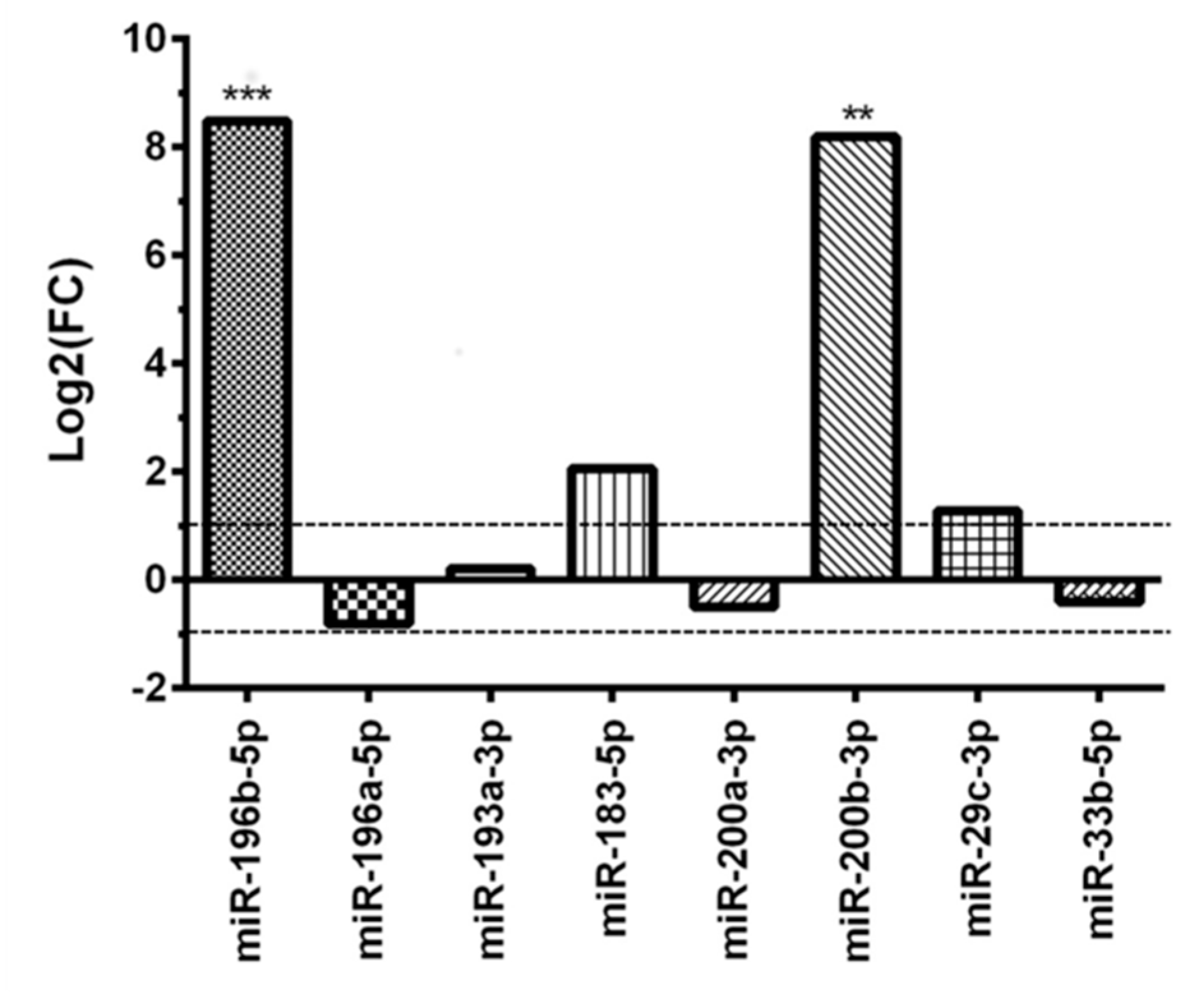
3.3. Validation of the Subset of Eight miRNAS in the Adult and Childhood Medulloblastoma Groups
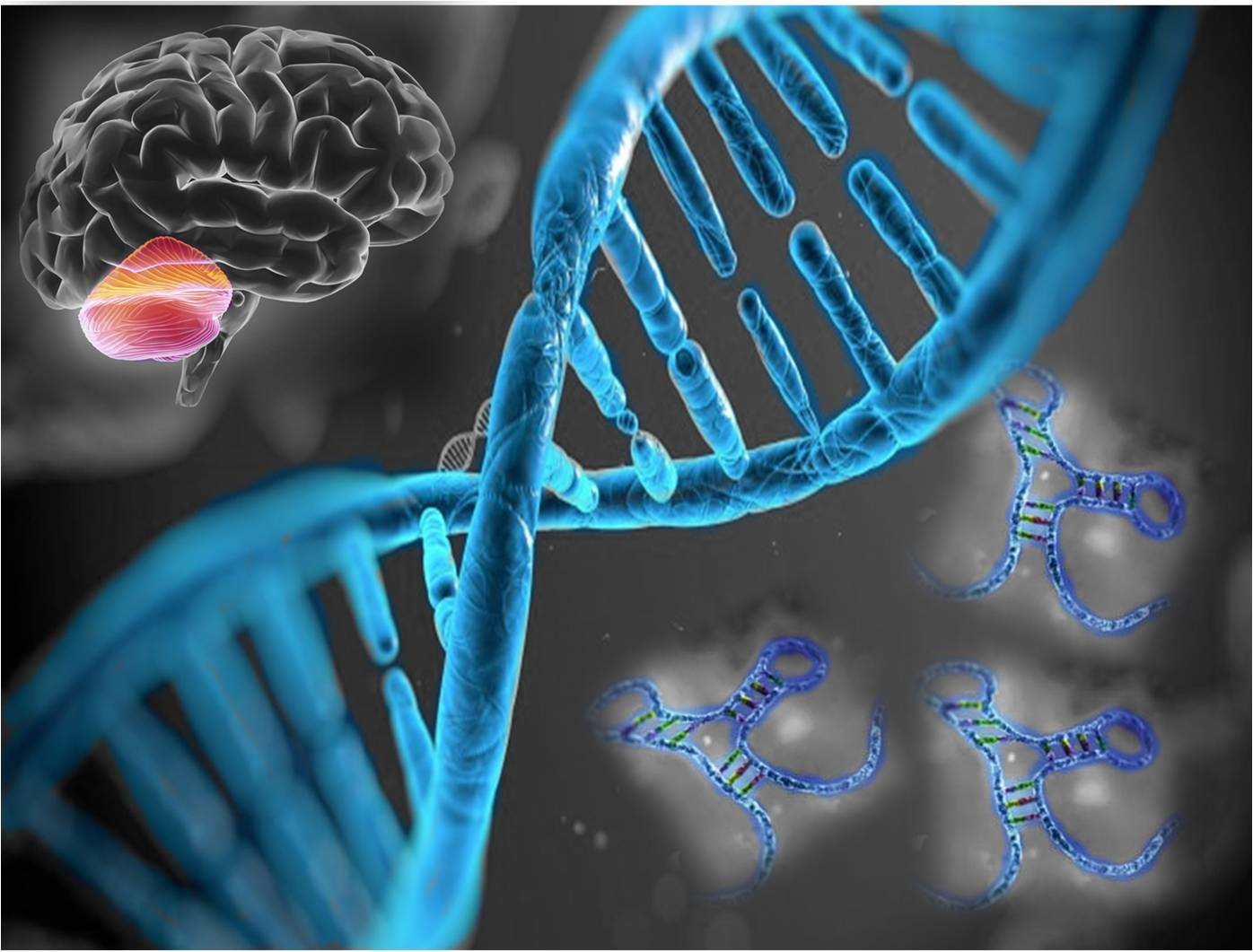
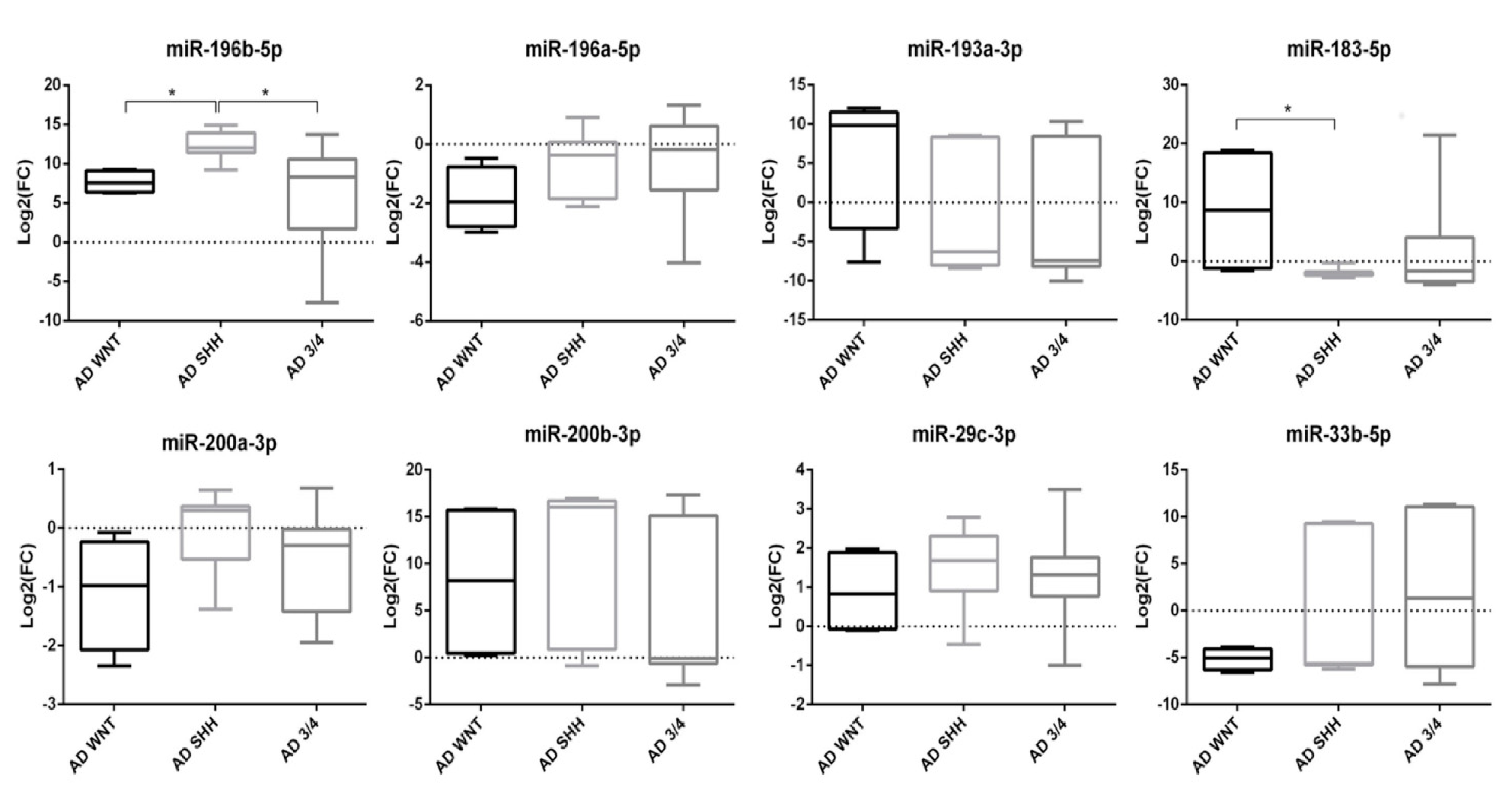
3.4. miRNAs Expression in Molecular Subgroups of Adult Medulloblastoma
3.5. Analysis of the Predicted Target Genes and Pathway Enrichment Analysis
3.6. Genes Differentially Expressed in Adult vs. Childhood MBs According to GEO Datasets
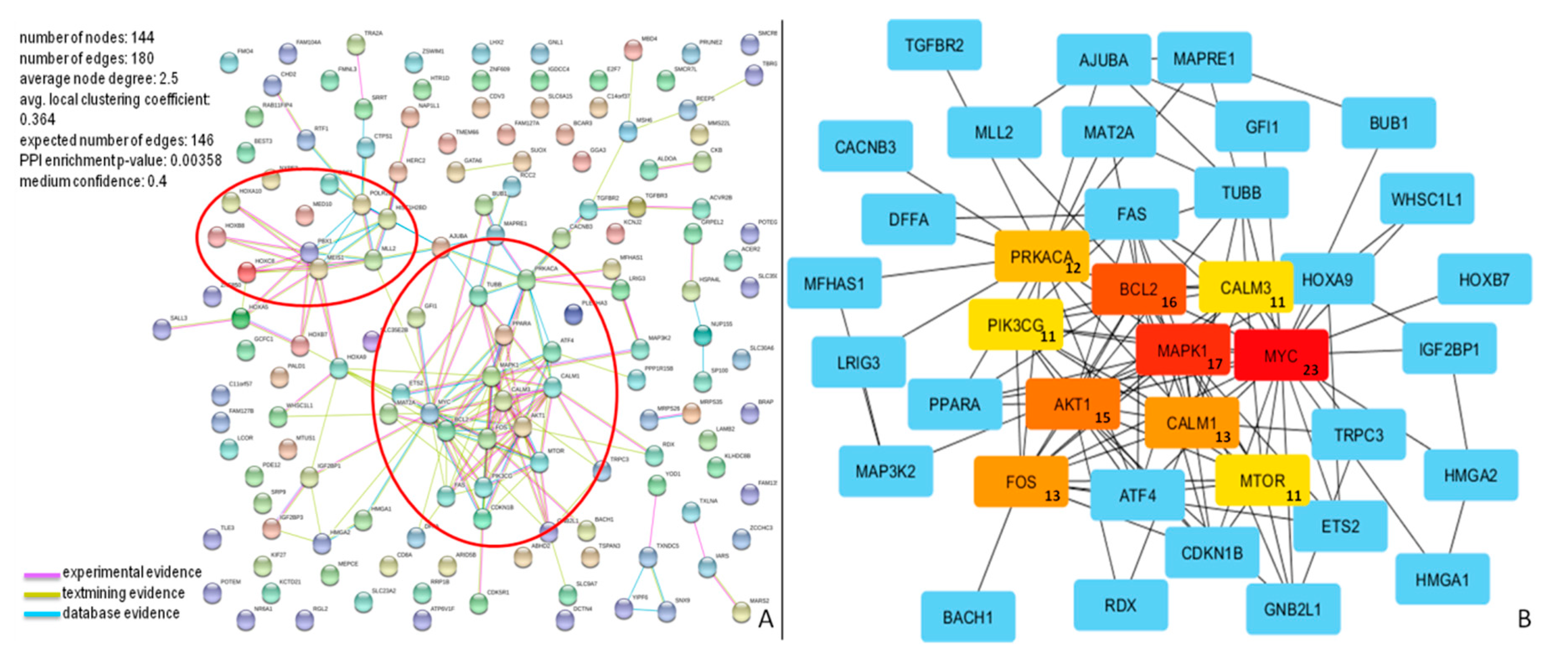
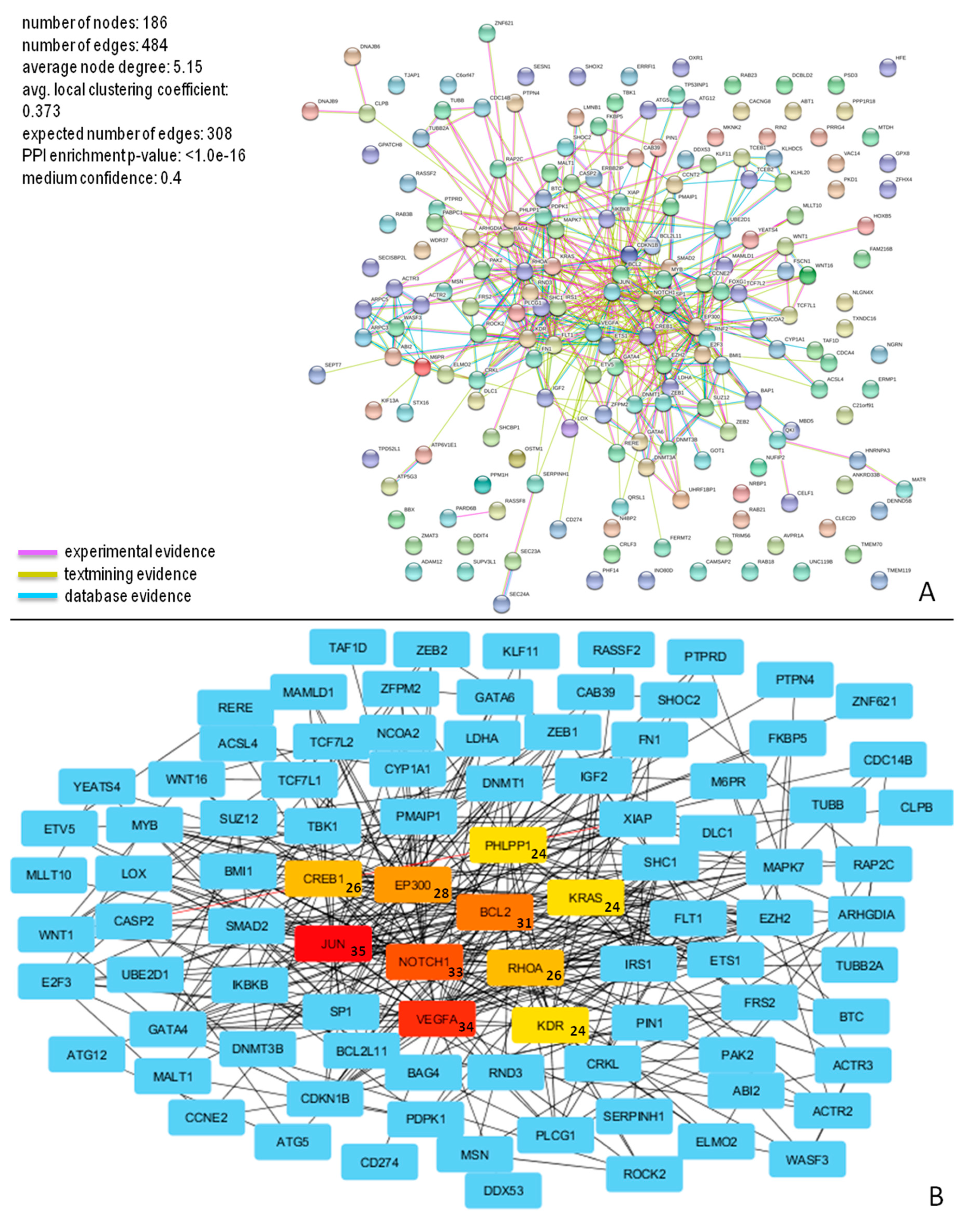
3.7. PPI Network of miR-196b-5p and miR-200b-3p Targets
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rickert, C.H.; Paulus, W. Epidemiology of central nervous system tumors in childhood and adolescence based on the new WHO classification. Childs Nerv. Syst. 2001, 17, 503–511. [Google Scholar] [CrossRef] [PubMed]
- McNeil, D.E.; Cote, T.R.; Clegg, L.; Rorke, L.B. Incidence and trends in pediatric malignancies medulloblastoma/primitive neuroectodermal tumor: A SEER update. Surveillance Epidemiology and End Results. Med. Pediatr. Oncol. 2002, 39, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; de Blank, P.M.; Kruchko, C.; Petersen, C.M.; Liao, P.; Finlay, J.L.; Stearns, D.S.; Wolff, J.E.; Wolinsky, Y.; Letterio, J.J.; et al. Alex’s Lemonade Stand Foundation Infant and Childhood Primary Brain and Central Nervous System Tumors Diagnosed in the United States in 2007–2011. Neuro Oncol. 2015, 16 (Suppl. 10), x1–x36. [Google Scholar] [CrossRef]
- Smoll, N.R.; Drummond, K.J. The incidence of medulloblastomas and primitive neurectodermal tumours in adults and children. J. Clin. Neurosci. 2012, 19, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Bloom, H.J.; Bessell, E.M. Medulloblastoma in adults: A review of 47 patients treated between 1952 and 1981. Int. J. Radiat. Oncol. Biol. Phys. 1990, 18, 763–772. [Google Scholar] [CrossRef]
- Wrensch, M.; Minn, Y.; Chew, T.; Bondy, M.; Berger, M.S. Epidemiology of primary brain tumors: Current concepts and review of the literature. Neuro Oncol. 2002, 4, 278–299. [Google Scholar] [CrossRef]
- Giordana, M.T.; Cavalla, P.; Dutto, A.; Borsotti, L.; Chio, A.; Schiffer, D. Is medulloblastoma the same tumor in children and adults? J. Neurooncol. 1997, 35, 169–176. [Google Scholar] [CrossRef]
- Korshunov, A.; Remke, M.; Werft, W.; Benner, A.; Ryzhova, M.; Witt, H.; Sturm, D.; Wittmann, A.; Schottler, A.; Felsberg, J.; et al. Adult and pediatric medulloblastomas are genetically distinct and require different algorithms for molecular risk stratification. J. Clin. Oncol. 2010, 28, 3054–3060. [Google Scholar] [CrossRef]
- Rodriguez, F.J.; Eberhart, C.; O’Neill, B.P.; Slezak, J.; Burger, P.C.; Goldthwaite, P.; Wu, W.; Giannini, C. Histopathologic grading of adult medulloblastomas. Cancer 2007, 109, 2557–2565. [Google Scholar] [CrossRef]
- Ang, C.; Hauerstock, D.; Guiot, M.C.; Kasymjanova, G.; Roberge, D.; Kavan, P.; Muanza, T. Characteristics and outcomes of medulloblastoma in adults. Pediatr. Blood Cancer 2008, 51, 603–607. [Google Scholar] [CrossRef]
- Parsons, D.W.; Li, M.; Zhang, X.; Jones, S.; Leary, R.J.; Lin, J.C.; Boca, S.M.; Carter, H.; Samayoa, J.; Bettegowda, C.; et al. The genetic landscape of the childhood cancer medulloblastoma. Science 2011, 331, 435–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, T.C.; Mahoney, E.L.; Pomeroy, S.L. Medulloblastoma: Molecular Classification-Based Personal Therapeutics. Neurotherapeutics 2017, 14, 265–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Halabi, H.; Nantel, A.; Klekner, A.; Guiot, M.C.; Albrecht, S.; Hauser, P.; Garami, M.; Bognar, L.; Kavan, P.; Gerges, N.; et al. Preponderance of sonic hedgehog pathway activation characterizes adult medulloblastoma. Acta Neuropathol. 2011, 121, 229–239. [Google Scholar] [CrossRef]
- Remke, M.; Hielscher, T.; Northcott, P.A.; Witt, H.; Ryzhova, M.; Wittmann, A.; Benner, A.; von Deimling, A.; Scheurlen, W.; Perry, A.; et al. Adult medulloblastoma comprises three major molecular variants. J. Clin. Oncol. 2011, 29, 2717–2723. [Google Scholar] [CrossRef]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, E.; De Smaele, E.; Po, A.; Di Marcotullio, L.; Tosi, E.; Espinola, M.S.; Di Rocco, C.; Riccardi, R.; Giangaspero, F.; Farcomeni, A.; et al. MicroRNA profiling in human medulloblastoma. Int. J. Cancer 2009, 124, 568–577. [Google Scholar] [CrossRef]
- Pang, J.C.; Kwok, W.K.; Chen, Z.; Ng, H.K. Oncogenic role of microRNAs in brain tumors. Acta Neuropathol. 2009, 117, 599–611. [Google Scholar] [CrossRef]
- Pierson, J.; Hostager, B.; Fan, R.; Vibhakar, R. Regulation of cyclin dependent kinase 6 by microRNA 124 in medulloblastoma. J. Neurooncol. 2008, 90, 1–7. [Google Scholar] [CrossRef]
- Northcott, P.A.; Fernandez, L.A.; Hagan, J.P.; Ellison, D.W.; Grajkowska, W.; Gillespie, Y.; Grundy, R.; Van Meter, T.; Rutka, J.T.; Croce, C.M.; et al. The miR-17/92 polycistron is up-regulated in sonic hedgehog-driven medulloblastomas and induced by N-myc in sonic hedgehog-treated cerebellar neural precursors. Cancer Res. 2009, 69, 3249–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weeraratne, S.D.; Amani, V.; Teider, N.; Pierre-Francois, J.; Winter, D.; Kye, M.J.; Sengupta, S.; Archer, T.; Remke, M.; Bai, A.H.; et al. Pleiotropic effects of miR-183~96~182 converge to regulate cell survival, proliferation and migration in medulloblastoma. Acta Neuropathol. 2012, 123, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Uziel, T.; Karginov, F.V.; Xie, S.; Parker, J.S.; Wang, Y.D.; Gajjar, A.; He, L.; Ellison, D.; Gilbertson, R.J.; Hannon, G.; et al. The miR-17~92 cluster collaborates with the Sonic Hedgehog pathway in medulloblastoma. Proc. Natl. Acad. Sci. USA 2009, 106, 2812–2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, B.L.; Obad, S.; Bihannic, L.; Ayrault, O.; Zindy, F.; Kauppinen, S.; Roussel, M.F. Silencing of the miR-17~92 cluster family inhibits medulloblastoma progression. Cancer Res. 2013, 73, 7068–7078. [Google Scholar] [CrossRef] [Green Version]
- Bai, A.H.; Milde, T.; Remke, M.; Rolli, C.G.; Hielscher, T.; Cho, Y.J.; Kool, M.; Northcott, P.A.; Jugold, M.; Bazhin, A.V.; et al. MicroRNA-182 promotes leptomeningeal spread of non-sonic hedgehog-medulloblastoma. Acta Neuropathol. 2012, 123, 529–538. [Google Scholar] [CrossRef]
- Grunder, E.; D’Ambrosio, R.; Fiaschetti, G.; Abela, L.; Arcaro, A.; Zuzak, T.; Ohgaki, H.; Lv, S.Q.; Shalaby, T.; Grotzer, M. MicroRNA-21 suppression impedes medulloblastoma cell migration. Eur. J. Cancer 2011, 47, 2479–2490. [Google Scholar] [CrossRef]
- Liu, W.; Gong, Y.H.; Chao, T.F.; Peng, X.Z.; Yuan, J.G.; Ma, Z.Y.; Jia, G.; Zhao, J.Z. Identification of differentially expressed microRNAs by microarray: A possible role for microRNAs gene in medulloblastomas. Chin. Med. J. (Engl.) 2009, 122, 2405–2411. [Google Scholar]
- Garzia, L.; Andolfo, I.; Cusanelli, E.; Marino, N.; Petrosino, G.; De Martino, D.; Esposito, V.; Galeone, A.; Navas, L.; Esposito, S.; et al. MicroRNA-199b-5p impairs cancer stem cells through negative regulation of HES1 in medulloblastoma. PLoS ONE 2009, 4, e4998. [Google Scholar] [CrossRef]
- Li, K.K.; Pang, J.C.; Ching, A.K.; Wong, C.K.; Kong, X.; Wang, Y.; Zhou, L.; Chen, Z.; Ng, H.K. miR-124 is frequently down-regulated in medulloblastoma and is a negative regulator of SLC16A1. Hum. Pathol. 2009, 40, 1234–1243. [Google Scholar] [CrossRef]
- Venkataraman, S.; Alimova, I.; Fan, R.; Harris, P.; Foreman, N.; Vibhakar, R. MicroRNA 128a increases intracellular ROS level by targeting Bmi-1 and inhibits medulloblastoma cancer cell growth by promoting senescence. PLoS ONE 2010, 5, e10748. [Google Scholar] [CrossRef]
- Li, K.K.; Xia, T.; Ma, F.M.; Zhang, R.; Mao, Y.; Wang, Y.; Zhou, L.; Lau, K.M.; Ng, H.K. miR-106b is overexpressed in medulloblastomas and interacts directly with PTEN. Neuropathol. Appl. Neurobiol. 2015, 41, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.A.; Lu, D.L.; Huang, X.; Tan, W. miR-219 inhibits the proliferation, migration and invasion of medulloblastoma cells by targeting CD164. Int. J. Mol. Med. 2014, 34, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Wang, Z.; Wan, B.; Zheng, Z. MicroRNA-206 inhibits the viability and migration of medulloblastoma cells by targeting LIM and SH3 protein 1. Exp. Ther. Med. 2017, 14, 3894–3900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.Y.; Zhu, B.; Zhao, X.W.; Zhan, Y.B.; Bao, J.J.; Zhou, J.Q.; Zhang, F.J.; Yu, B.; Liu, J.; Wang, Y.M.; et al. Regulation of UHRF1 by microRNA-378 modulates medulloblastoma cell proliferation and apoptosis. Oncol. Rep. 2017, 38, 3078–3084. [Google Scholar] [CrossRef]
- Singh, S.V.; Dakhole, A.N.; Deogharkar, A.; Kazi, S.; Kshirsagar, R.; Goel, A.; Moiyadi, A.; Jalali, R.; Sridhar, E.; Gupta, T.; et al. Restoration of miR-30a expression inhibits growth, tumorigenicity of medulloblastoma cells accompanied by autophagy inhibition. Biochem. Biophys. Res. Commun. 2017, 491, 946–952. [Google Scholar] [CrossRef]
- Gao, Y.; Li, P.; Liu, Z.; Diao, X.; Song, C. Expression levels of vascular endothelial cell growth factor and microRNA-210 are increased in medulloblastoma and metastatic medulloblastoma. Exp. Ther. Med. 2015, 10, 2138–2144. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.; Greene, S. microRNA-10b Is Overexpressed and Critical for Cell Survival and Proliferation in Medulloblastoma. PLoS ONE 2015, 10, e0137845. [Google Scholar] [CrossRef]
- Kaid, C.; Silva, P.B.; Cortez, B.A.; Rodini, C.O.; Semedo-Kuriki, P.; Okamoto, O.K. miR-367 promotes proliferation and stem-like traits in medulloblastoma cells. Cancer Sci. 2015, 106, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Yun, Z.; Zhao, T.; Liu, X.; Ma, X. MiR-495 is a Predictive Biomarker that Downregulates GFI1 Expression in Medulloblastoma. Cell Physiol. Biochem. 2015, 36, 1430–1439. [Google Scholar] [CrossRef]
- Yogi, K.; Sridhar, E.; Goel, N.; Jalali, R.; Goel, A.; Moiyadi, A.; Thorat, R.; Panwalkar, P.; Khire, A.; Dasgupta, A.; et al. MiR-148a, a microRNA upregulated in the WNT subgroup tumors, inhibits invasion and tumorigenic potential of medulloblastoma cells by targeting Neuropilin 1. Oncoscience 2015, 2, 334–348. [Google Scholar] [CrossRef] [Green Version]
- Panwalkar, P.; Moiyadi, A.; Goel, A.; Shetty, P.; Goel, N.; Sridhar, E.; Shirsat, N. MiR-206, a Cerebellum Enriched miRNA Is Downregulated in All Medulloblastoma Subgroups and Its Overexpression Is Necessary for Growth Inhibition of Medulloblastoma Cells. J. Mol. Neurosci. 2015, 56, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Genovesi, L.A.; Carter, K.W.; Gottardo, N.G.; Giles, K.M.; Dallas, P.B. Integrated analysis of miRNA and mRNA expression in childhood medulloblastoma compared with neural stem cells. PLoS ONE 2011, 6, e23935. [Google Scholar] [CrossRef] [PubMed]
- Gokhale, A.; Kunder, R.; Goel, A.; Sarin, R.; Moiyadi, A.; Shenoy, A.; Mamidipally, C.; Noronha, S.; Kannan, S.; Shirsat, N.V. Distinctive microRNA signature of medulloblastomas associated with the WNT signaling pathway. J. Cancer Res. Ther. 2010, 6, 521–529. [Google Scholar] [PubMed]
- Lv, S.Q.; Kim, Y.H.; Giulio, F.; Shalaby, T.; Nobusawa, S.; Yang, H.; Zhou, Z.; Grotzer, M.; Ohgaki, H. Genetic alterations in microRNAs in medulloblastomas. Brain Pathol. 2012, 22, 230–239. [Google Scholar] [CrossRef]
- de Biase, D.; Acquaviva, G.; Visani, M.; Sanza, V.; Argento, C.M.; De Leo, A.; Maloberti, T.; Pession, A.; Tallini, G. Molecular Diagnostic of solid tumor using a Next Generation Sequencing custom-designed multi-gene panel. Diagnostics 2020, 10, 250. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Thomas, P.D. PANTHER in 2013: Modeling the evolution of gene function, and other gene attributes, in the context of phylogenetic trees. Nucleic Acids Res. 2013, 41, D377–D386. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef] [Green Version]
- Reimand, J.; Arak, T.; Adler, P.; Kolberg, L.; Reisberg, S.; Peterson, H.; Vilo, J. g:Profiler-a web server for functional interpretation of gene lists (2016 update). Nucleic Acids Res. 2016, 44, W83–W89. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Poschl, J.; Stark, S.; Neumann, P.; Grobner, S.; Kawauchi, D.; Jones, D.T.; Northcott, P.A.; Lichter, P.; Pfister, S.M.; Kool, M.; et al. Genomic and transcriptomic analyses match medulloblastoma mouse models to their human counterparts. Acta Neuropathol. 2014, 128, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Kool, M.; Jones, D.T.; Jager, N.; Northcott, P.A.; Pugh, T.J.; Hovestadt, V.; Piro, R.M.; Esparza, L.A.; Markant, S.L.; Remke, M.; et al. Genome sequencing of SHH medulloblastoma predicts genotype-related response to smoothened inhibition. Cancer Cell. 2014, 25, 393–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northcott, P.A.; Korshunov, A.; Witt, H.; Hielscher, T.; Eberhart, C.G.; Mack, S.; Bouffet, E.; Clifford, S.C.; Hawkins, C.E.; French, P.; et al. Medulloblastoma comprises four distinct molecular variants. J. Clin. Oncol. 2011, 29, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Kool, M.; Korshunov, A.; Remke, M.; Jones, D.T.; Schlanstein, M.; Northcott, P.A.; Cho, Y.J.; Koster, J.; Schouten-van Meeteren, A.; van Vuurden, D.; et al. Molecular subgroups of medulloblastoma: An international meta-analysis of transcriptome, genetic aberrations, and clinical data of WNT, SHH, Group 3, and Group 4 medulloblastomas. Acta Neuropathol. 2012, 123, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.D.; Northcott, P.A.; Korshunov, A.; Remke, M.; Cho, Y.J.; Clifford, S.C.; Eberhart, C.G.; Parsons, D.W.; Rutkowski, S.; Gajjar, A.; et al. Molecular subgroups of medulloblastoma: The current consensus. Acta Neuropathol. 2012, 123, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Northcott, P.A.; Hielscher, T.; Dubuc, A.; Mack, S.; Shih, D.; Remke, M.; Al-Halabi, H.; Albrecht, S.; Jabado, N.; Eberhart, C.G.; et al. Pediatric and adult sonic hedgehog medulloblastomas are clinically and molecularly distinct. Acta Neuropathol. 2011, 122, 231–240. [Google Scholar] [CrossRef]
- Ellison, D.W.; Kocak, M.; Dalton, J.; Megahed, H.; Lusher, M.E.; Ryan, S.L.; Zhao, W.; Nicholson, S.L.; Taylor, R.E.; Bailey, S.; et al. Definition of disease-risk stratification groups in childhood medulloblastoma using combined clinical, pathologic, and molecular variables. J. Clin. Oncol. 2011, 29, 1400–1407. [Google Scholar] [CrossRef] [Green Version]
- Christopher, A.F.; Kaur, R.P.; Kaur, G.; Kaur, A.; Gupta, V.; Bansal, P. MicroRNA therapeutics: Discovering novel targets and developing specific therapy. Perspect Clin. Res. 2016, 7, 68–74. [Google Scholar] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Doss, C.G.P.; Lee, S.S. Therapeutic miRNA and siRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol. Ther. Nucleic Acids 2017, 8, 132–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visani, M.; de Biase, D.; Marucci, G.; Taccioli, C.; Baruzzi, A.; Pession, A. Definition of miRNAs expression profile in glioblastoma samples: The relevance of non-neoplastic brain reference. PLoS ONE 2013, 8, e55314. [Google Scholar] [CrossRef] [PubMed]
- McGlinn, E.; Yekta, S.; Mansfield, J.H.; Soutschek, J.; Bartel, D.P.; Tabin, C.J. In ovo application of antagomiRs indicates a role for miR-196 in patterning the chick axial skeleton through Hox gene regulation. Proc. Natl. Acad. Sci. USA 2009, 106, 18610–18615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.; Yan, W.; Zhang, G.; Lv, H.; Liu, Z.; Fang, F.; Zhang, W.; Zhang, J.; Tao, T.; You, Y.; et al. Upregulation of miR-196b confers a poor prognosis in glioblastoma patients via inducing a proliferative phenotype. PLoS ONE 2012, 7, e38096. [Google Scholar] [CrossRef]
- Coskun, E.; von der Heide, E.K.; Schlee, C.; Kuhnl, A.; Gokbuget, N.; Hoelzer, D.; Hofmann, W.K.; Thiel, E.; Baldus, C.D. The role of microRNA-196a and microRNA-196b as ERG regulators in acute myeloid leukemia and acute T-lymphoblastic leukemia. Leuk. Res. 2011, 35, 208–213. [Google Scholar] [CrossRef]
- Lim, J.Y.; Yoon, S.O.; Seol, S.Y.; Hong, S.W.; Kim, J.W.; Choi, S.H.; Lee, J.S.; Cho, J.Y. Overexpression of miR-196b and HOXA10 characterize a poor-prognosis gastric cancer subtype. World J. Gastroenterol. 2013, 19, 7078–7088. [Google Scholar] [CrossRef]
- Kanno, S.; Nosho, K.; Ishigami, K.; Yamamoto, I.; Koide, H.; Kurihara, H.; Mitsuhashi, K.; Shitani, M.; Motoya, M.; Sasaki, S.; et al. MicroRNA-196b is an independent prognostic biomarker in patients with pancreatic cancer. Carcinogenesis 2017, 38, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Ge, J.; Chen, Z.; Li, R.; Lu, T.; Xiao, G. Upregulation of microRNA-196a and microRNA-196b cooperatively correlate with aggressive progression and unfavorable prognosis in patients with colorectal cancer. Cancer Cell Int. 2014, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Mo, J.S.; Alam, K.J.; Kang, I.H.; Park, W.C.; Seo, G.S.; Choi, S.C.; Kim, H.S.; Moon, H.B.; Yun, K.J.; Chae, S.C. MicroRNA 196B regulates FAS-mediated apoptosis in colorectal cancer cells. Oncotarget 2015, 6, 2843–2855. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Lin, B.; Zhang, X.; Peng, Y.; Ye, Z.; Ma, Y.; Liang, Y.; Cao, L.; Li, X.; Li, R.; et al. Maintenance of cancer stemness by miR-196b-5p contributes to chemoresistance of colorectal cancer cells via activating STAT3 signaling pathway. Oncotarget 2017, 8, 49807–49823. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.C.; Chang, J.T.; Liao, C.T.; Kang, C.J.; Huang, S.F.; Chen, I.H.; Huang, C.C.; Huang, Y.C.; Chen, W.H.; Tsai, C.Y.; et al. OncomiR-196 promotes an invasive phenotype in oral cancer through the NME4-JNK-TIMP1-MMP signaling pathway. Mol. Cancer 2014, 13, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, D.; Hu, L.; Lei, D.; Fang, X.; Zhang, Z.; Wang, T.; Lin, M.; Huang, J.; Yang, H.; Zhou, X.; et al. MicroRNA-196b promotes cell proliferation and suppress cell differentiation in vitro. Biochem. Biophys. Res. Commun. 2015, 457, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Abe, W.; Nasu, K.; Nakada, C.; Kawano, Y.; Moriyama, M.; Narahara, H. miR-196b targets c-myc and Bcl-2 expression, inhibits proliferation and induces apoptosis in endometriotic stromal cells. Hum. Reprod. 2013, 28, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Wei, J.; You, W.; Wang, R.; Shang, J.; Xiong, Y.; Yang, H.; Yang, X.; Fu, Z. Disruption of the c-Myc/miR-200b-3p/PRDX2 regulatory loop enhances tumor metastasis and chemotherapeutic resistance in colorectal cancer. J. Transl. Med. 2017, 15, 257. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Hu, J.; Shen, Z.; Yao, R.; Liu, S.; Li, Y.; Cong, H.; Wang, X.; Qiu, W.; Yue, L. MiR-200b expression in breast cancer: A prognostic marker and act on cell proliferation and apoptosis by targeting Sp1. J. Cell Mol. Med. 2015, 19, 760–769. [Google Scholar] [CrossRef]
- Ye, F.; Tang, H.; Liu, Q.; Xie, X.; Wu, M.; Liu, X.; Chen, B. miR-200b as a prognostic factor in breast cancer targets multiple members of RAB family. J. Transl. Med. 2014, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.; Yu, H.; Yuan, J.; Guo, C.; Cao, H.; Li, W.; Xiao, C. MicroRNA-200b Impacts Breast Cancer Cell Migration and Invasion by Regulating Ezrin-Radixin-Moesin. Med. Sci. Monit. 2016, 22, 1946–1952. [Google Scholar] [CrossRef]
- Li, D.; Wang, H.; Song, H.; Xu, H.; Zhao, B.; Wu, C.; Hu, J.; Wu, T.; Xie, D.; Zhao, J.; et al. The microRNAs miR-200b-3p and miR-429-5p target the LIMK1/CFL1 pathway to inhibit growth and motility of breast cancer cells. Oncotarget 2017, 8, 85276–85289. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Wang, G.; Wang, Z.; An, S.; Ye, P.; Luo, S. A negative feedback loop between miR-200b and the nuclear factor-kappaB pathway via IKBKB/IKK-beta in breast cancer cells. FEBS J. 2016, 283, 2259–2271. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.V.; Martin, E.C.; Segar, H.C.; Miller, D.F.; Buechlein, A.; Rusch, D.B.; Nephew, K.P.; Burow, M.E.; Collins-Burow, B.M. Dual regulation by microRNA-200b-3p and microRNA-200b-5p in the inhibition of epithelial-to-mesenchymal transition in triple-negative breast cancer. Oncotarget 2015, 6, 16638–16652. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Ding, X.; Li, X.; Gao, S.; Yang, Q. 53BP1 suppresses epithelial-mesenchymal transition by downregulating ZEB1 through microRNA-200b/429 in breast cancer. Cancer Sci. 2015, 106, 982–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Cao, Y.; Wang, P.; Song, H.; Bie, T.; Li, M.; Huai. miR-200b-3p in plasma is a potential diagnostic biomarker in oral squamous cell carcinoma. Biomarkers 2018, 23, 137–141. [Google Scholar] [CrossRef]
- Vosa, U.; Vooder, T.; Kolde, R.; Vilo, J.; Metspalu, A.; Annilo, T. Meta-analysis of microRNA expression in lung cancer. Int. J. Cancer 2013, 132, 2884–2893. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Yin, Z.; Li, X.; Wu, W.; Zhou, B. Meta-analysis of human lung cancer microRNA expression profiling studies comparing cancer tissues with normal tissues. J. Exp. Clin. Cancer Res. 2012, 31, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Todd, N.W.; Xing, L.; Xie, Y.; Zhang, H.; Liu, Z.; Fang, H.; Zhang, J.; Katz, R.L.; Jiang, F. Early detection of lung adenocarcinoma in sputum by a panel of microRNA markers. Int. J. Cancer 2010, 127, 2870–2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bont, J.M.; Packer, R.J.; Michiels, E.M.; den Boer, M.L.; Pieters, R. Biological background of pediatric medulloblastoma and ependymoma: A review from a translational research perspective. Neuro Oncol. 2008, 10, 1040–1060. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, W.; Koch, A.; Brune, H.; Waha, A.; Schuller, U.; Dani, I.; Denkhaus, D.; Langmann, W.; Bode, U.; Wiestler, O.D.; et al. Insulin-like growth factor II is involved in the proliferation control of medulloblastoma and its cerebellar precursor cells. Am. J. Pathol. 2005, 166, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Borowska, A.; Jozwiak, J. Medulloblastoma: Molecular pathways and histopathological classification. Arch. Med. Sci. 2016, 12, 659–666. [Google Scholar] [CrossRef] [Green Version]
- Sandberg, A.A.; Stone, J.F. The Genetics and Molecular Biology of Neural Tumors; Humana Press: Totowa, NJ, USA, 2008. [Google Scholar]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, T.J.; Brown, K.M.; LaFleur, B.; Peterson, K.; Lawlor, C.; Chen, Y.; Packer, R.J.; Cogen, P.; Stephan, D.A. Expression profiling of medulloblastoma: PDGFRA and the RAS/MAPK pathway as therapeutic targets for metastatic disease. Nat. Genet. 2001, 29, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Park, A.K.; Lee, J.Y.; Cheong, H.; Ramaswamy, V.; Park, S.H.; Kool, M.; Phi, J.H.; Choi, S.A.; Cavalli, F.; Taylor, M.D.; et al. Subgroup-specific prognostic signaling and metabolic pathways in pediatric medulloblastoma. BMC Cancer 2019, 19, 571. [Google Scholar] [CrossRef] [PubMed]
- Kieran, M.W.; Chisholm, J.; Casanova, M.; Brandes, A.A.; Aerts, I.; Bouffet, E.; Bailey, S.; Leary, S.; MacDonald, T.J.; Mechinaud, F.; et al. Phase I study of oral sonidegib (LDE225) in pediatric brain and solid tumors and a phase II study in children and adults with relapsed medulloblastoma. Neuro Oncol. 2017, 19, 1542–1552. [Google Scholar] [CrossRef] [PubMed]
- Meiss, F.; Andrlova, H.; Zeiser, R. Vismodegib. Recent Results Cancer Res. 2018, 211, 125–139. [Google Scholar]
- Robinson, G.W.; Orr, B.A.; Wu, G.; Gururangan, S.; Lin, T.; Qaddoumi, I.; Packer, R.J.; Goldman, S.; Prados, M.D.; Desjardins, A.; et al. Vismodegib Exerts Targeted Efficacy Against Recurrent Sonic Hedgehog-Subgroup Medulloblastoma: Results From Phase II Pediatric Brain Tumor Consortium Studies PBTC-025B and PBTC-032. J. Clin. Oncol. 2015, 33, 2646–2654. [Google Scholar] [CrossRef]
- Lou, E.; Schomaker, M.; Wilson, J.D.; Ahrens, M.; Dolan, M.; Nelson, A.C. Complete and sustained response of adult medulloblastoma to first-line sonic hedgehog inhibition with vismodegib. Cancer Biol. Ther. 2016, 17, 1010–1016. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Song, Q.; Day, B.W. Phase I and phase II sonidegib and vismodegib clinical trials for the treatment of paediatric and adult MB patients: A systemic review and meta-analysis. Acta Neuropathol. Commun. 2019, 7, 123. [Google Scholar] [CrossRef]
- Zhao, X.; Ponomaryov, T.; Ornell, K.J.; Zhou, P.; Dabral, S.K.; Pak, E.; Li, W.; Atwood, S.X.; Whitson, R.J.; Chang, A.L.; et al. RAS/MAPK Activation Drives Resistance to Smo Inhibition, Metastasis, and Tumor Evolution in Shh Pathway-Dependent Tumors. Cancer Res. 2015, 75, 3623–3635. [Google Scholar] [CrossRef] [Green Version]
- Gilbertson, R.J.; Langdon, J.A.; Hollander, A.; Hernan, R.; Hogg, T.L.; Gajjar, A.; Fuller, C.; Clifford, S.C. Mutational analysis of PDGFR-RAS/MAPK pathway activation in childhood medulloblastoma. Eur. J. Cancer 2006, 42, 646–649. [Google Scholar] [CrossRef]
- Schuller, U.; Schober, F.; Kretzschmar, H.A.; Herms, J. Bcl-2 expression inversely correlates with tumour cell differentiation in medulloblastoma. Neuropathol. Appl. Neurobiol. 2004, 30, 513–521. [Google Scholar] [CrossRef]
- Mancarella, C.; Scotlandi, K. IGF2BP3 From Physiology to Cancer: Novel Discoveries, Unsolved Issues, and Future Perspectives. Front. Cell Dev. Biol. 2019, 7, 363. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, R.G.; Gonda, T.J. MYB function in normal and cancer cells. Nat. Rev. Cancer 2008, 8, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Lassaletta, A.; Ramaswamy, V. Medulloblastoma in adults: They’re not just big kids. Neuro Oncol. 2016, 18, 895–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamm, G.M.; Christofori, G. Impairment of survival factor function potentiates chemotherapy-induced apoptosis in tumor cells. Cancer Res. 1998, 58, 801–807. [Google Scholar]
- Call, J.A.; Naik, M.; Rodriguez, F.J.; Giannini, C.; Wu, W.; Buckner, J.C.; Parney, I.F.; Laack, N.N. Long-term outcomes and role of chemotherapy in adults with newly diagnosed medulloblastoma. Am. J. Clin. Oncol. 2014, 37, 1–7. [Google Scholar] [CrossRef]
- Brandes, A.A.; Ermani, M.; Amista, P.; Basso, U.; Vastola, F.; Gardiman, M.; Iuzzolino, P.; Turazzi, S.; Rotilio, A.; Volpin, L.; et al. The treatment of adults with medulloblastoma: A prospective study. Int. J. Radiat. Oncol. Biol. Phys. 2003, 57, 755–761. [Google Scholar] [CrossRef]
- Riffaud, L.; Saikali, S.; Leray, E.; Hamlat, A.; Haegelen, C.; Vauleon, E.; Lesimple, T. Survival and prognostic factors in a series of adults with medulloblastomas. J. Neurosurg. 2009, 111, 478–487. [Google Scholar] [CrossRef]
- Zhao, F.; Ohgaki, H.; Xu, L.; Giangaspero, F.; Li, C.; Li, P.; Yang, Z.; Wang, B.; Wang, X.; Wang, Z.; et al. Molecular subgroups of adult medulloblastoma: A long-term single-institution study. Neuro Oncol. 2016, 18, 982–990. [Google Scholar] [CrossRef]
- Ramaswamy, V.; Remke, M.; Bouffet, E.; Bailey, S.; Clifford, S.C.; Doz, F.; Kool, M.; Dufour, C.; Vassal, G.; Milde, T.; et al. Risk stratification of childhood medulloblastoma in the molecular era: The current consensus. Acta Neuropathol. 2016, 131, 821–831. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA Therapeutics in Cancer—An Emerging Concept. EBio Med. 2016, 12, 34–42. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEO Accession Number | Number Samples (Children/Adult) | Platform |
|---|---|---|
| GSE41842 | 19 (15/4) | Affymetrix Human Gene 1.0 ST Array (transcript (gene) version) |
| GSE49243 | 71 (26/45) | Affymetrix Human Genome U133 Plus 2.0 Array |
| GSE21140 | 102 (89/13) | Affymetrix Human Exon 1.0 ST Array (transcript (gene) version] |
| AD | CH | |||
|---|---|---|---|---|
| Mean age | 30.48 | 6.58 | ||
| Range | 16–66 | 1–15 | ||
| Sex ratio M:F | 14:7 | 12:7 | ||
| Histological subtype | ||||
| No. | % | No. | % | |
| CL | 9 | 42.86% | 12 | 63.16% |
| D/N | 9 | 42.86% | 4 | 21.05% |
| LCA | 3 | 14.28% | 3 | 15.79% |
| Molecular subtype | ||||
| No. | % | No. | % | |
| WNT | 4 | 19.05% | 4 | 21.05% |
| SHH | 7 | 33.33% | 3 | 15.79% |
| non-WNT/non-SHH | 10 | 47.62% | 12 | 63.16% |
| miRNA Upregulated | |||||
| F.C. | F.C. | F.C. | |||
| miR-196b-5p | 77.891 | miR-221-3p | 3.613 | miR-485-3p | 2.194 |
| miR-200b-3p | 11.904 | miR-376b-3p | 3.613 | miR-888-5p | 2.134 |
| miR-193a-3p | 10.080 | miR-299-5p | 3.539 | miR-296-5p | 2.119 |
| miR-376a-3p | 8.187 | let-7f-5p | 3.515 | miR-29b-3p | 2.119 |
| miR-451a | 8.075 | miR-539-5p | 3.490 | miR-345-5p | 2.119 |
| miR-452-5p | 7.534 | miR-346 | 3.348 | let-7b-5p | 2.104 |
| miR-190a | 7.328 | miR-223-3p | 3.302 | miR-302a-3p | 2.090 |
| miR-590-5p | 7.127 | miR-9-3p | 3.302 | miR-411-5p | 2.075 |
| miR-625-3p | 6.513 | let-7i-5p | 3.212 | miR-20a-5p | 2.061 |
| miR-27a-3p | 6.379 | miR-24-3p | 3.146 | miR-140-5p | 2.047 |
| miR-142-3p | 6.035 | miR-146b-5p | 3.124 | miR-335-5p | 2.047 |
| miR-374b-5p | 5.911 | miR-153 | 3.102 | miR-98-5p | 2.047 |
| miR-27b-3p | 5.670 | miR-28-5p | 2.875 | miR-660-5p | 2.033 |
| miR-376c-3p | 5.515 | miR-127-3p | 2.855 | ||
| miR-21-3p | 5.364 | miR-136-5p | 2.777 | ||
| miR-100-5p | 5.290 | miR-582-5p | 2.720 | ||
| miR-31-5p | 5.075 | miR-92b-3p | 2.701 | ||
| miR-374a-5p | 5.040 | miR-20b-3p | 2.609 | ||
| miR-365a-3p | 4.835 | miR-195-5p | 2.555 | ||
| miR-193a-5p | 4.702 | miR-323a-3p | 2.434 | ||
| miR-135a-5p | 4.670 | let-7d-3p | 2.401 | ||
| miR-363-3p | 4.574 | miR-627 | 2.351 | ||
| miR-137 | 4.511 | let-7d-5p | 2.303 | ||
| miR-671-5p | 4.357 | miR-424-5p | 2.287 | ||
| miR-33a-5p | 3.954 | miR-497-5p | 2.287 | ||
| miRNA Downregulated | |||||
| F.C. | F.C. | F.C. | |||
| miR-183-5p | −70.356 | miR-665 | −3.379 | miR-184 | −2.066 |
| miR-642a-5p | −53.320 | miR-449a | −3.356 | miR-339-5p | −2.051 |
| miR-182-5p | −43.010 | miR-455-5p | −3.356 | miR-133b | −2.037 |
| miR-196a-5p | −33.281 | miR-873-5p | −3.287 | ||
| miR-96-5p | −29.377 | miR-181a-3p | −3.197 | ||
| miR-206 | −20.629 | miR-890 | −3.109 | ||
| miR-760 | −10.979 | miR-135b-5p | −2.983 | ||
| miR-181b-5p | −10.827 | miR-181d | −2.764 | ||
| miR-877-5p | −8.554 | miR-595 | −2.633 | ||
| miR-124-3p | −7.344 | miR-662 | −2.615 | ||
| miR-506-3p | −5.965 | miR-10b-5p | −2.561 | ||
| miR-181a-5p | −5.843 | miR-211-5p | −2.508 | ||
| miR-490-3p | −4.845 | miR-187-5p | −2.457 | ||
| miR-373-5p | −4.616 | miR-187-3p | −2.423 | ||
| miR-302c-5p | −4.277 | miR-454-3p | −2.356 | ||
| miR-542-5p | −4.277 | miR-301a-3p | −2.308 | ||
| miR-200c-3p | −4.074 | miR-421 | −2.260 | ||
| miR-203a | −3.991 | miR-155-5p | −2.245 | ||
| miR-501-5p | −3.855 | miR-324-3p | −2.245 | ||
| miR-488-3p | −3.802 | miR-520c-3p | −2.229 | ||
| miR-138-5p | −3.749 | miR-34b-3p | −2.214 | ||
| miR-324-5p | −3.698 | miR-20b-5p | −2.199 | ||
| miR-141-3p | −3.672 | miR-765 | −2.168 | ||
| miR-29b-2-5p | −3.622 | miR-126-5p | −2.139 | ||
| miR-628-3p | −3.523 | miR-152 | −2.139 | ||
| PANTHER Pathways | Number of Genes | |||||
|---|---|---|---|---|---|---|
| Reference List * | Target List † | Fold Enrichment Scores | p-Value | miR-196b-5p Target Genes | miR-200b-3p Target Genes | |
| Insulin/IGF pathway-mitogen activated protein kinase kinase/MAP kinase cascade | 32 | 5 | 9.84 | 2.84 × 10−2 | MAPK1, FOS | IGF2, M6PR, IRS1 |
| Ras Pathway | 71 | 11 | 9.76 | 4.58 × 10−6 | MAPK1, AKT1, PIK3CG, RGL2 | KRAS, RHOA, SHC1, JUN, ETS1, PDPK1, PAK2 |
| Insulin/IGF pathway-protein kinase B signaling cascade | 41 | 6 | 9.22 | 9.18 × 10−3 | AKT1, PIK3CG | IGF2, M6PR, PDPK1, IRS1 |
| Interleukin signaling pathway | 83 | 12 | 9.11 | 2.28 × 10−6 | MAPK1, AKT1, MTOR, MYC, FOS, CDKN1B | IKBKB, SHC1, MKNK2, PDPK1, IRS1, CDKN1B, MAPK7 |
| VEGF signaling pathway | 65 | 9 | 8.72 | 2.09 × 10−4 | MAPK1, AKT1, PIK3CG, PRKACA | KDR, PKD1, ETS1, VEGFA, PLCG1 |
| Apoptosis signaling pathway | 119 | 16 | 8.47 | 2.52 × 10−8 | MAPK1, FAS, AKT1, PIK3CG, ATF4, PRKACA, BCL2, FOS | IKBKB, M6PR, CREB1, KLHL20, JUN, BAG4, XIAP, BCL2, BCL2L11 |
| B cell activation | 65 | 8 | 7.75 | 1.88 × 10−3 | MAPK1, MAP3K2, PIK3CG, CALM1, FOS | IKBKB, JUN, RHOA |
| p53 pathway feedback loops 2 | 51 | 6 | 7.41 | 2.98 × 10−2 | AKT1, PIK3CG, MYC | KRAS, E2F3, PDPK1 |
| Angiogenesis | 161 | 18 | 7.04 | 3.36 × 10−8 | MAPK1, AKT1, PIK3CG, PRKACA, FOS | PKD1, RHOA, JUN, FRS2, CRKL, KDR, SHC1, ETS1, WNT1, TCF7L2, PAK2, VEGFA, PLCG1 |
| CCKR signaling map | 162 | 17 | 7.00 | 3.71 × 10−8 | MAPK1, AKT1, PRKACA, CALM1, MYC, BCL2, FOS | CREB1, PKD1, RHOA, SHC1, JUN, PDPK1, IRS1, MAPK7, BCL2, SP1, PLCG1 |
| PDGF signaling pathway | 144 | 16 | 7.00 | 3.73 × 10−7 | MAPK1, MAP3K2, MTOR, PIK3CG, MYC, PRKACA, FOS | IKBKB, DLC1, SHC1, JUN, ETS1, MKNK2, PDPK1, MAPK7, PLCG1 |
| T cell activation | 85 | 9 | 6.67 | 1.77 × 10−3 | MAPK1, AKT1, PIK3CG, CALM1, FOS | IKBKB, JUN, PAK2, PLCG1 |
| p53 pathway | 88 | 8 | 5.73 | 1.56 × 10−2 | FAS, AKT1, PIK3CG | E2F3, PMAIP1, EP300, ZMAT3, PDPK1 |
| Integrin signalling pathway | 162 | 12 | 5.44 | 7.61 × 10−5 | MAPK1, MAP3K2, LAMB2, PIK3CG | CRKL, FN1, PTPN12, RND3, RHOA, SHC1, ARPC3, RAP2C |
| Gonadotropin-releasing hormone receptor pathway | 227 | 18 | 5.27 | 1.14 × 10−6 | MAPK1, MAP3K2, LHX2, ACVR2B, AKT1, TGFBR3, PBX1, PRKACA, FOS | CREB1, GATA4, ZEB1, SMAD2, EP300, JUN, TCF7L1, SP1, PPARA |
| EGF receptor signaling pathway | 135 | 11 | 5.13 | 2.20 × 10−3 | MAPK1, MAP3K2, AKT1, PIK3CG, PRKACA | BTC, PKD1, RHOA, SHC1, MAPK7, PLCG1 |
| FGF signaling pathway | 120 | 9 | 4.73 | 2.44 × 10−2 | MAPK1, MAP3K2, AKT1, PIK3CG, PRKACA | FRS2, RHOA, SHC1, PLCG1 |
| Inflammation mediated by chemokine and cytokine signaling pathway | 230 | 13 | 3.56 | 1.59 × 10−2 | MAPK1, AKT1, PRKACA, PIK3CG | IKBKB, KRAS, RHOA, JUN, PLCG1, SHC1, ARPC3, PDPK1, PAK2 |
| Index | Name | Overlap | miR-196b-5p Target Genes | miR-200b-3p Target Genes |
|---|---|---|---|---|
| 1 | Colorectal cancer | 16/86 | FOS, MTOR, TGFBR2, MSH6, MYC, BCL2, AKT1, MAPK1 | SMAD2, TCF7L2, JUN, TCF7L1, RHOA, BCL2L11, BCL2, KRAS, PMAIP1 |
| 2 | Prostate cancer | 17/97 | CDKN1B, MTOR, IKBKB, BCL2, AKT1, MAPK1, ATF4 | TCF7L2, TCF7L1, CDKN1B, PDPK1, ETV5, CREB1, ZEB1, CCNE2, BCL2, EP300, E2F3, KRAS |
| 3 | Neurotrophin signaling pathway | 18/119 | BLC2, AKT1, MAPK1, CALM3, CALM1, ATF4 | JUN, SHC1, IRS1, PDPK1, FRS2, RHOA, CRKL, IKBKB, MAPK7, ARHGDIA, BCL2, KRAS, PLCG1 |
| 4 | MicroRNAs in cancer | 32/299 | CDKN1B, MYC, IGFBP1, MAPK1, RDX, HMGA2, MTOR, BLC2 | DNMT1, CDKN1B, NOTCH1, IRS1, SHC1, BMI1, CRKL, IKBKB, BCL2L11, MAPK7, DNMT3B, EP300, E2F3, PLCG1, DNMT3A, RHOA, VEGFA, ZEB2, ZEB1, CCNE2, DDIT4, FSCN1, BCL2, KRAS, ZFPM2, EZH2 |
| 5 | Proteoglycans in cancer | 21/201 | RDX, MTOR, MYC, AKT1, MAPK1, FAS, PRKACA | SMAD2, PDPK1, ROCK2, IGF2, FN1, MSN, FRS2, WNT16, RHOA, VEGFA, KDR, KRAS, PLCG1, WNT1 |
| 6 | ErbB signaling pathway | 12/85 | CDKN1B, MYC, AKT1, MAPK1, MTOR | BTC, JUN, CDKN1B, SHC1, KRAS, PLCG1, PAK2, CRKL |
| 7 | Pathways in cancer | 39/530 | CDKN1B, MYC, AKT1, MAPK1, PRKACA, LAMB2, FOS, MTOR, TGFBR2, MSH6, BCL2, FAS, CALM3, CALM1 | CDKN1B, NOTCH1, BCL2L11, KRAS, CRKL, IKBKB, ROCK2, XIAP, ETS1, EP300, PMAIP1, E2F3, PLCG1, ELOC, WNT1, SMAD2, TCF7L2, JUN, TCF7L1, FN1, IGF2, WNT16, RHOA, VEGFA, CCNE2, SP1, BCL2 |
| 8 | HIF-1 signaling pathway | 13/100 | CDKN1B, MTOR, BCL2, AKT1, MAPK1, ALDOA | FLT1, CDKN1B, VEGFA, LDHA, MKNK2, BCL2, EP300, PLCG1, ELOC |
| 9 | Apoptosis | 16/143 | DFFA, FOS, BCL2, AKT1, MAPK1, FAS, ATF4 | JUN, PDPK1, XIAP, LMNB1, IKBKB, BCL2L11, BCL2, PMAIP1, CASP2, KRAS |
| 10 | Renal cell carcinoma | 10/69 | AKT1, MAPK1 | JUN, EP300, KRAS, ELOC, ETS1, PAK2, CRKL, VEGFA |
| DataSets | Number of Downregulated Genes in Ad vs. Ch Mbs | Downregulated Genes Targeted by miR-196b-5p or miR-200b-3p | KEGG 2019 Human Pathway |
|---|---|---|---|
| GSE49243 | 404 | N = 10 | |
| IGF2BP3 | - | ||
| ETS1 | Pathways in cancer, Renal cell carcinoma, Human T-cell leukemia virus 1 infection, Ras signaling pathway, Cellular senescence | ||
| PMAIP1 | Pathways in cancer, p53 signaling pathway, Colorectal cancer, Apoptosis, Viral carcinogenesis | ||
| ZFPM2 | MicroRNAs in cancer | ||
| LRIG3 | - | ||
| CD8A | Primary immunodeficiency, Antigen processing and presentation, Hematopoietic cell lineage, T cell receptor signaling pathway, Cell adhesion molecules (CAMs) | ||
| OXR1 | - | ||
| FKBP5 | Estrogen signaling pathway | ||
| TPD52L1 | - | ||
| MYB | PI3K-Akt signaling pathway | ||
| GSE41842 | 105 | N = 4 | |
| LMNB1 | Apoptosis | ||
| DCBLD2 | - | ||
| CTPS1 | Pyrimidine metabolism | ||
| E2F3 | Bladder cancer, Non-small cell lung cancer, Melanoma, Pancreatic cancer, Glioma, Chronic myeloid leukemia, Small cell lung cancer, Prostate cancer, Cell cycle, Breast cancer, Gastric cancer, Cushing syndrome, Hepatitis C, Cellular senescence, Hepatitis B, Hepatocellular carcinoma, Kaposi sarcoma-associated herpesvirus infection, Epstein-Barr virus infection, Human T-cell leukemia virus 1 infection, Human cytomegalovirus infection, MicroRNAs in cancer, Pathways in cancer | ||
| GSE21140 | 622 | N = 13 | |
| KCNJ2 | Cholinergic synapse, Oxytocin signaling pathway, Renin secretion, Gastric acid secretion | ||
| KDR | Fluid shear stress and atherosclerosis, MAPK signaling pathway, PI3K-Akt signaling pathway, VEGF signaling pathway, Focal adhesion, Proteoglycans in cancer, Rap1 signaling pathway, Ras signaling pathway | ||
| IGF2BP3 | - | ||
| HERC2 | Ubiquitin mediated proteolysis | ||
| FOS | Cholinergic synapse, Fluid shear stress and atherosclerosis, Oxytocin signaling pathway, MAPK signaling pathway, Amphetamine addiction, Prolactin signaling pathway, B cell receptor signaling pathway, Leishmaniasis, Pertussis, Salmonella infection, Colorectal cancer, Rheumatoid arthritis, Th1 and Th2 cell differentiation, IL-17 signaling pathway, Circadian entrainment, Choline metabolism in cancer, T cell receptor signaling pathway, Chagas disease (American trypanosomiasis), Toll-like receptor signaling pathway, Parathyroid hormone synthesis secretion and action, Th17 cell differentiation, TNF signaling pathway, Osteoclast differentiation, Relaxin signaling pathway, Dopaminergic synapse, Estrogen signaling pathway, Measles, Apoptosis, Breast cancer, Hepatitis B, Kaposi sarcoma-associated herpesvirus infection, Human immunodeficiency virus 1 infection, cAMP signaling pathway, Human T-cell leukemia virus 1 infection, Pathways in cancer | ||
| ARID5B | Protein processing in endoplasmic reticulum | ||
| HSPA4L | - | ||
| ZFHX4 | - | ||
| DLC1 | - | ||
| TRIM56 | - | ||
| MTUS1 | - | ||
| FOXG1 | FoxO signaling pathway | ||
| MYB | PI3K-Akt signaling pathway |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visani, M.; Marucci, G.; de Biase, D.; Giangaspero, F.; Buttarelli, F.R.; Brandes, A.A.; Franceschi, E.; Acquaviva, G.; Ciarrocchi, A.; Rhoden, K.J.; et al. miR-196B-5P and miR-200B-3P Are Differentially Expressed in Medulloblastomas of Adults and Children. Diagnostics 2020, 10, 265. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10050265
Visani M, Marucci G, de Biase D, Giangaspero F, Buttarelli FR, Brandes AA, Franceschi E, Acquaviva G, Ciarrocchi A, Rhoden KJ, et al. miR-196B-5P and miR-200B-3P Are Differentially Expressed in Medulloblastomas of Adults and Children. Diagnostics. 2020; 10(5):265. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10050265
Chicago/Turabian StyleVisani, Michela, Gianluca Marucci, Dario de Biase, Felice Giangaspero, Francesca Romana Buttarelli, Alba Ariela Brandes, Enrico Franceschi, Giorgia Acquaviva, Alessia Ciarrocchi, Kerry Jane Rhoden, and et al. 2020. "miR-196B-5P and miR-200B-3P Are Differentially Expressed in Medulloblastomas of Adults and Children" Diagnostics 10, no. 5: 265. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10050265