Antimicrobial Activity of an Extract of Hermetia illucens Larvae Immunized with Lactobacillus casei against Salmonella Species


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. H. illucens Larvae and Bacteria Strains
2.2. Immunization of HIL
2.3. Preparation of HIL Extracts
2.4. Evaluation of the Antimicrobial Activities in HIL Hemolymph and Extract
2.5. Determination of Minimum Inhibitory Concentrations (MICs) of HIL Extracts
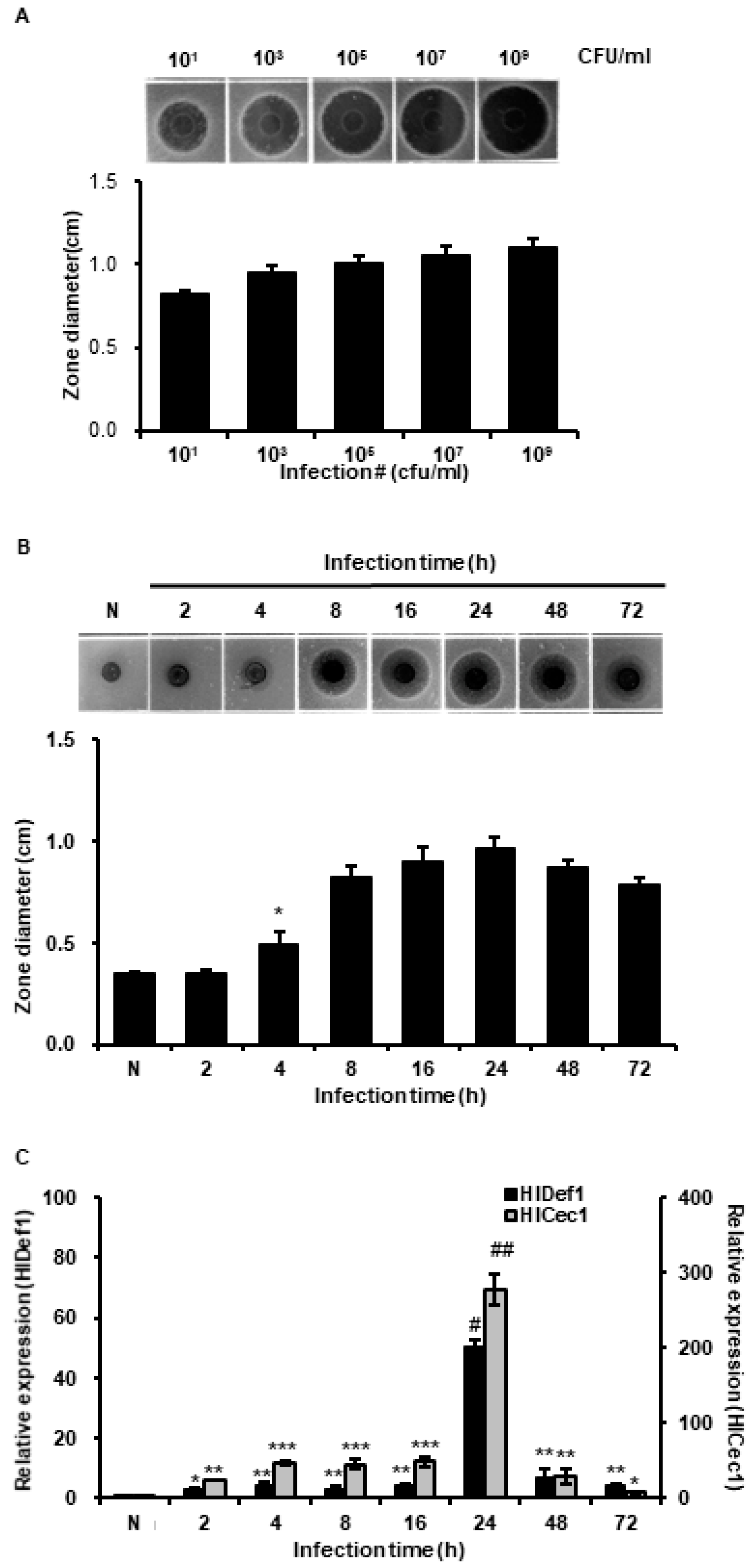
2.6. Analysis of AMPs Transcription
2.7. Cytotoxicity Analysis by MTT Assay
2.8. Statistical Analysis
3. Results
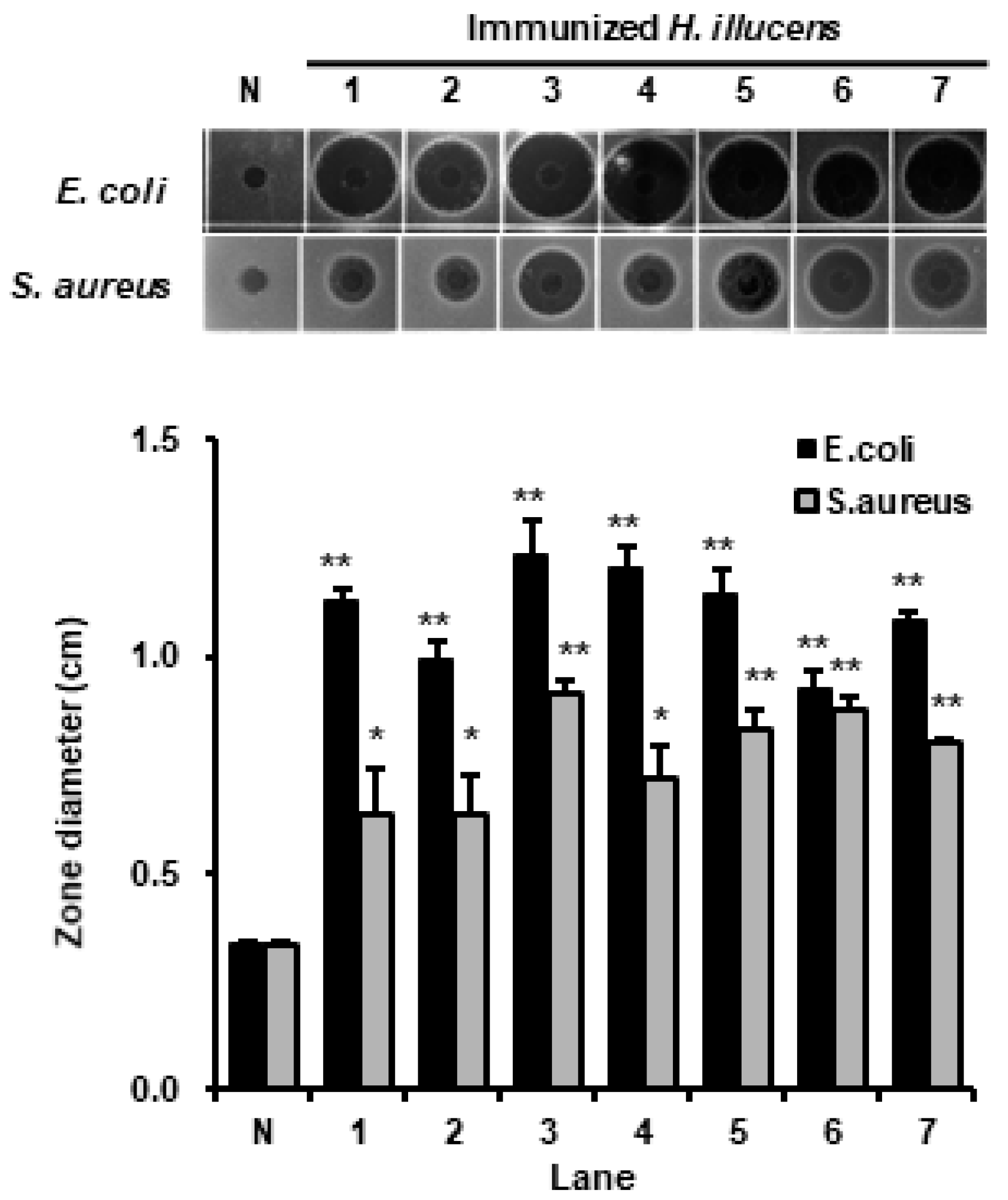
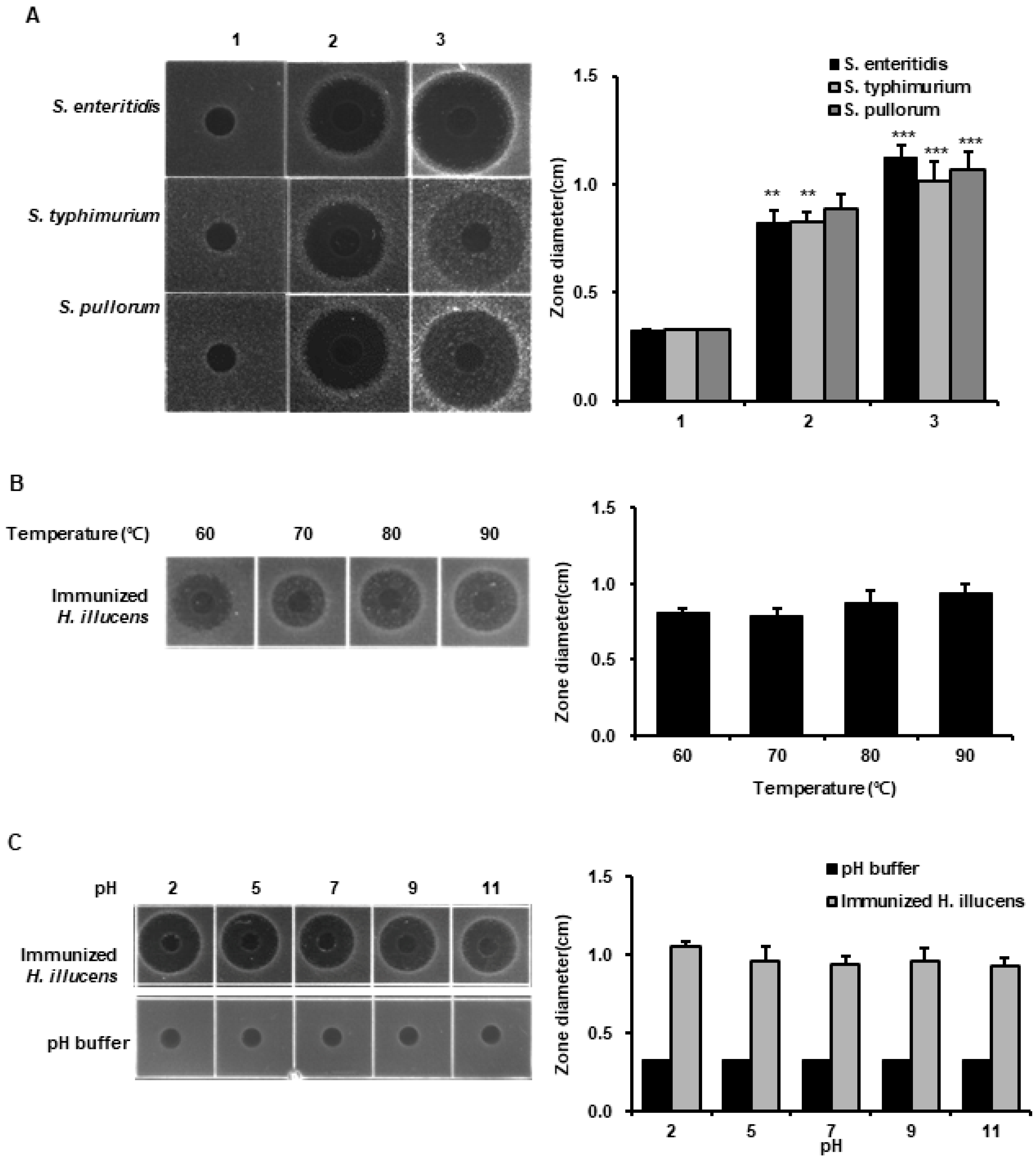
3.1. Optimization of Lactobacillus Species for the Mass Production of AMPs in H. illucens
3.2. Determination of Optimal Conditions for the Mass Production of AMPs
3.3. Analysis of the Antimicrobial Activities of HIL Hemolymph against Salmonella Species
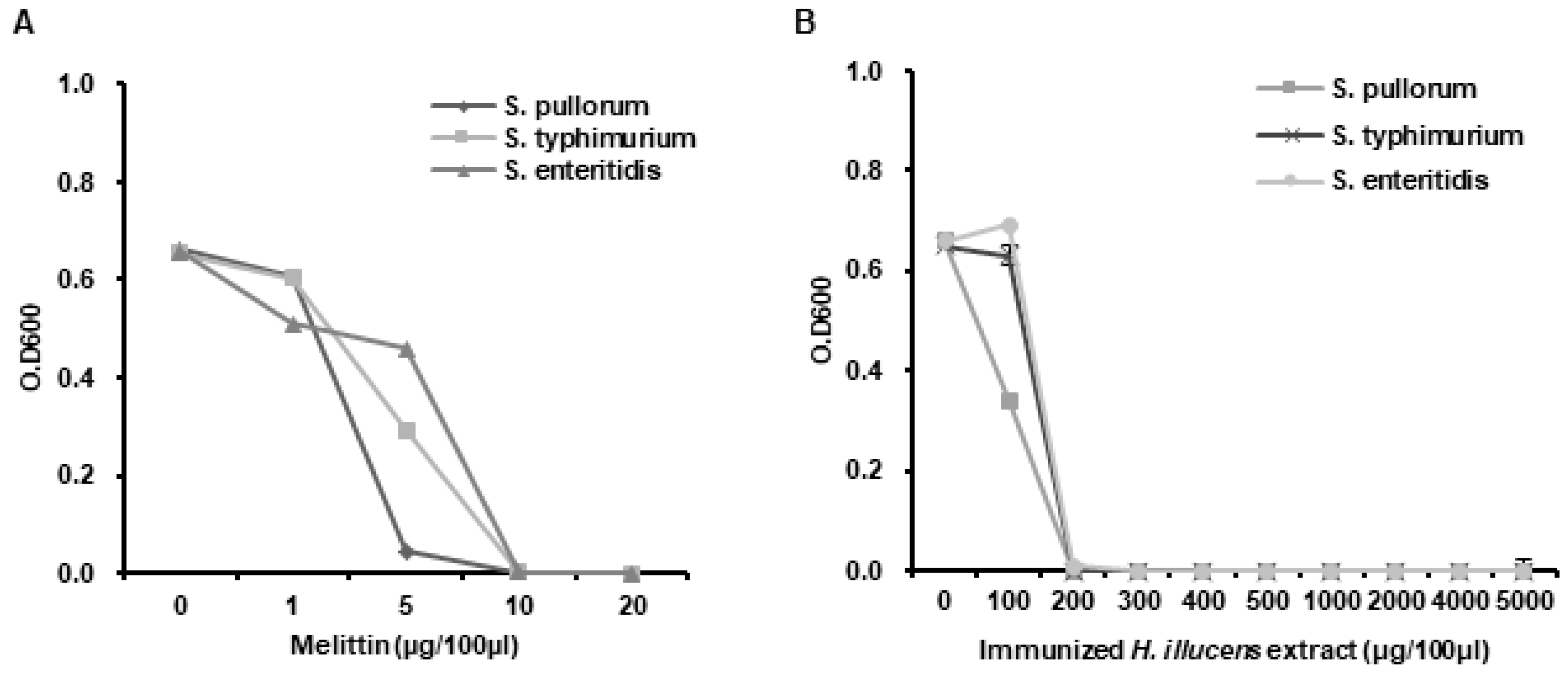
3.4. Determination of the MICs of L. casei-Immunized HIL Extract
3.5. Analysis of the Antimicrobial Activities of L. casei-Immunized HIL Extract against Salmonella Species
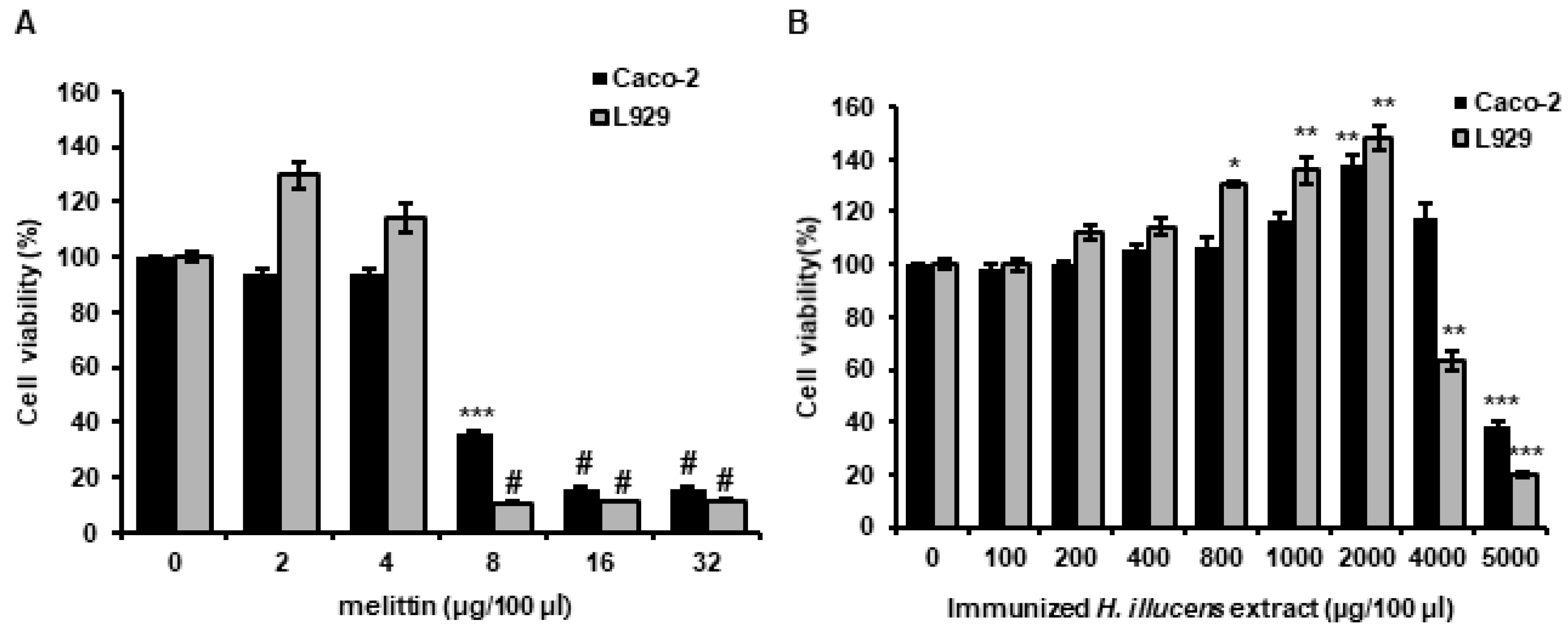
3.6. Analysis of the Cytotoxicities of L. casei-Immunized HIL Extracts to Mammalian Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltzer, S.A.; Brown, M.H. Antimicrobial peptides–promising alternatives to conventional antibiotics. J. Mol. Microbiol. Biotechnol. 2011, 20, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.D. Antibiotic use in animal feed and its impact on human healt. Nutr. Res. Rev. 2000, 13, 279–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.; Seo, O.; Choi, H.; Chae, H.; Na, J.; Bang, H.; Kim, D.; Park, S.; Kim, M.; Jung, S. Effect of dietary supplementation of chlorella powder on production performances, blood components. In Proceedings of the Korean Journal of Poultty Science 27th Regular Conference, Daejeon, Korea, 12 November 2010; pp. 106–108. [Google Scholar]
- Yoo, J.; Park, G.H.; Sung, J.S.; Song, H.; Shin, S.Y.; Jung, W.H.; Heo, J.M. Feed additives in broiler diets to produce healthy chickens without in-feed antimicrobial compounds. Korean J. Agric. Sci. 2014, 41, 441–453. [Google Scholar] [CrossRef]
- Kalsy, M.; Tonk, M.; Hardt, M.; Dobrindt, U.; Zdybicka-Barabas, A.; Cytrynska, M.; Vilcinskas, A.; Mukherjee, K. The insect antimicrobial peptide cecropin A disrupts uropathogenic Escherichia coli biofilms. NPJ Biofilm. Microbiomes 2020, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Patočka, J.; Kuča, K. Insect antimicrobial peptides, a mini review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Boman, H.G. Peptide antibiotics and their role in innate immunity. Annu. Rev. Immunol. 1995, 13, 61–92. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Hoffmann, J.A.; Kafatos, F.C.; Janeway, C.A.; Ezekowitz, R. Phylogenetic perspectives in innate immunity. Science 1999, 284, 1313–1318. [Google Scholar] [CrossRef] [Green Version]
- Casteels, P.; Ampe, C.; Rivière, L.; Van Damme, J.; Elicone, C.; Fleming, M.; JACOBS, F.; Tempst, P. Isolation and characterization of abaecin, a major antibacterial response peptide in the honeybee (Apis mellifera). Eur. J. Biochem. 1990, 187, 381–386. [Google Scholar] [CrossRef]
- Goo, T.-W.; Yun, E.-Y.; Kim, S.-W.; Choi, K.-H.; Kang, S.-W.; Kwon, K.-S.; Yu, K.; Kwon, O.-Y. Bombyx mori protein disulfide isomerase enhances the production of nuecin, an antibacterial protein. BMB Rep. 2008, 41, 400–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goo, T.W.; Yun, E.Y.; Kim, S.W.; Choi, K.H.; Kang, S.W.; Kwon, K.; Choi, J.-S.; Kwon, O.-Y. Secretion of the antibacterial recombinant protein enbocin. Z. Nat. C 2008, 63, 284–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-R.; Lee, E.-M.; Yoon, H.-J.; Choi, Y.-S.; Yun, E.-Y.; Hwang, J.-S.; Jin, B.-R.; Lee, I.-H.; Kim, I.-S. Antibacterial Activity of Peptides Synthesized Based on the Bombus ignites abaecin, A Novel Proline-Rich Antimicrobial Peptide. Int. J. Ind. Entomol. 2007, 14, 147–150. [Google Scholar]
- Otvos, L., Jr. Antibacterial peptides isolated from insects. J. Pept. Sci. Off. Publ. Eur. Pept. Soc. 2000, 6, 497–511. [Google Scholar]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jang, A.Y.; Lin, S.; Lim, S.; Kim, D.; Park, K.; Han, S.M.; Yeo, J.H.; Seo, H.S. Melittin, a honeybee venom-derived antimicrobial peptide, may target methicillin-resistant Staphylococcus aureus. Mol. Med. Rep. 2015, 12, 6483–6490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosler, S.; Gerceker, A.A. In vitro activities of antimicrobial cationic peptides; melittin and nisin, alone or in combination with antibiotics against Gram-positive bacteria. J. Chemother. 2012, 24, 137–143. [Google Scholar] [CrossRef]
- Pashaei, F.; Bevalian, P.; Akbari, R.; Bagheri, K.P. Single dose eradication of extensively drug resistant Acinetobacter spp. in a mouse model of burn infection by melittin antimicrobial peptide. Microb. Pathog. 2019, 127, 60–69. [Google Scholar] [CrossRef]
- Choi, Y.-C.; Park, K.-H.; Nam, S.-H.; Jang, B.-G.; Kim, J.-H.; Kim, D.-W.; Yu, D.-J. The effect on growth performance of chicken meat in broiler chicks by dietary supplementation of black soldier fly larvae, Hermetia illucens (Diptera: Stratmyidae). J. Sericultural Entomol. Sci. 2013, 51, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-I.; Kim, J.-W.; Yoe, S.M. Purification and characterization of a novel antibacterial peptide from black soldier fly (Hermetia illucens) larvae. Dev. Comp. Immunol. 2015, 52, 98–106. [Google Scholar] [CrossRef]
- Choi, W.H.; Choi, H.J.; Goo, T.W.; Quan, F.S. Novel antibacterial peptides induced by probiotics in Hermetia illucens (Diptera: Stratiomyidae) larvae. Entomol. Res. 2018, 48, 237–247. [Google Scholar] [CrossRef]
- Park, K.; Yun, E.-Y.; Park, S.-W.; Goo, T.-W. The antimicrobial activity of bacterial-challenged black soldier fly, Hermetia illucens. J. Life Sci. 2016, 26, 1409–1414. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Lee, Y.J.; Kang, M.S.; Kwag, S.I.; Cho, J.K. Dissemination and tracking of Salmonella spp. in integrated broiler operation. J. Vet. Sci. 2007, 8, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Olsen, S.J.; MacKinon, L.C.; Goulding, J.S.; Bean, N.H.; Slutsker, L. Surveillance for Foodborne-Disease Outbreaks, United States, 1993–1997; U.S. Department of Health & Human Servieces: Atlanta, GA, USA, 2000.
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Vetterli, S.U.; Zerbe, K.; Müller, M.; Urfer, M.; Mondal, M.; Wang, S.-Y.; Moehle, K.; Zerbe, O.; Vitale, A.; Pessi, G. Thanatin targets the intermembrane protein complex required for lipopolysaccharide transport in Escherichia coli. Sci. Adv. 2018, 4, eaau2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.H.; Kwak, K.W.; Nam, S.H.; Choi, J.Y.; Lee, S.; Kim, H.; Kim, S.H. Antibacterial activity of larval extract from the black soldier fly hermetia illucens (diptera: Stratiomyidae) against plant pathogens. J. Entomol. Zool. Stud. 2015, 3, 176–179. [Google Scholar]
- Rossoni, R.D.; Fuchs, B.B.; De Barros, P.P.; Velloso, M.d.S.; Jorge, A.O.C.; Junqueira, J.C.; Mylonakis, E. Lactobacillus paracasei modulates the immune system of Galleria mellonella and protects against Candida albicans infection. PLoS ONE 2017, 12, e0173332. [Google Scholar] [CrossRef]
- Daisley, B.A.; Trinder, M.; McDowell, T.W.; Welle, H.; Dube, J.S.; Ali, S.N.; Leong, H.S.; Sumarah, M.W.; Reid, G. Neonicotinoid-induced pathogen susceptibility is mitigated by Lactobacillus plantarum immune stimulation in a Drosophila melanogaster model. Sci. Rep. 2017, 7, 2703. [Google Scholar] [CrossRef]
- Nishida, S.; Ono, Y.; Sekimizu, K. Lactic acid bacteria activating innate immunity improve survival in bacterial infection model of silkworm. Drug Discov. Ther. 2016, 10, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Jorjão, A.L.; de Oliveira, F.E.; Leão, M.V.P.; Jorge, A.O.C.; de Oliveira, L.D. Effect of Lactobacillus rhamnosus on the response of Galleria mellonella against Staphylococcus aureus and Escherichia coli infections. Arch. Microbiol. 2018, 200, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Fiore, E.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Gram-Posit. Pathog 2019, 378–397. [Google Scholar]
- Khanna, A.; KhAnnA, M.; AggArWAl, A. Serratia marcescens-a rare opportunistic nosocomial pathogen and measures to limit its spread in hospitalized patients. J. Clin. Diagn. Res. JCDR 2013, 7, 243. [Google Scholar] [PubMed]
- Perumal Samy, R.; Gopalakrishnakone, P.; Thwin, M.; Chow, T.; Bow, H.; Yap, E.; Thong, T. Antibacterial activity of snake, scorpion and bee venoms: A comparison with purified venom phospholipase A2 enzymes. J. Appl. Microbiol. 2007, 102, 650–659. [Google Scholar] [CrossRef]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.; Cunha, W.R.; Almeida, R.d.; Martins, C.H. Antimicrobial activity of apitoxin, melittin and phospholipase A2 of honey bee (Apis mellifera) venom against oral pathogens. An. Acad. Bras. Ciências 2015, 87, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.J.; Kang, S.J.; Kim, B.M.; Kim, Y.J.; Woo, H.D.; Chung, H.W. Cytotoxicity of honeybee (Apis mellifera) venom in normal human lymphocytes and HL-60 cells. Chem. Biol. Interact. 2007, 169, 189–197. [Google Scholar] [CrossRef]
- Xiu-Qin, L.; Chao, J.; Wei, Y.; Yun, L.; Min-Li, Y.; Xiao-Gang, C. UPLC-PDAD analysis for simultaneous determination of ten synthetic preservatives in foodstuff. Chromatographia 2008, 68, 57–63. [Google Scholar] [CrossRef]
- Anand, S.; Sati, N. Artificial preservatives and their harmful effects: Looking toward nature for safer alternatives. Int. J. Pharm. Sci. Res. 2013, 4, 2496. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequences | |
|---|---|---|
| HiCec1 | Forward | 5′-TTGGTCAACGAGTTCGTGATGC-3′ |
| Reverse | 5′-TCCTTGTTGTGGTGGTCCACCT-3′ | |
| HiDef1 | Forward | 5′-AGGTGGTGGAGCAGCATTAC-3′ |
| Reverse | 5′-ACGACGTCCCAAAGCAATAC-3′ | |
| Act5C | Forward | 5′-AAGGACTCGTACGTGGGTG-3′ |
| Reverse | 5′-CATCTTCTCACGGTTGGC-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.-S.; Yun, E.-Y.; Goo, T.-W. Antimicrobial Activity of an Extract of Hermetia illucens Larvae Immunized with Lactobacillus casei against Salmonella Species. Insects 2020, 11, 704. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100704
Lee K-S, Yun E-Y, Goo T-W. Antimicrobial Activity of an Extract of Hermetia illucens Larvae Immunized with Lactobacillus casei against Salmonella Species. Insects. 2020; 11(10):704. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100704
Chicago/Turabian StyleLee, Kyu-Shik, Eun-Young Yun, and Tae-Won Goo. 2020. "Antimicrobial Activity of an Extract of Hermetia illucens Larvae Immunized with Lactobacillus casei against Salmonella Species" Insects 11, no. 10: 704. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11100704