Exploring Simplified Methods for Insect Chitin Extraction and Application as a Potential Alternative Bioethanol Resource

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cockroach Chitin Extraction and Characterization
2.1.1. Chitin Extraction
2.1.2. Characterization of the Extracted Cockroach Chitin
Fourier Transform Infrared Spectroscopy (FT-IR)
X-ray Diffraction (XRD)
Elemental Analysis
Scanning Electron Microscopy (SEM)
2.1.3. The Chitin Content in the Dried Exoskeleton
2.2. Bioethanol Production
2.2.1. Fungal Strains and Culture Media
Fungal Strains and Their Maintenance
Production Medium Preparation
2.2.2. Cockroach Chitin Fermentation Mechanisms
Direct Fermentation Mechanism
Indirect Fermentation Mechanism
Bioethanol Quantification
Statistical Analysis
3. Results
3.1. Chitin Extraction and Characterization
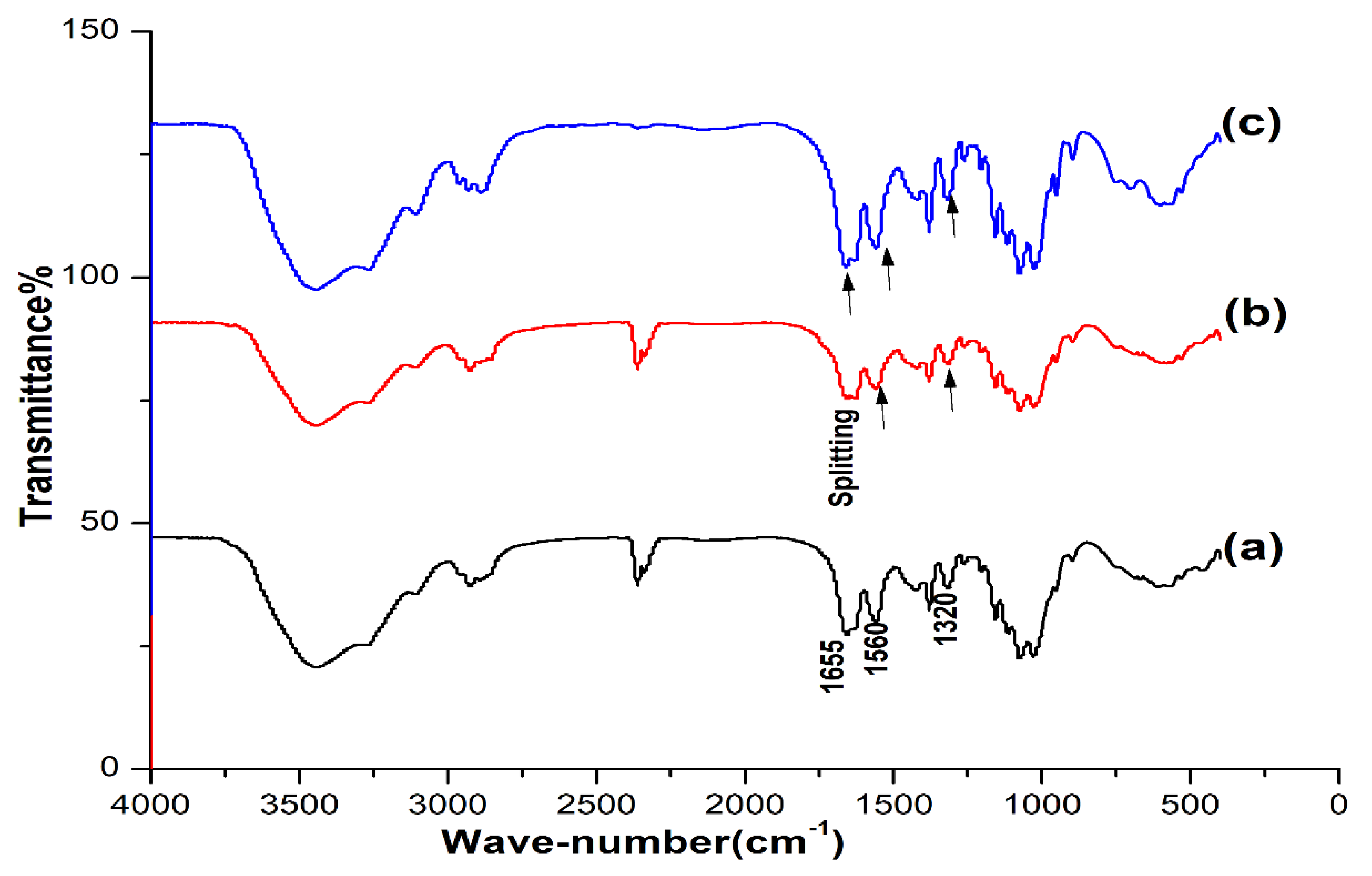
3.1.1. FT-IR
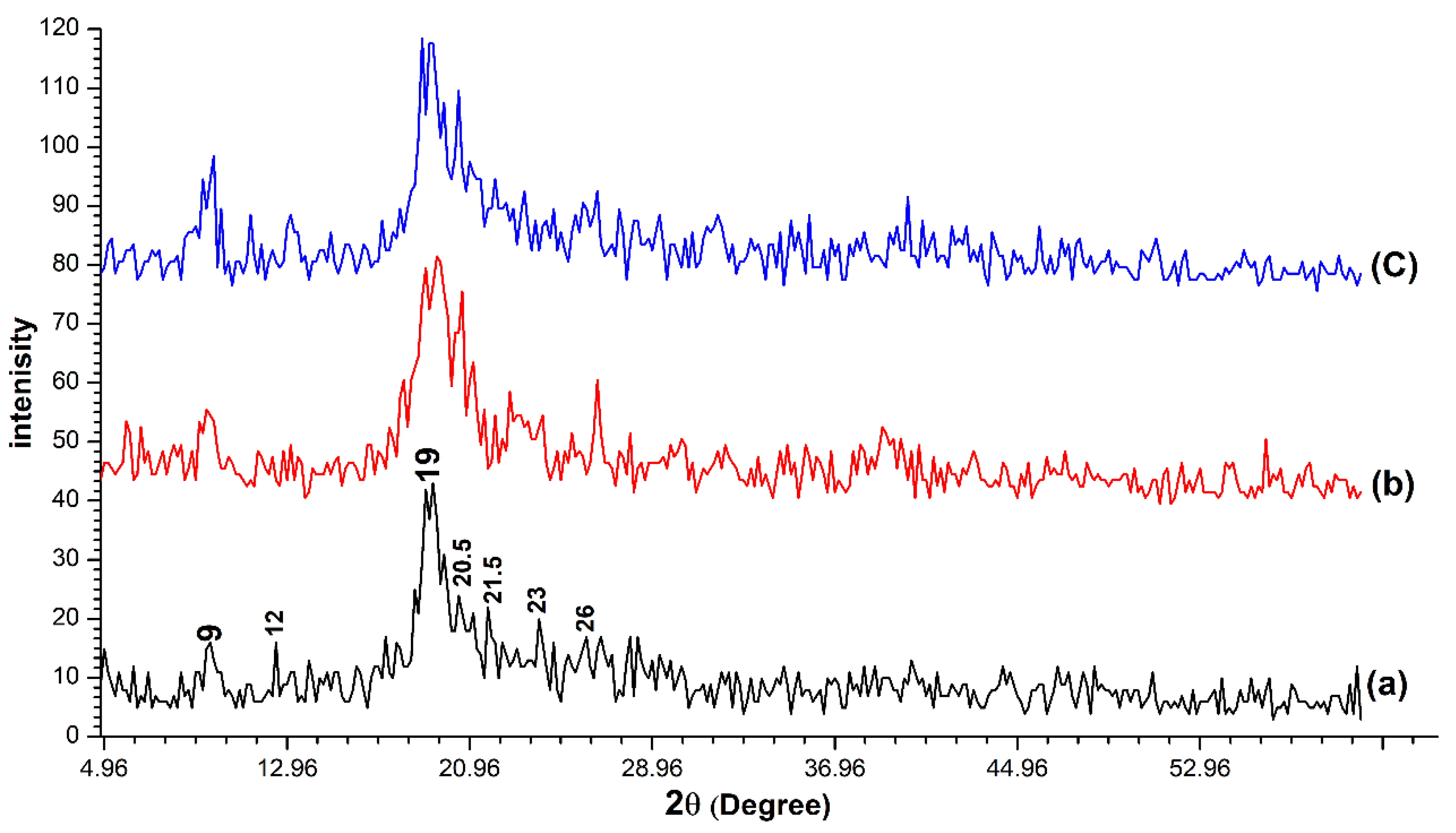
3.1.2. XRD
3.1.3. Elemental Analysis
3.1.4. SEM
Chitin Content in the Dried Cockroach Exoskeleton
3.2. Effect of Initial pH on Chitin Degradation
3.3. Bioethanol Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hill, J.; Nelson, E.; Tilman, D.; Polasky, S.; Tiffany, D. Environmental, economic, and energetic costs and benefits of biodiesel and ethanol biofuels. Proc. Natl. Acad. Sci. USA 2006, 103, 11206–11210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, L.; Owende, P. Biofuels from microalgae-A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Searchinger, T.; Heimlich, R.; Houghton, R.A.; Dong, F.; Elobeid, A.; Fabiosa, J.; Tokgoz, S.; Hayes, D.; Yu, T.-H. Use of U.S. Croplands for Biofuels Increases Greenhouse Gases Through Emissions from Land Use Change. Science 2008, 319, 1238–1240. [Google Scholar] [CrossRef] [PubMed]
- Himmel, M.E.; Ding, S.-Y.; Johnson, D.K.; Adney, W.S.; Nimlos, M.R.; Brady, J.W.; Foust, T.D. Biomass recalcitrance: Engineering plants and enzymes for biofuels production. Science 2007, 315, 804–807. [Google Scholar] [CrossRef] [Green Version]
- Rosegrant, M.W.; Zhu, T.; Msangi, S.; Sulser, T. Global scenarios for biofuels: Impacts and implications. Rev. Agric. Econ. 2008, 30, 495–505. [Google Scholar] [CrossRef]
- RFA. Accelerating Industry Innovation: 2012 Ethanol Industry Outlook; RFA: Washington, DC, USA, 2007. [Google Scholar] [CrossRef]
- Sarkar, N.; Ghosh, S.K.; Bannerjee, S.; Aikat, K. Bioethanol production from agricultural wastes: An overview. Renew. Energy 2012, 37, 19–27. [Google Scholar] [CrossRef]
- Inokuma, K.; Takano, M.; Hoshino, K. Direct ethanol production from N-acetylglucosamine and chitin substrates by Mucor species. Biochem. Eng. J. 2013, 72, 24–32. [Google Scholar] [CrossRef]
- Tsurkan, M.V.; Voronkina, A.; Khrunyk, Y.; Wysokowski, M.; Petrenko, I.; Ehrlich, H. Progress in Chitin Analytics. Carbohydr. Polym. 2020, 252, 117204. [Google Scholar] [CrossRef]
- Evvyernie, D.; Morimoto, K.; Karita, S.; Kimura, T.; Sakka, K.; Ohmiya, K. Conversion of chitinous wastes to hydrogen gas by Clostridium paraputrificum M-21. J. Biosci. Bioeng. 2001, 91, 339–343. [Google Scholar] [CrossRef]
- Morimoto, K.; Kimura, T.; Sakka, K.; Ohmiya, K. Overexpression of a hydrogenase gene in Clostridium paraputrificum to enhance hydrogen gas production. FEMS Microbiol. Lett. 2005, 246, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Evvyernie, D.; Yamazaki, S.; Morimoto, K.; Karita, S.; Kimura, T.; Sakka, K.; Ohmiya, K. Identification and characterization of Clostridium paraputrificum M-21, a chitinolytic, mesophilic and hydrogen-producing bacterium. J. Biosci. Bioeng. 2000, 89, 596–601. [Google Scholar] [CrossRef]
- Inokuma, K.; Hasunuma, T.; Kondo, A. Ethanol production from N-acetyl-d-glucosamine by Scheffersomyces stipitis strains. AMB Express 2016, 6, 83. [Google Scholar] [CrossRef] [Green Version]
- Millati, R.; Edebo, L.; Taherzadeh, M.J. Performance of Rhizopus, Rhizomucor, and Mucor in ethanol production from glucose, xylose, and wood hydrolyzates. Enzym. Microb. Technol. 2005, 36, 294–300. [Google Scholar] [CrossRef]
- Sues, A.; Millati, R.; Edebo, L.; Taherzadeh, M.J. Ethanol production from hexoses, pentoses, and dilute-acid hydrolyzate by Mucor indicus. FEMS Yeast Res. 2005, 5, 669–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, A.M.; Gooday, G.W. Properties of Chitinase Activities from Mucor mucedo: Evidence for a Membrane-bound Zymogenic Form. J. Gen. Microbiol. 1984, 130, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Tsuji, Y.; Matsushita, S.; Kumagai, H.; Tochikura, T. Purification and Properties of beta-N-Acetylhexosaminidase from Mucor fragilis Grown in Bovine Blood. Appl. Environ. Microbiol. 1986, 51, 1019–1023. [Google Scholar] [CrossRef] [Green Version]
- Brunner, E.; Richthammer, P.; Ehrlich, H.; Paasch, S.; Simon, P.; Ueberlein, S.; van Pée, K.H. Chitin-based organic networks: An integral part of cell wall biosilica in the diatom Thalassiosira pseudonana. Angew. Chem. Int. Ed. 2009, 48, 9724–9727. [Google Scholar] [CrossRef]
- Ehrlich, H. Chitin and collagen as universal and alternative templates in biomineralization. Int. Geol. Rev. 2010, 52, 661–699. [Google Scholar] [CrossRef]
- Petrenko, I.; Khrunyk, Y.; Voronkina, A.; Kovalchuk, V.; Fursov, A.; Tsurkan, D.; Ivanenko, V. Poriferan chitin: 3D scaffolds from nano-to macroscale. A review. Lett. Appl. Nanobiosci. 2020, 9, 1004–1014. [Google Scholar]
- Tsurkan, D.; Wysokowski, M.; Petrenko, I.; Voronkina, A.; Khrunyk, Y.; Fursov, A.; Ehrlich, H. Modern scaffolding strategies based on naturally pre-fabricated 3D biomaterials of poriferan origin. Appl. Phys. A 2020, 126, 382. [Google Scholar] [CrossRef]
- Arbia, W.; Arbia, L.; Adour, L.; Abdeltif, A. Chitin Extraction from Crustacean Shells Using Biological Methods—A Review. Food Technol. Biotech. 2013, 51, 12–25. [Google Scholar]
- Dutta, P.K.; Dutta, J.; Tripathi, V.S. Chitin, and chitosan: Chemistry, properties, and applications. J. Sci. Ind. Res. 2004, 63, 20–31. [Google Scholar]
- Liu, S.; Sun, J.; Yu, L.; Zhang, C.; Bi, J.; Zhu, F.; Qu, M.; Jiang, C.; Yang, Q. Extraction and Characterization of Chitin from the Beetle Holotrichia parallela Motschulsky. Molecules 2012, 17, 4604–4611. [Google Scholar] [CrossRef] [PubMed]
- Labandeira, C.C.; Sepkoski, J.J. Insect Diversity. Science 1993, 261, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agugliaro, F.M.; Muros, M.J.S.; Barroso, F.G.; Sanchez, A.M.; Rojo, S.; Banon, C.P. Insects for biodiesel production. Renew. Sustain. Energy Rev. 2012, 16, 3744–3753. [Google Scholar] [CrossRef]
- Barbara, K.A. American Cockroach, Periplaneta americana (Linnaeus) (Insecta: Blattodea: Blattidae). Featur. Creat. 2014, 141, 1–4. [Google Scholar]
- Kaya, M.; Baran, T. Description of a new surface morphology for chitin extracted from wings of cockroach (Periplaneta americana). Int. J. Biol. Macromol. 2015, 75, 7–12. [Google Scholar] [CrossRef]
- Wanule, D.; Balkhande, J.V.; Ratnakar, P.U.; Kulkarni, A.N.; Bhowate, C.S. Extraction and FTIR Analysis of Chitosan from American cockroach, Periplaneta americana. Int. J. Eng. Sci. Innov. Technol. 2014, 3, 299–304. [Google Scholar]
- Kang, S.C.; Park, S.; Lee, D.G. Purification and characterization of a novel chitinase from the entomopathogenic fungus, Metarhizium anisopliae. J. Invertebr. Pathol. 1999, 73, 276–281. [Google Scholar] [CrossRef]
- Harrigan, W. Laboratory Methods in Food Microbiology, 3rd ed.; Academic Press: London, UK, 1998. [Google Scholar]
- Yanai, K.; Takaya, N.; Kojima, N.; Horiuchi, H.; Ohta, A.; Takagi, M. Purification of two chitinases from Rhizopus oligosporus and isolation and sequencing of the encoding genes. J. Bacteriol. 1992, 174, 7398–7406. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Abuzaytoun, R. Chitin, chitosan, and co-products: Chemistry, production, applications, and health effects. Adv. Food Nutr. Res. 2005, 49, 93–135. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Baran, T.; Asan-Ozusaglam, M.; Cakmak, Y.S.; Tozak, K.O.; Mol, A.; Mentes, A.; Sezen, G. Extraction and characterization of chitin and chitosan with antimicrobial and antioxidant activities from cosmopolitan Orthoptera species (Insecta). Biotechnol. Bioprocess Eng. 2015, 20, 168–179. [Google Scholar] [CrossRef]
- Lee, E.A.; Pan, C.H.; Son, J.M.; Kim, S.I. Isolation and charcterization of Basic exochitinase from Leaf Extract of Rehmannia glutinosa. Biosci. Biotechnol. Biochem. 1999, 63, 1781–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajomsang, W.; Gonil, P. Preparation and characterization of α-chitin from cicada sloughs. Mater. Sci. Eng. 2010, 30, 357–363. [Google Scholar] [CrossRef]
- Majtán, J.; Bíliková, K.; Markovič, O.; Gróf, J.; Kogan, G.; Šimúth, J. Isolation and characterization of chitin from bumblebee (Bombus terrestris). Int. J. Biol. Macromol. 2007, 40, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Mujtaba, M.; Ehrlich, H.; Salaberria, A.M.; Baran, T.; Amemiya, C.T.; Labidi, J. On chemistry of γ-chitin. Carbohydr. Polym. 2017, 176, 177–186. [Google Scholar] [CrossRef]
- Zhang, M.; Haga, A.; Sekiguchi, H.; Hirano, S. Structure of insect chitin isolated from beetle larva cuticle and silkworm (Bombyx mori) pupa exuvia. Int. J. Biol. Macromol. 2000, 27, 99–105. [Google Scholar] [CrossRef]
- Chen, J.K.; Shen, C.R.; Liu, C.L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef] [Green Version]
- Engstrom, C.A.; Sarkodee-Adoo, C. The molecular biology of lymphoma. Semin. Oncol. Nurs. 1998, 14, 256–261. [Google Scholar] [CrossRef]
- Chen, J.K.; Shen, C.R.; Yeh, C.H.; Fang, B.S.; Huang, T.L.; Liu, C.L. N-Acetyl glucosamine obtained from chitin by chitin degrading factors in Chitinbacter tainanesis. Int. J. Mol. Sci. 2011, 12, 1187–1195. [Google Scholar] [CrossRef]
- Chern, L.L.; Stackebrandt, E.; Lee, S.F.; Lee, F.L.; Chen, J.K.; Fu, H.M. Chitinibacter tainanensis gen. nov., sp. nov., a chitin-degrading aerobe from soil in Taiwan. Int. J. Syst. Evol. Microbiol. 2004, 54, 1387–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.L.; Wu, S.T.; Yu, S.T.; Too, J.R. Screening of a Microbe to Degrade Chitin. Taiwan. J. Agric. Chem. Food Sci. 2005, 43, 410–418. [Google Scholar]
- Karthik, N.; Pandey, A.; Akanksha, K. Production, purification, and properties of fungal chitinases. Indian J. Exp. Biol. 2014, 11, 1025–1035. [Google Scholar]
- Limón, M.C.; Chacón, M.R.; Mejías, R.; Delgado-Jarana, J.; Rincón, A.M.; Codón, A.C.; Benítez, T. Increased antifungal and chitinase specific activities of Trichoderma harzianum CECT 2413 by addition of a cellulose binding domain. Appl. Microbiol. Biotechnol. 2004, 64, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Nampoothiri, K.M.; Baiju, T.V.; Sandhya, C.; Sabu, A.; George, S.; Ashok, P. Process optimization for antifungal chitinase production by Trichoderma harzianum. Process Biochem. 2004, 39, 1583–1590. [Google Scholar] [CrossRef]
- Wasli, A.S.; Salleh, M.M.; Abd-Aziz, S.; Hassan, O.; Mahadi, N.M. Medium optimization for chitinase production from Trichoderma virens using central composite design. Biotechnol. Bioprocess Eng. 2009, 14, 781–787. [Google Scholar] [CrossRef]
- Massimiliano, F.; Leuba, J.; Federico, F. Chitinolytic Enzyme Activity of Penicillium janthinellum P9 in Bench-Top Bioreactor. J. Ferment. Bioeng. 1998, 86, 620–623. [Google Scholar]
- El-Katatny, M.H.; Gudelj, M.; Robra, K.H.; Elnaghy, M.A.; Gübitz, G.M. Characterization of a chitinase and an endo-β-1,3-glucanase from Trichoderma harzianum Rifai T24 involved in control of the phytopathogen Sclerotium rolfsii. Appl. Microbiol. Biotechnol. 2001, 56, 137–143. [Google Scholar] [CrossRef]
- Sharaf, E.F. A potent chitinolytic activity of Alternaria alternata isolated from Egyptian black sand. Pol. J. Microbiol. 2005, 54, 145–151. [Google Scholar]
- Duo-Chuan, L.I.; Chen, S.; Jing, L.U. Purification and partial characterization of two chitinases from the mycoparasitic fungus Talaromyces flavus. Mycopathologia 2005, 159, 223–229. [Google Scholar] [CrossRef]
- Agrawal, T.; Kotasthane, A.S. Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. Springer Plus 2012, 1, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Strain | AUMC Number | Source of Isolation |
|---|---|---|
| Trichoderma harzianum Rifai | AUMC 5408 | Soil of Lens esculentus (lentil) plantation Assiut, 2009 |
| Mucor circinelloides van Tieghem | AUMC 6017 | Honeybee, Egypt, 2009 |
| Mucor circinelloides van Tieghem | AUMC 6027 | Cow dung, Sohag, 2009 |
| Sample | C% | H% | N% | DA% |
|---|---|---|---|---|
| * PDEX | 32.48 | 5.74 | 3.12 | 306.3 |
| * OSPC | 43.84 | 6.93 | 6.33 | 103 |
| Crab chitin | 43.49 | 6.99 | 6.12 | 107.6 |
| Shrimp chitin | 43.32 | 6.85 | 6.41 | 94 |
| pH | * N | Mean NAG * (±STD) g/L | Grouping | |||
|---|---|---|---|---|---|---|
| 4 | 3 | 12.785 ± 0.77 | A | |||
| 3 | 3 | 10.545 ± 1.3 | A | |||
| 6 | 3 | 7.549 ± 0.85 | B | |||
| 5 | 3 | 5.950 ± 0.23 | B | C | ||
| 7 | 3 | 4.335 ± 1.13 | C | D | ||
| 2 | 3 | 2.1425 ± 0.56 | D | |||
| 8 | 3 | 1.965 ± 0.23 | D | |||
| Strain/Fermentation Mechanism | * N | Mean Bioethanol * (±Std) g/L | Grouping | |
|---|---|---|---|---|
| M. circinelloides AUMC 6017/Indirect | 3 | 11.92 ± 0.06807 | A | |
| M. circinelloides AUMC 6017/Direct | 3 | 11.22 ± 0.312 | A | |
| M. circinelloides AUMC 6027/Indirect | 3 | 2.68 ± 0.03512 | B | |
| M. circinelloides AUMC 6027/Direct | 3 | 1.94 ± 0.05033 | B | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamal, M.; Adly, E.; Alharbi, S.A.; Khaled, A.S.; Rady, M.H.; Ibrahim, N.A. Exploring Simplified Methods for Insect Chitin Extraction and Application as a Potential Alternative Bioethanol Resource. Insects 2020, 11, 788. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110788
Kamal M, Adly E, Alharbi SA, Khaled AS, Rady MH, Ibrahim NA. Exploring Simplified Methods for Insect Chitin Extraction and Application as a Potential Alternative Bioethanol Resource. Insects. 2020; 11(11):788. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110788
Chicago/Turabian StyleKamal, Mahmoud, Eslam Adly, Sulaiman Ali Alharbi, Amany Soliman Khaled, Magda Hassan Rady, and Nevin Ahmed Ibrahim. 2020. "Exploring Simplified Methods for Insect Chitin Extraction and Application as a Potential Alternative Bioethanol Resource" Insects 11, no. 11: 788. https://0-doi-org.brum.beds.ac.uk/10.3390/insects11110788