
Sphingomonas and Phenylobacterium as Major Microbiota in Thymic Epithelial Tumors

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. Patient Characteristics
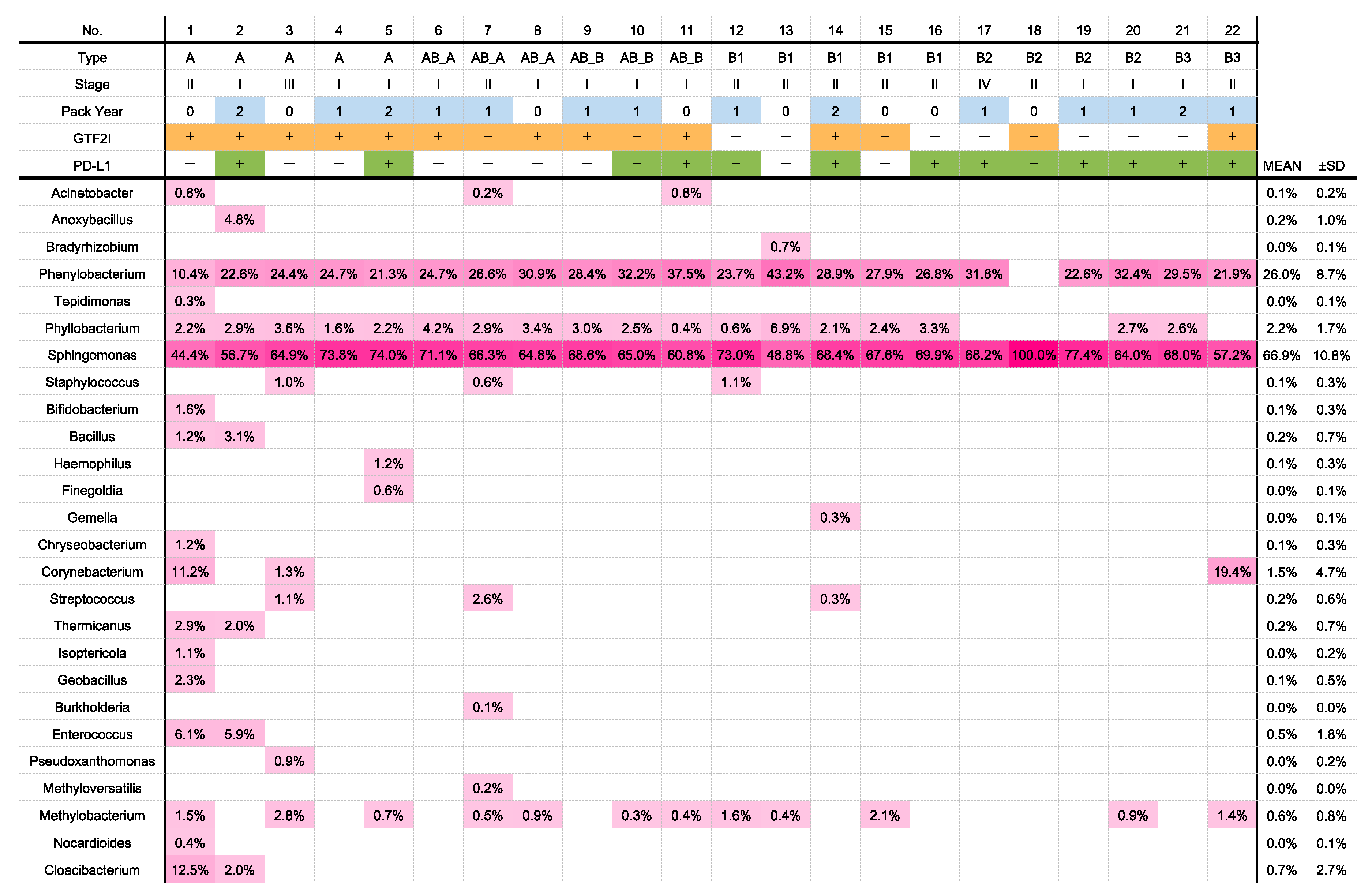
2.2. OTU Analyses
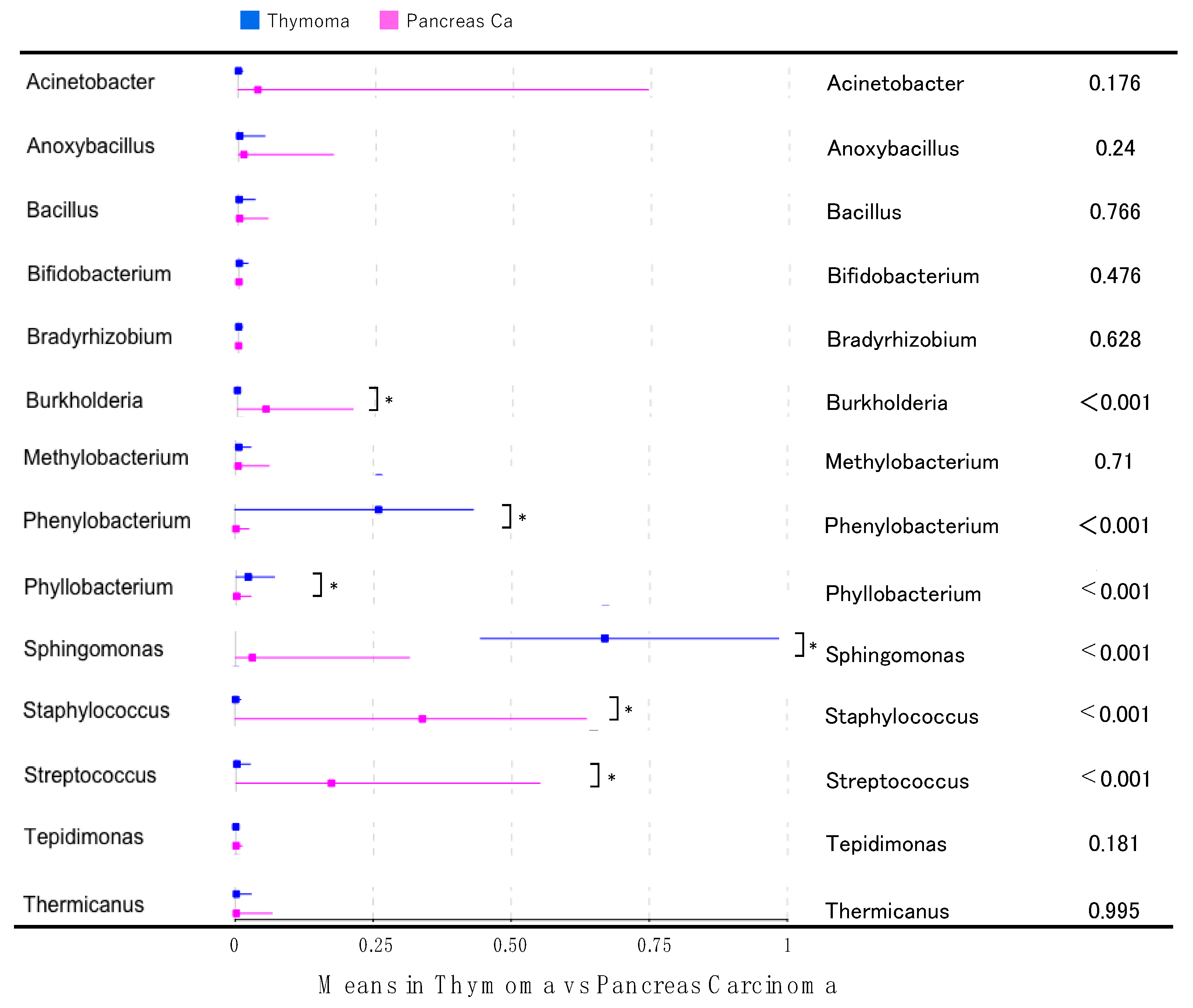
2.3. Differences in Microbiota between Thymomas and Pancreatic Cancers
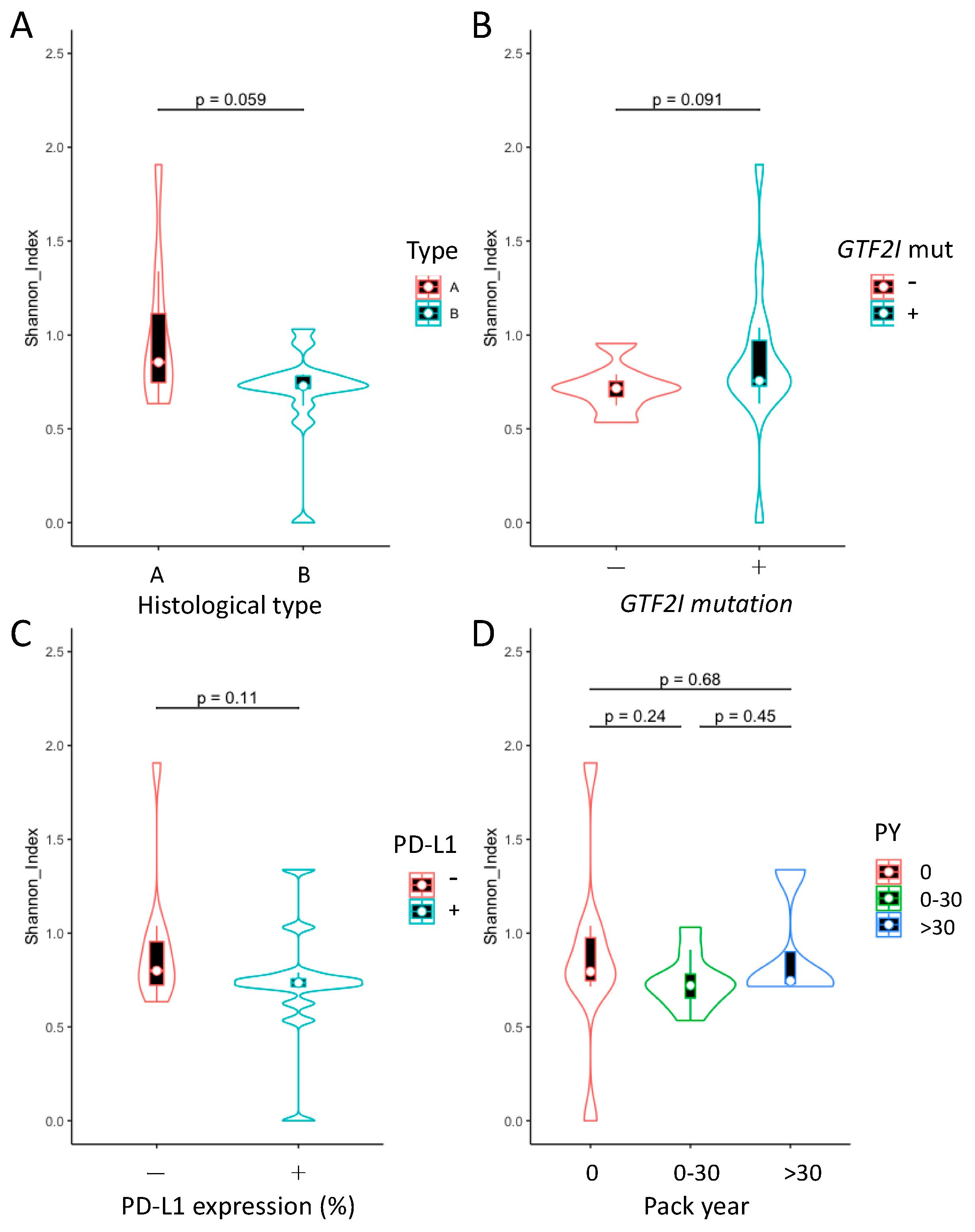
2.4. Analysis of Microbial Diversity within Groups
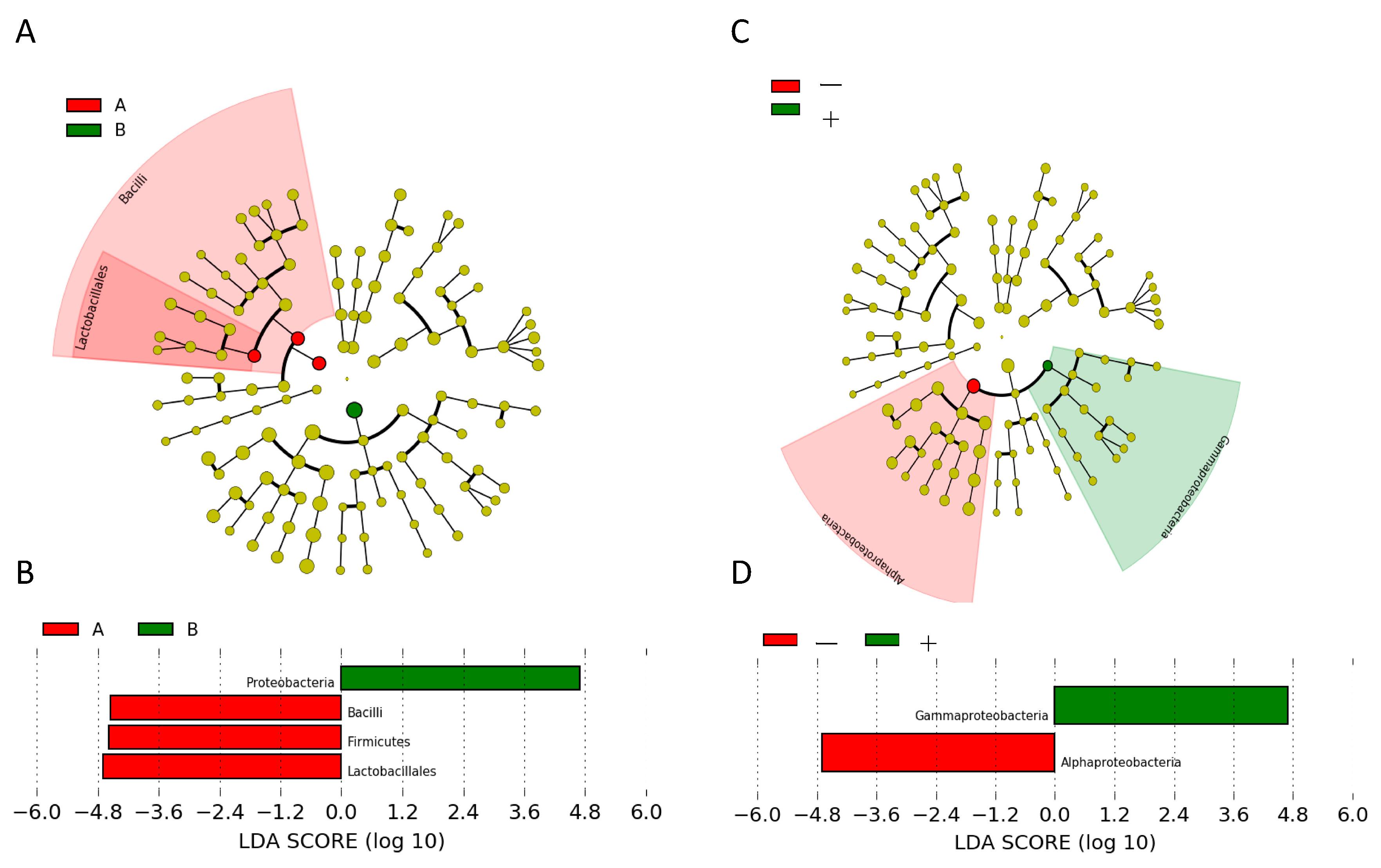
2.5. Analysis of Differentially Abundant Taxa
3. Discussion
4. Methods
4.1. Patients and Sample Preparation
4.2. 16S rRNA Amplification and Targeted Sequencing
4.3. Data Analysis
4.4. Targeted Deep Sequencing of GTF2I Mutation
4.5. Immunohistochemistry for PD-L1
4.6. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Power, S.E.; O’Toole, P.W.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F. Intestinal microbiota, diet and health. Br. J. Nutr. 2014, 111, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, O.V.; Romashin, D.; Zborovskaya, I.B.; Davydov, M.M.; Shogenov, M.S.; Gratchev, A. Human lung microbiome on the way to cancer. J. Immunol. Res. 2019, 2019, 1394191. [Google Scholar] [PubMed]
- Mao, Q.; Jiang, F.; Yin, R.; Wang, J.; Xia, W.; Dong, G.; Ma, W.; Yang, Y.; Xu, L.; Hu, J. Interplay between the lung microbiome and lung cancer. Cancer Lett. 2018, 415, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Goto, T. Airway microbiota as a modulator of lung cancer. Int. J. Mol. Sci. 2020, 21, 3044. [Google Scholar] [CrossRef] [PubMed]
- Laborda-Illanes, A.; Sanchez-Alcoholado, L.; Dominguez-Recio, M.E.; Jimenez-Rodriguez, B.; Lavado, R.; Comino-Méndez, I.; Alba, E.; Queipo-Ortuño, M.I. Breast and gut microbiota action mechanisms in breast cancer pathogenesis and treatment. Cancers 2020, 12, 2465. [Google Scholar] [CrossRef]
- Peters, B.A.; Hayes, R.B.; Goparaju, C.; Reid, C.; Pass, H.I.; Ahn, J. The microbiome in lung cancer tissue and recurrence-free survival. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; San Lucas, A.; et al. Tumor microbiome diversity and composition influence pancreatic cancer outcomes. Cell 2019, 178, 795–806.e12. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.Y.; Shi, S.; Liang, C.; Meng, Q.C.; Hua, J.; Zhang, Y.Y.; Liu, J.; Zhang, B.; Xu, J.; Yu, X.J. The microbiota and microbiome in pancreatic cancer: More influential than expected. Mol. Cancer 2019, 18, 97. [Google Scholar] [CrossRef] [PubMed]
- Engels, E.A. Epidemiology of thymoma and associated malignancies. J. Thorac. Oncol. 2010, 5 (Suppl. S4), S260–S265. [Google Scholar] [CrossRef] [Green Version]
- Venuta, F.; Anile, M.; Diso, D.; Vitolo, D.; Rendina, E.A.; De Giacomo, T.; Francioni, F.; Coloni, G.F. Thymoma and thymic carcinoma. Eur. J. Cardiothorac. Surg. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, A.; Chan, J.K.; Coindre, J.M.; Detterbeck, F.; Girard, N.; Harris, N.L.; Jaffe, E.S.; Kurrer, M.O.; Marom, E.M.; Moreira, A.L.; et al. The 2015 world health organization classification of tumors of the thymus: Continuity and changes. J. Thorac. Oncol. 2015, 10, 1383–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, A.; Strobel, P.; Badve, S.S.; Chalabreysse, L.; Chan, J.K.; Chen, G.; de Leval, L.; Detterbeck, F.; Girard, N.; Huang, J.; et al. ITMIG consensus statement on the use of the WHO histological classification of thymoma and thymic carcinoma: Refined definitions, histological criteria, and reporting. J. Thorac. Oncol. 2014, 9, 596–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.W.; Lee, K.S.; Shin, M.H.; Kim, S.; Woo, S.Y.; Lee, G.; Han, J.; Shim, Y.M.; Choi, Y.S. Thymic epithelial tumors: Prognostic determinants among clinical, histopathologic, and computed tomography findings. Ann. Thorac. Surg. 2015, 99, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.R.; Wright, C.D.; Loehrer, P.J. Thymoma: State of the art. J. Clin. Oncol. 1999, 17, 2280–2289. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Pala, L.; Giaccone, G.; De Pas, T. Thymic epithelial tumors: From biology to treatment. Cancer Treat Rev. 2020, 86, 102014. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R. Myasthenia gravis and thymic neoplasms: A brief review. World J. Clin. Cases 2015, 3, 980–983. [Google Scholar] [CrossRef] [PubMed]
- Romi, F. Thymoma in myasthenia gravis: From diagnosis to treatment. Autoimmune Dis. 2011, 2011, 474512. [Google Scholar] [CrossRef] [Green Version]
- Girard, N.; Lal, R.; Wakelee, H.; Riely, G.J.; Loehrer, P.J. Chemotherapy definitions and policies for thymic malignancies. J. Thorac. Oncol. 2011, 6 (Suppl. S3), S1749–S1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litvak, A.M.; Woo, K.; Hayes, S.; Huang, J.; Rimner, A.; Sima, C.S.; Moreira, A.L.; Tsukazan, M.; Riely, G.J. Clinical characteristics and outcomes for patients with thymic carcinoma: Evaluation of Masaoka staging. J. Thorac. Oncol. 2014, 9, 1810–1815. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, J.; Loehrer, P.J., Sr. The role of chemotherapy in advanced thymoma. J. Thorac. Oncol. 2010, 5 (Suppl. S4), S357–S360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Shi, J.; Fan, L.; Hu, D.; Yang, J.; Zhao, H. Surgical treatment of thymoma: An 11-year experience with 761 patients. Eur. J. Cardiothorac. Surg. 2016, 49, 1144–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gethings-Behncke, C.; Coleman, H.G.; Jordao, H.W.T.; Longley, D.B.; Crawford, N.; Murray, L.J.; Kunzmann, A.T. Fusobacterium nucleatum in the colorectum and its association with cancer risk and survival: A systematic review and meta-analysis. Cancer Epidemiol. Biomarkers Prev. 2020, 29, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S.; et al. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [Green Version]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/beta-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hujoel, P.P.; Drangsholt, M.; Spiekerman, C.; Weiss, N.S. An exploration of the periodontitis-cancer association. Ann. Epidemiol. 2003, 13, 312–316. [Google Scholar] [CrossRef]
- Yang, J.; Mu, X.; Wang, Y.; Zhu, D.; Zhang, J.; Liang, C.; Chen, B.; Wang, J.; Zhao, C.; Zuo, Z.; et al. Dysbiosis of the salivary microbiome is associated with non-smoking female lung cancer and correlated with immunocytochemistry markers. Front. Oncol. 2018, 8, 520. [Google Scholar] [CrossRef] [PubMed]
- Maddi, A.; Sabharwal, A.; Violante, T.; Manuballa, S.; Genco, R.; Patnaik, S.; Yendamuri, S. The microbiome and lung cancer. J. Thorac. Dis. 2019, 11, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Labrada, A.G.; Isla, D.; Artal, A.; Arias, M.; Rezusta, A.; Pardo, J.; Galvez, E.M. The influence of lung microbiota on lung carcinogenesis, immunity, and immunotherapy. Trends Cancer 2020, 6, 86–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Wang, L.; Li, C.; Ding, C.; Li, C.; Fan, W.; Cheng, C.; Gu, B. Microbiota dysbiosis in lung cancer: Evidence of association and potential mechanisms. Transl. Lung Cancer Res. 2020, 9, 1554–1568. [Google Scholar] [CrossRef]
- Stolz, A. Molecular characteristics of xenobiotic-degrading sphingomonads. Appl. Microbiol. Biotechnol. 2009, 81, 793–811. [Google Scholar] [CrossRef] [PubMed]
- Aso, Y.; Miyamoto, Y.; Harada, K.M.; Momma, K.; Kawai, S.; Hashimoto, W.; Mikami, B.; Murata, K. Engineered membrane superchannel improves bioremediation potential of dioxin-degrading bacteria. Nat. Biotechnol. 2006, 24, 188–189. [Google Scholar] [CrossRef]
- Miller, T.R.; Delcher, A.L.; Salzberg, S.L.; Saunders, E.; Detter, J.C.; Halden, R.U. Genome sequence of the dioxin-mineralizing bacterium Sphingomonas wittichii RW1. J. Bacteriol. 2010, 192, 6101–6102. [Google Scholar] [CrossRef] [Green Version]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Dong, Z.; Chen, B.; Pan, H.; Wang, D.; Liu, M.; Yang, Y.; Zou, M.; Yang, J.; Xiao, K.; Zhao, R.; et al. Detection of microbial 16S rRNA gene in the serum of patients with gastric cancer. Front. Oncol. 2019, 9, 608. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.F.; Chen, Y.J.; Fan, T.C.; Chang, N.C.; Chen, Y.J.; Midha, M.K.; Chen, T.H.; Yang, H.H.; Wang, Y.T.; Yu, A.L.; et al. Analysis of microbial sequences in plasma cell-free DNA for early-onset breast cancer patients and healthy females. BMC Med. Genom. 2018, 11 (Suppl. S1), 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yu, Y.; Choi, L.; Song, Y.; Wu, M.; Wang, G.; Li, M. Phenylobacterium soli sp. nov., isolated from arsenic and cadmium contaminated farmland soil. Int. J. Syst. Evol. Microbiol. 2019, 69, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wen, X.; Zhao, L.; Shi, Y.; Jin, H. Crude oil treatment leads to shift of bacterial communities in soils from the deep active layer and upper permafrost along the China-Russia Crude Oil Pipeline route. PLoS ONE 2014, 9, e96552. [Google Scholar] [CrossRef]
- Huang, Y.; Pan, H.; Wang, Q.; Ge, Y.; Liu, W.; Christie, P. Enrichment of the soil microbial community in the bioremediation of a petroleum-contaminated soil amended with rice straw or sawdust. Chemosphere 2019, 224, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Warmbrunn, M.V.; Nieuwdorp, M.; Clément, K. Metabolism and metabolic disorders and the microbiome: The intestinal microbiota associated with obesity, lipid metabolism and metabolic health: Pathophysiology and therapeutic strategies. Gastroenterology 2021, 160, 573–599. [Google Scholar] [CrossRef] [PubMed]
- Sommariva, M.; Le Noci, V.; Bianchi, F.; Camelliti, S.; Balsari, A.; Tagliabue, E.; Sfondrini, L. The lung microbiota: Role in maintaining pulmonary immune homeostasis and its implications in cancer development and therapy. Cell. Mol. Life Sci. 2020, 77, 2739–2749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, J.M.; Malfertheiner, P.; Lee, Y.C.; Sheu, B.S.; Sugano, K.; Cheng, H.C.; Yeoh, K.G.; Hsu, P.I.; Goh, K.L.; Mahachai, V.; et al. Screening and eradication of Helicobacter pylori for gastric cancer prevention: The Taipei global consensus. Gut 2020, 69, 2093–2112. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Gail, M.H.; Consonni, D.; Carugno, M.; Humphrys, M.; Pesatori, A.C.; Caporaso, N.E.; Goedert, J.J.; Ravel, J.; Landi, M.T. Characterizing human lung tissue microbiota and its relationship to epidemiological and clinical features. Genome Biol. 2016, 17, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, R.; Goto, T.; Hirotsu, Y.; Yokoyama, Y.; Nakagomi, T.; Otake, S.; Amemiya, K.; Oyama, T.; Mochizuki, H.; Omata, M. Primary driver mutations in GTF2I specific to the development of thymomas. Cancers 2020, 12, 2032. [Google Scholar] [CrossRef] [PubMed]
- Loessner, H.; Endmann, A.; Leschner, S.; Westphal, K.; Rohde, M.; Miloud, T.; Hämmerling, G.; Neuhaus, K.; Weiss, S. Remote control of tumour-targeted Salmonella enterica serovar Typhimurium by the use of L-arabinose as inducer of bacterial gene expression in vivo. Cell. Microbiol. 2007, 9, 1529–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, A.R.; Thunnissen, F.B. Histological typing of lung and pleural tumours: Third edition. J. Clin. Pathol. 2001, 54, 498–499. [Google Scholar] [CrossRef] [Green Version]
- Travis, W.D.; Brambilla, E.; Nicholson, A.G.; Yatabe, Y.; Austin, J.H.M.; Beasley, M.B.; Chirieac, L.R.; Dacic, S.; Duhig, E.; Flieder, D.B.; et al. The 2015 World Health Organization classification of lung tumors: Impact of genetic, clinical and radiologic advances since the 2004 classification. J. Thorac. Oncol. 2015, 10, 1243–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chansky, K.; Detterbeck, F.C.; Nicholson, A.G.; Rusch, V.W.; Vallieres, E.; Groome, P.; Kennedy, C.; Krasnik, M.; Peake, M.; Shemanski, L.; et al. The IASLC lung cancer staging project: External validation of the revision of the TNM Stage groupings in the eighth edition of the TNM classification of lung cancer. J. Thorac. Oncol. 2017, 12, 1109–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffini, E.; Fang, W.; Guerrera, F.; Huang, J.; Okumura, M.; Kim, D.K.; Girard, N.; Bille, A.; Boubia, S.; Cangir, A.K.; et al. The international association for the study of lung cancer thymic tumors staging project: The impact of the Eighth edition of the Union for international cancer control and american joint committee on cancer TNM stage classification of thymic tumors. J. Thorac. Oncol. 2020, 15, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, K.; Hirotsu, Y.; Goto, T.; Nakagomi, H.; Mochizuki, H.; Oyama, T.; Omata, M. Touch imprint cytology with massively parallel sequencing (TIC-seq): A simple and rapid method to snapshot genetic alterations in tumors. Cancer Med. 2016, 5, 3426–3436. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirotsu, Y.; Amemiya, K.; Nakagomi, T.; Shikata, D.; Yokoyama, Y.; Okimoto, K.; Oyama, T.; Mochizuki, H.; Omata, M. Distribution of circulating tumor DNA in lung cancer: Analysis of the primary lung and bone marrow along with the pulmonary venous and peripheral blood. Oncotarget 2017, 8, 59268–59281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, R.; Nakagomi, T.; Goto, T.; Hirotsu, Y.; Shikata, D.; Yokoyama, Y.; Otake, S.; Amemiya, K.; Oyama, T.; Mochizuki, H.; et al. Identification of clonality through genomic profile analysis in multiple lung cancers. J. Clin. Med. 2020, 9, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagomi, T.; Goto, T.; Hirotsu, Y.; Shikata, D.; Yokoyama, Y.; Higuchi, R.; Otake, S.; Amemiya, K.; Oyama, T.; Mochizuki, H.; et al. Genomic characteristics of invasive mucinous adenocarcinomas of the lung and potential therapeutic targets of B7-H3. Cancers 2018, 10, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagomi, T.; Hirotsu, Y.; Goto, T.; Shikata, D.; Yokoyama, Y.; Higuchi, R.; Otake, S.; Amemiya, K.; Oyama, T.; Mochizuki, H.; et al. Clinical implications of noncoding indels in the surfactant-encoding genes in lung cancer. Cancers 2019, 11, 552. [Google Scholar] [CrossRef] [Green Version]
- Oyama, T.; Goto, T.; Amemiya, K.; Hirotsu, Y.; Omata, M. Squamous cell carcinoma of the lung with micropapillary pattern. J. Thorac. Oncol. 2020, 15, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Hirotsu, Y.; Oyama, T.; Amemiya, K.; Omata, M. Analysis of tumor-derived DNA in plasma and bone marrow fluid in lung cancer patients. Med. Oncol. 2016, 33, 29. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wang, Y.; Qian, P.Y. Sensitivity and correlation of hypervariable regions in 16S rRNA genes in phylogenetic analysis. BMC Bioinform. 2016, 17, 135. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Number of Patients | Overall Percentage | |
|---|---|---|---|
| Total number | 19 | ||
| Age, median (range) | 68 (42–81) | ||
| Sex | |||
| Male | 11 | 57.9% | |
| Female | 8 | 42.1% | |
| Histology | |||
| Type A | 5 | 26.3% | |
| Type AB | 3 | 15.8% | |
| Type B1 | 5 | 26.3% | |
| Type B2 | 4 | 21.1% | |
| Type B3 | 2 | 10.5% | |
| Tumor size (cm) | |||
| ≤3 | 7 | 36.8% | |
| 3 < size ≤ 5 | 8 | 42.1% | |
| >5 | 4 | 21.1% | |
| Masaoka Stage | |||
| I | 7 | 36.8% | |
| II | 9 | 47.4% | |
| III | 2 | 10.5% | |
| IV | 1 | 5.3% | |
| Smoking Status (Pack year) | |||
| 0 | 7 | 36.8% | |
| 0 < PY ≤ 30 | 8 | 42.1% | |
| >30 | 4 | 21.1% | |
| Myasthenia gravis | |||
| present | 1 | 5.3% | |
| absent | 18 | 94.7% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Higuchi, R.; Goto, T.; Hirotsu, Y.; Otake, S.; Oyama, T.; Amemiya, K.; Ohyama, H.; Mochizuki, H.; Omata, M. Sphingomonas and Phenylobacterium as Major Microbiota in Thymic Epithelial Tumors. J. Pers. Med. 2021, 11, 1092. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm11111092
Higuchi R, Goto T, Hirotsu Y, Otake S, Oyama T, Amemiya K, Ohyama H, Mochizuki H, Omata M. Sphingomonas and Phenylobacterium as Major Microbiota in Thymic Epithelial Tumors. Journal of Personalized Medicine. 2021; 11(11):1092. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm11111092
Chicago/Turabian StyleHiguchi, Rumi, Taichiro Goto, Yosuke Hirotsu, Sotaro Otake, Toshio Oyama, Kenji Amemiya, Hiroshi Ohyama, Hitoshi Mochizuki, and Masao Omata. 2021. "Sphingomonas and Phenylobacterium as Major Microbiota in Thymic Epithelial Tumors" Journal of Personalized Medicine 11, no. 11: 1092. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm11111092