Population Dynamics of Aedes aegypti and Aedes albopictus in Two Rural Villages in Southern Mexico: Baseline Data for an Evaluation of the Sterile Insect Technique

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
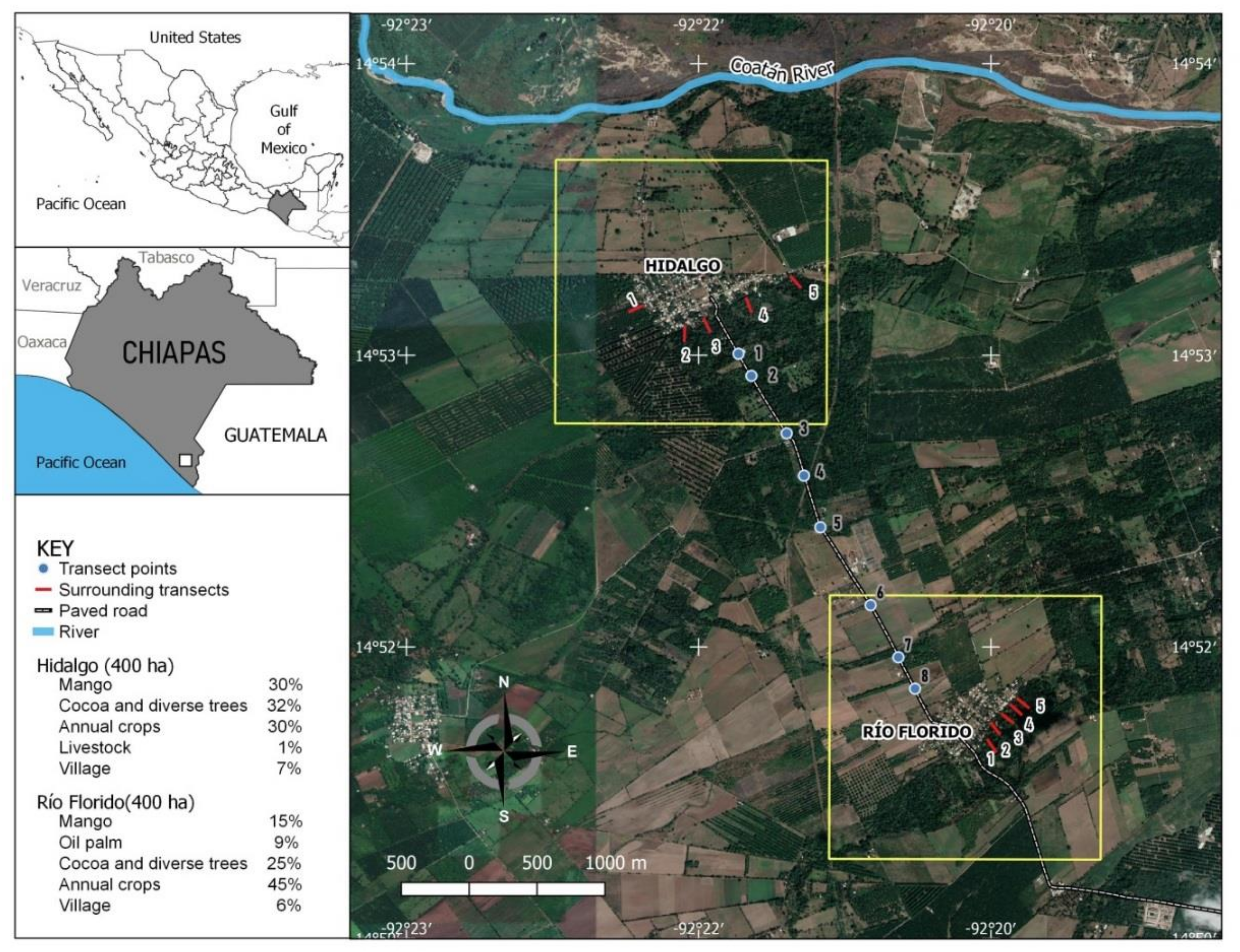
2.1. Study Sites
2.2. Methods for Mosquito Collections
2.3. Experimental Design
2.3.1. Sampling in Houses of the Pilot Communities
2.3.2. Sampling in Surrounding Areas
2.4. Laboratory Processing of Samples
2.5. Statistical Analyses
3. Results
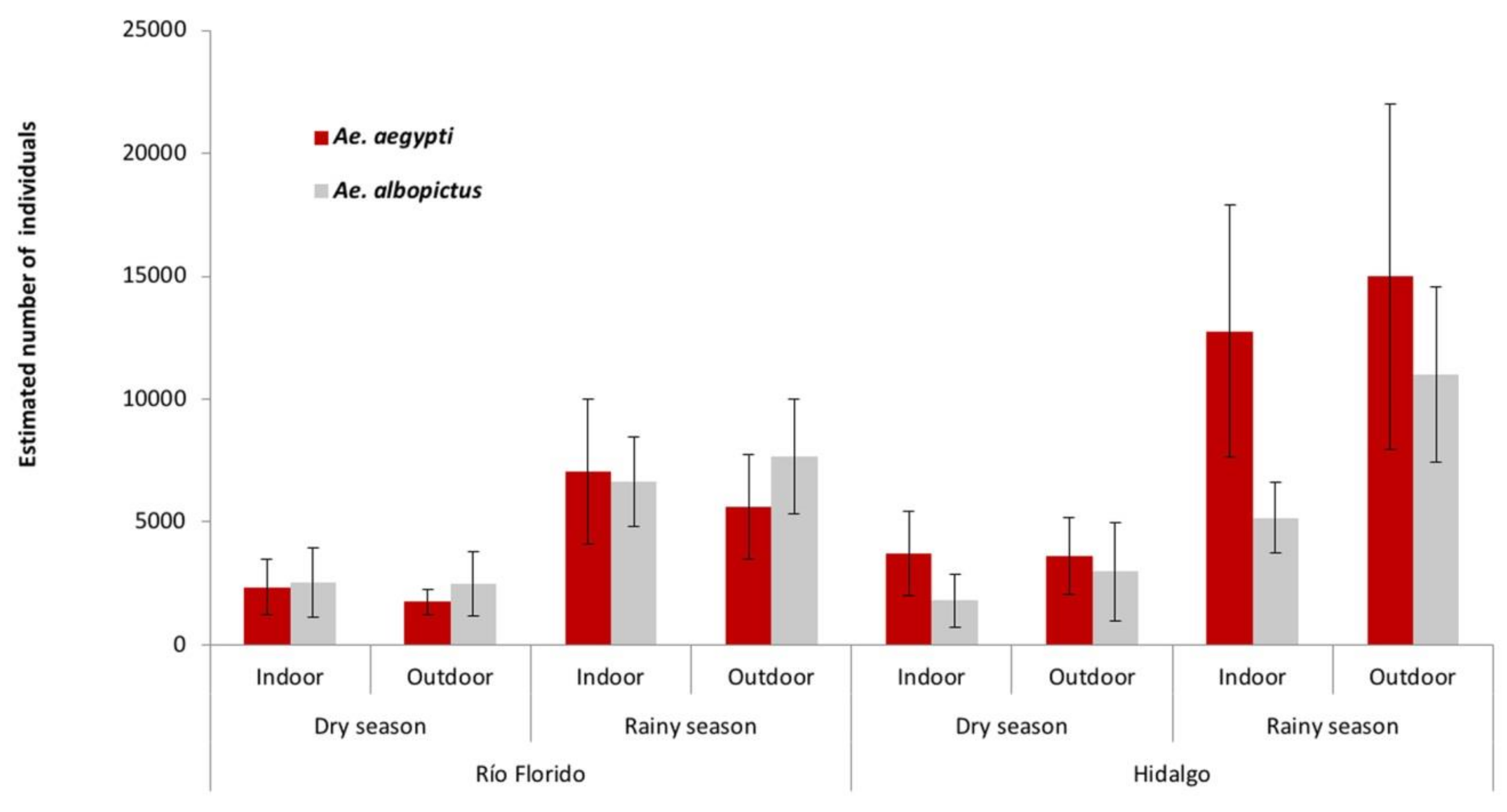
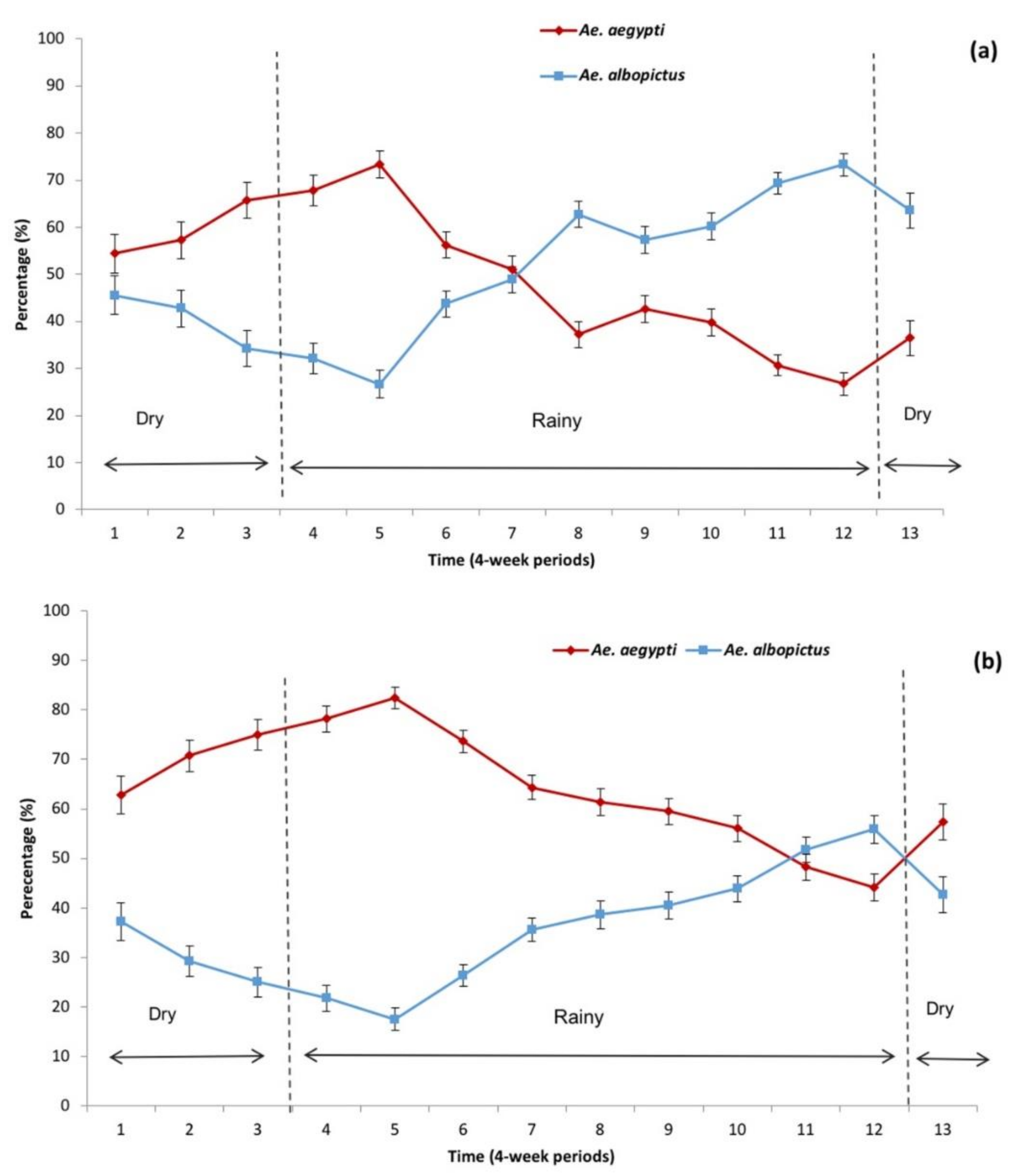
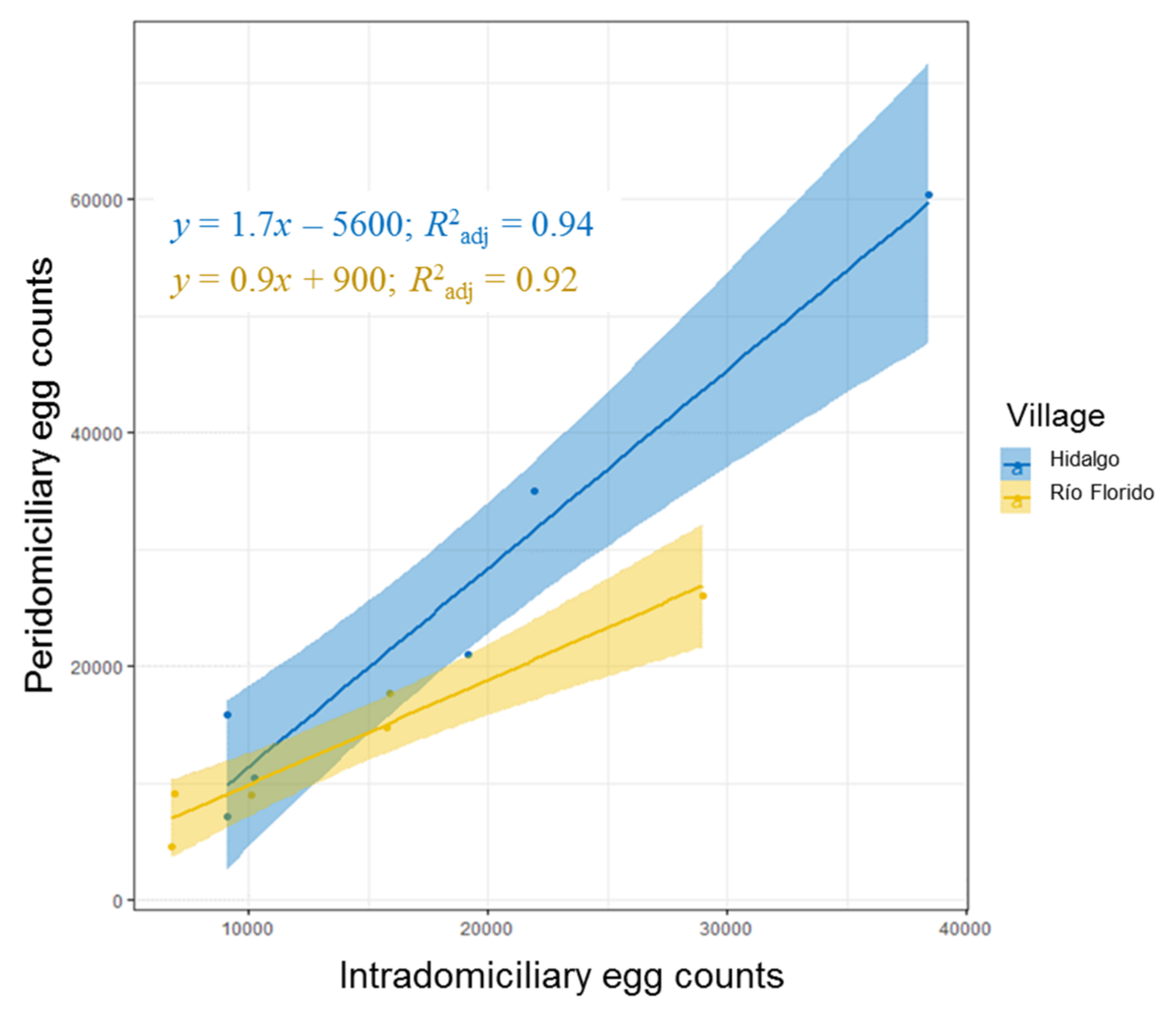
3.1. Sampling in and Around Houses
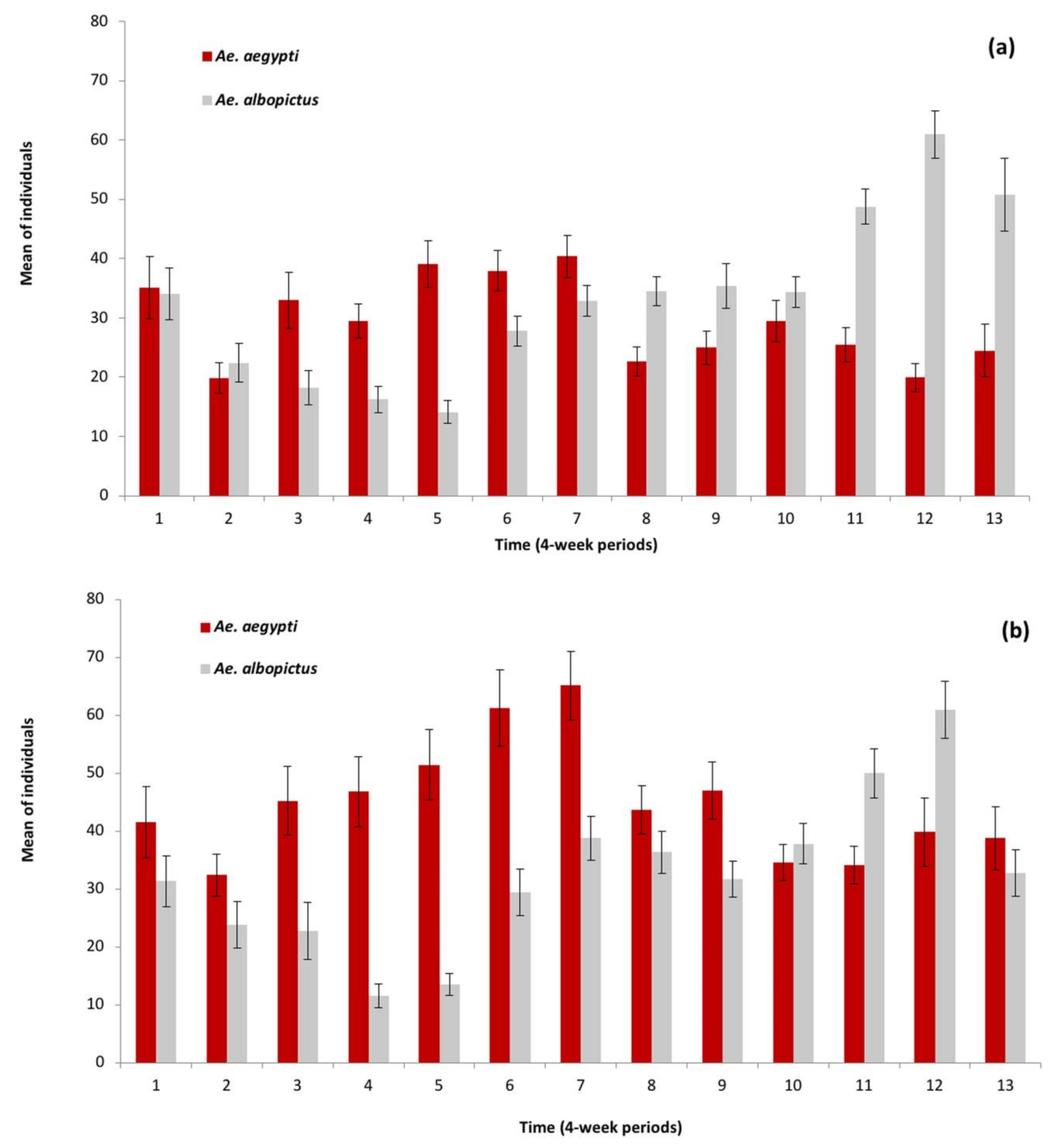
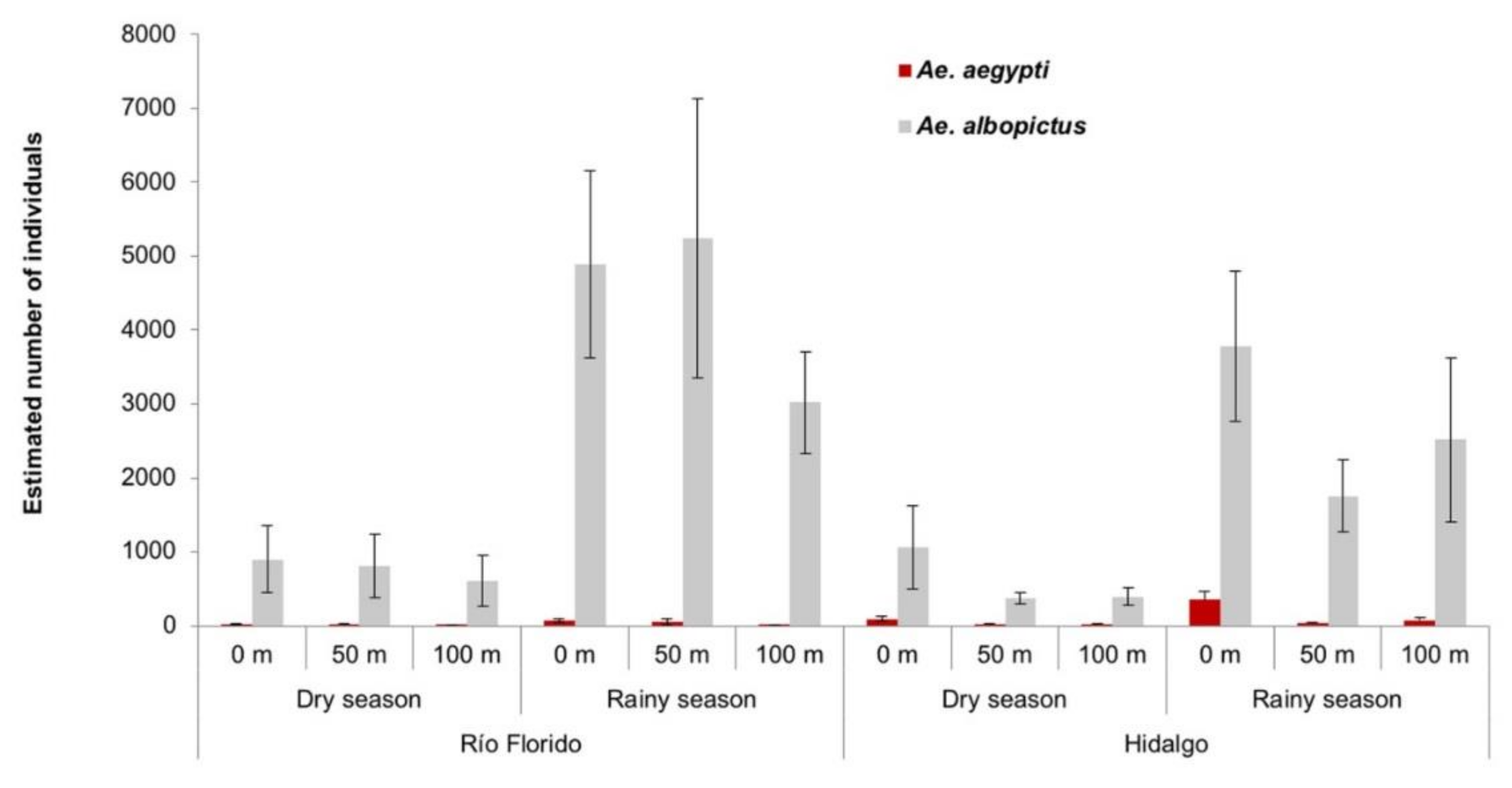
3.2. Sampling in Surrounding Transects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Fernández-Salas, I.; Danis-Lozano, R.; Casas-Martínez, M.; Ulloa, A.; Bond, J.G.; Marina, C.F.; Lopez-Ordóñez, T.; Elizondo-Quiroga, A.; Torres-Monzón, J.A.; Díaz-González, E.E. Historical inability to control Aedes aegypti as a main contributor of fast dispersal of chikungunya outbreaks in Latin America. Antivir. Res. 2015, 124, 30–42. [Google Scholar] [CrossRef]
- Rey, J.R.; Lounibos, P. Ecología de Aedes aegypti y Aedes albopictus en América y la transmisión de enfermedades. Biomédica 2015, 35, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Díaz-González, E.E.; Kautz, T.F.; Dorantes-Delgado, A.; Malo-García, I.R.; Laguna-Aguilar, M.; Langsjoen, R.M.; Chen, R.; Auguste, D.I.; Sánchez-Casas, R.M.; Danis-Lozano, R.; et al. First report of Aedes aegypti transmission of chikungunya virus in the Americas. Am. J. Trop. Med. Hyg. 2015, 93, 1325–1329. [Google Scholar] [CrossRef] [Green Version]
- Guerbois, M.; Fernandez-Salas, I.; Azar, S.R.; Danis-Lozano, R.; Alpuche-Aranda, C.M.; Leal, G.; Garcia-Malo, I.R.; Diaz-Gonzalez, E.E.; Casas-Martinez, M.; Rossi, S.L.; et al. Outbreak of Zika virus infection, Chiapas State, Mexico, 2015, and first confirmed transmission by Aedes aegypti mosquitoes in the Americas. J. Infect. Dis. 2016, 214, 1349–1356. [Google Scholar] [CrossRef] [Green Version]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef]
- Lozano-Fuentes, S.; Kenney, J.L.; Varnado, W.; Byrd, B.D.; Burkhalter, K.L.; Savage, H.M. Susceptibility and vectorial capacity of American Aedes albopictus and Aedes aegypti (Diptera: Culicidae) to American Zika virus strains. J. Med. Entomol. 2019, 56, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Tabachnick, W.J. Evolutionary genetics and arthropod-borne disease: The yellow fever mosquito. Am. Entomol. 1991, 37, 14–26. [Google Scholar] [CrossRef]
- Soper, F.L. Elimination of urban yellow-fever in Americas through eradication of Aedes aegypti. Am. J. Public Health Nat. Health 1963, 53, 7–16. [Google Scholar] [CrossRef]
- Novo, S. Breve historia y antología sobre la fiebre amarilla. Salud Pub. Mex. 1995, 37, 99–102. [Google Scholar]
- Ibáñez-Bernal, S.; Gómez-Dantés, H. Los vectores del dengue en México: Una revisión crítica. Salud Pub. Mex. 1995, 37, S53–S63. [Google Scholar]
- Wilson, M.A.; Chen, L.H. Dengue in the Americas. Dengue Bull. 2002, 26, 44–61. [Google Scholar]
- Navarrete, J.; Vásquez, J.L.; Vásquez, J.A.; Gómez, H. Epidemiologia del dengue y dengue hemorrágico en el Instituto Mexicano del Seguro Social (IMSS). Rev. Peruana Epidemiol. 2002, 10, 1–7. [Google Scholar]
- Sprenger, D.; Wuithiranyagool, T. The discovery and distribution of Aedes albopictus in Harris County, Texas. J. Am. Mosq. Control Assoc. 1986, 2, 217–219. [Google Scholar] [PubMed]
- Hawley, W.A.; Reiter, P.; Copeland, R.S.; Pumpuni, C.B.; Craig, G.B., Jr. Aedes albopictus in North America: Probable introduction in used tires from northern Asia. Science 1987, 236, 1114–1116. [Google Scholar] [CrossRef]
- Forattini, O.P. Identificacao de Aedes (Stegomyia) albopictus (Skuse) no Brasil. Ver. Saude Pub. de Sao Paulo 1986, 20, 244–245. [Google Scholar] [CrossRef]
- Moore, C.G.; Mitchell, C.J. Aedes albopictus in the United States: Ten-year presence and public health implications. Emerg. Infect. Dis. 1997, 3, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez-Bernal, S.; Martínez-Campos, C. Aedes albopictus in Mexico. J. Am. Mosq. Contr. Assoc. 1994, 10, 231–232. [Google Scholar]
- Casas-Martinez, M.; Torres-Estrada, J. First evidence of Aedes albopictus (Skuse) in southern Chiapas in Mexico. Emerg. Infect. Dis. 2003, 9, 606–607. [Google Scholar] [CrossRef]
- Bond, J.G.; Casas-Martinez, M.; Quiroz-Martinez, H.; Novelo-Gutierrez, R.; Marina, C.F.; Ulloa, A.; Orozco-Bonilla, A.; Muñoz, M.; Williams, T. Diversity of mosquitoes of medical importance and the aquatic insects associated with their oviposition sites along the Pacific coast Mexico. Parasit. Vect. 2014, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Bond, J.G.; Moo-Llanes, D.A.; Ortega-Morales, A.I.; Marina, C.F.; Casas-Martínez, M. Diversity and distribution potential of culicids of medical importance of the Yucatan Peninsula, Mexico. Salud Púb. Méx. 2020, 62, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Baak-Baak, C.M.; Cigarroa-Toledo, N.; Arana-Guardia, R.; Chi Chim, W.A.; Chan Orilla, J.A.; Machain-Williams, C.; Torres-Chable, O.M.; Ortega-Morales, A.I.; Moo-Llanes, D.A.; Elizondo-Quiroga, A.; et al. Mosquito fauna associated with Aedes aegypti (Diptera: Culicidae) in Yucatán State of southeastern México, and checklist with new records. Florida Entomol. 2016, 99, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Pech-May, A.; Moo-Llanes, D.A.; Puerto-Avila, M.B.; Casas, M.; Danis-Lozano, R.; Ponce, G.; Tun-Ku, E.; Pinto-Castillo, J.F.; Villegas, A.; Ibáñez-Piñon, C.R.; et al. Population genetics and ecological niche of invasive Aedes albopictus in México. Acta Trop. 2016, 157, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Torres-Chable, O.M.; Baak-Baak, C.M.; Cigarroa-Toledo, N.; Zaragoza-Vera, C.V.; Arjona-Jimenez, G.; Moreno-Perez, L.G.; Machain-Williams, C.; Garcia-Rejon, J.E. Mosquito fauna in home environments of Tabasco, Mexico. Southwest. Entomol. 2017, 42, 969–982. [Google Scholar] [CrossRef]
- Williams, T.; Farfán, J.L.; Mercado, G.; Valle, J.; Abella, A.; Marina, C.F. Efficacy of spinosad granules and lambda-cyhalothrin contrasts with reduced performance of temephos for control of Aedes spp. in vehicle tires in Veracruz, Mexico. Insects 2019, 10, 242. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Morales, A.; Zavortink, T.; Huerta-Jiménez, H.; Ibáñez-Bernal, S.; Siller-Rodríguez, Q. The mosquitoes (Diptera: Culicidae) of Hidalgo state, Mexico. Acta Trop. 2019, 189, 94–103. [Google Scholar] [CrossRef]
- Reiter, P.; Gubler, D.J. Surveillance and control of urban dengue vectors. In Dengue and Dengue Hemorrhagic Fever; Gubler, D.A., Kuno, G., Eds.; CAB International: London, UK, 1997; pp. 425–462. [Google Scholar]
- Horstick, O.; Runge-Ranzinger, S.; Nathan, M.B.; Kroeger, A. Dengue vector control services: How do they work? A systematic literature review and country case studies. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 379–386. [Google Scholar] [CrossRef]
- Esu, E.; Lenhart, A.; Smith, L.; Horstick, O. Effectiveness of peridomestic space spraying with insecticide on dengue transmission; systematic review. Trop. Med. Int. Health 2010, 15, 619–631. [Google Scholar] [CrossRef]
- Rodriguez, M.M.; Bisset, J.A.; Fernandez, D. Levels of insecticide resistance and resistance mechanisms in Aedes aegypti from some Latin American countries. J. Am. Mosq. Control Assoc. 2007, 23, 420–429. [Google Scholar] [CrossRef]
- Nauen, R. Insecticide resistance in disease vectors of public health importance. Pest Manag. Sci. 2007, 63, 628–633, PMID: 17533649. [Google Scholar] [CrossRef] [PubMed]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef] [PubMed]
- Vontas, J.; Kioulos, E.; Pavlidi, N.; Morou, E.; della Torre, A.; Ranson, H. Insecticide resistance in the major dengue vectors Aedes albopictus and Aedes aegypti. Pestic. Biochem. Physiol. 2012, 104, 126–131. [Google Scholar] [CrossRef]
- Monath, T.P. Vaccines against diseases transmitted from animals to humans: A one health paradigm. Vaccine 2013, 31, 5321–5338. [Google Scholar] [CrossRef]
- Knipling, E.F. The Basic Principles of Insect Population Suppression and Management; United States Department of Agriculture (USDA): Washington, DC, USA, 1979; p. 659. [Google Scholar]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique Principles and Practice in Area Wide Integrated Pest Management; Springer: Dordrecht, The Netherlands, 2005; p. 787. [Google Scholar]
- Liedo, P.; Enkerlin, W.; Hendrichs, J. La Técnica del Insecto Estéril. In Moscas de la Fruta: Fundamentos y Procedimientos Para su Manejo; Montoya, P., Toledo, J., Hernández, E., Eds.; S y G Editores: Ciudad de México, Mexico, 2010; pp. 243–255. [Google Scholar]
- Liedo, P.; Enkerlin, W.; Hendrichs, J. La Técnica del Insecto Estéril. In Moscas de la Fruta: Fundamentos y Procedimientos para su Manejo, 2nd ed.; Montoya, P., Toledo, J., Hernández, E., Eds.; S y G Editores: Ciudad de México, Mexico, 2020; pp. 357–373. [Google Scholar]
- Hendrichs, J.; Robinson, A.S.; Cayol, J.P.; Enkerlin, W. Medfly area wide sterile insect technique programmes for prevention, suppression or eradication: The importance of mating behavior studies. Fla. Entomol. 2002, 85, 1–13. [Google Scholar] [CrossRef]
- Oliva, C.F.; Jacquet, M.; Gilles, J.; Lemperiere, G.; Maquart, P.O.; Quilici, S.; Schooneman, F.; Vreysen, M.J.; Boyer, S. The sterile insect technique for controlling populations of Aedes albopictus (Diptera: Culicidae) on Reunion Island: Mating vigour of sterilized males. PLoS ONE 2012, 7, e49414, PMID: 23185329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marina, C.F.; Bond, J.G.; Muñoz, J.; Valle, J.; Quiroz-Martínez, H.; Torres-Monzón, J.A.; Williams, T. Comparison of novaluron, pyriproxyfen, spinosad (granules and tablets) and temephos as larvicides against Aedes aegypti in oviposition traps and domestic water tanks in Chiapas, Mexico. Salud Pub. Mex. 2020, 62, 424–431. [Google Scholar] [CrossRef]
- García, E. Modificaciones al sistema de clasificación climática de Köppen, para adaptarlo a las condiciones de la República Mexicana. In Instituto de Geografía; UNAM: Mexico City, Mexico, 1988; p. 205. [Google Scholar]
- Bond, J.G.; Ramírez-Osorio, A.; Marina, C.F.; Fernández-Salas, I.; Liedo, P.; Dor, A.; Williams, T. Efficiency of two larval diets for mass-rearing of the mosquito Aedes aegypti. PLoS ONE 2017, 12, e0187420. [Google Scholar] [CrossRef] [Green Version]
- Rueda, L.M. Pictorial keys for the identification of mosquitoes (Diptera: Culicidae) associated with dengue virus transmission. Zootaxa 2004, 589, 1–60. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: www.r-project.org (accessed on 18 December 2020).
- Lourenço-de-Oliveira, R.; Castro, M.G.; Braks, M.A.; Lounibos, L.P. The invasion of urban forest by dengue vectors in Rio de Janeiro. J. Vector Ecol. 2004, 29, 94–100. [Google Scholar]
- Honório, N.A.; Castro, M.G.; Barros, F.S.M.; Magalhães, M.A.F.M.; Sabroza, P.C. The spatial distribution of Aedes aegypti and Aedes albopictus in a transition zone, Rio de Janeiro, Brazil. Cad Saude Publica 2009, 25, 1203–1214. [Google Scholar] [CrossRef]
- Savage, H.; Niebylski, M.; Smith, G.; Mitchell, C.; Craig, G. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) at a temperate North American site. J. Med. Entomol. 1993, 30, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Faraji, A.; Egizi, A.; Fonseca, D.M.; Unlu, I.; Crepeau, T.; Healy, S.P.; Gaugler, R. Comparative host feeding patterns of the Asian Tiger Mosquito, Aedes albopictus, in urban and suburban northeastern USA and implications for disease transmission. PLoS Negl. Trop. Dis. 2014, 8, e3037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, K.R.; Williamson, D.; Carrière, Y.; Reyes-Castro, P.A.; Haenchen, S.; Hayden, M.H.; Jeffrey Gutierrez, E.; Ernst, K.C. Socioeconomic and human behavioral factors associated with Aedes aegypti (Diptera: Culicidae) immature habitat in Tucson, AZ. J. Med. Entomol. 2018, 55, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, J.M.; Bond, J.G.; Macotela, M.E.; Facchinelli, L.; Valerio, L.; Brown, D.M.; Scott, T.W.; James, A.A. A regulatory structure for working with genetically modified mosquitoes: Lessons from Mexico. PLoS Negl. Trop. Dis. 2014, 8, e2623. [Google Scholar] [CrossRef] [PubMed]
- Unlu, I.; Farajollahi, A.; Strickman, D.; Fonseca, D.M. Crouching tiger, hidden trouble: Urban sources of Aedes albopictus (Diptera: Culicidae) refractory to source-reduction. PLoS ONE 2013, 8, e77999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey, J.R.; Nishimura, N.; Wagner, B.; Braks, M.A.H.; O’Connell, S.M.; Lounibos, L.P. Habitat segregation of mosquito arbovirus vectors in South Florida. J. Med. Entomol. 2006, 43, 1134–1141. [Google Scholar] [CrossRef]
- Juliano, S.; O’Meara, G.; Morrill, J.; Cutwa, M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Harrington, L.C.; Edman, J.D.; Scott, T.W. Why do female Aedes aegypti (Diptera: Culicidae) feed preferentially and frequently on human blood? J. Med. Entomol. 2001, 38, 411–422. [Google Scholar] [CrossRef]
- Jiang, Y. Survival of overwintering Aedes albopictus eggs under natural conditions in North-Central Florida. J. Am. Mosq. Control Assoc. 2018, 34, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Tippelt, L.; Werner, D.; Kampen, H. Tolerance of three Aedes albopictus strains (Diptera: Culicidae) from different geographical origins towards winter temperatures under field conditions in northern Germany. PLoS ONE 2019, 14, e0219553. [Google Scholar] [CrossRef] [Green Version]
- Rey, J.R.; O’Connell, S.M. Oviposition by Aedes aegypti and Aedes albopictus: Influence of congeners and of oviposition site characteristics. J. Vector Ecol. 2014, 39, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.J.; Kaufman, P.E.; Hogsette, J.A.; Kline, D.L. The effects of larval habitat quality on Aedes albopictus skip oviposition. J. Am. Mosq. Control Assoc. 2015, 31, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Sota, T.; Mogi, M.; Hayamizu, E. Seasonal distribution and habitat selection by Aedes albopictus and Aedes riversi (Diptera: Culicidae) in northern Kyushu, Japan. J. Med. Entomol. 1992, 29, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Chareonviriyaphap, T.; Akratanakul, P.; Nettanomsak, S.; Huntamai, S. Larval habitats and distribution patterns of Aedes aegypti (Linnaeus) and Aedes albopictus (Skuse), in Thailand. Southeast Asian J. Trop. Med. Public Health 2003, 34, 529–535. [Google Scholar]
- Simard, F.; Nchoutpouen, E.; Toto, J.C.; Fontenille, D. Geographic distribution and breeding site preference of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) in Cameroon, Central Africa. J. Med. Entomol. 2005, 42, 726–731. [Google Scholar] [CrossRef]
- Kumari, R.; Kumar, K.; Chauhan, L.S. First dengue virus detection in Aedes albopictus from Delhi, India: Its breeding ecology and role in dengue transmission. Trop. Med. Int. Health 2011, 16, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, K.; Sudheesh Kumar, T.K.; Nujum, Z.T.; Umarul, F.; Kuriakose, A. A study on container breeding mosquitoes with special reference to Aedes (Stegomyia) aegypti and Aedes albopictus in Thiruvananthapuram district, India. J. Vector Borne Dis. 2014, 51, 27–32. [Google Scholar]
- Rozilawati, H.; Tanaselvi, K.; Nazni, W.A.; Mohd Masri, S.; Zairi, J.; Adanan, C.R.; Lee, H.L. Surveillance of Aedes albopictus Skuse breeding preference in selected dengue outbreak localities in Peninsular Malaysia. Trop. Biomed. 2015, 32, 49–64. [Google Scholar]
- Preechaporn, W.; Mullica, J.; Jaroensutasinee, K. The larval ecology of Aedes aegypti and Ae. albopictus in three topographical areas of southern Thailand. WHO Reg. Office South-East Asia. Dengue Bull. 2006, 30, 204–213. [Google Scholar]
- Dieng, H.; Saifur, R.G.; Hassan, A.A.; Salmah, M.C.; Boots, M.; Satho, T.; Jaal, Z.; AbuBakar, S. Indoor-breeding of Aedes albopictus in northern peninsular Malaysia and its potential epidemiological implications. PLoS ONE 2010, 5, e11790. [Google Scholar] [CrossRef] [Green Version]
- Lounibos, L.P. Competitive displacement and reduction. J. Am. Mosq. Control Assoc. 2007, 23, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, L.; Kendell, D.; Robertson, D.; Livdahl, T.; Khatchikian, C. Aedes aegypti and Aedes albopictus in Bermuda: Extinction, invasion and extinction. Biol. Invasions 2010, 12, 3277–3288. [Google Scholar] [CrossRef]
- Braks, M.A.H.; Honorio, N.A.; Lounibos, L.P.; Lourenco-De-Oliveira, R.; Juliano, S.A. Interspecific competition between two invasive species of container mosquitoes, Aedes aegypti and Aedes albopictus (Diptera: Culicidae), in Brazil. Ann. Entomol. Soc. Am. 2004, 97, 130–139. [Google Scholar] [CrossRef]
- Aliabadi, B.K.; Juliano, S.A. Escape from gregarine parasites affects the competitive impact of an invasive mosquito. Biol. Invas. 2002, 4, 283–297. [Google Scholar] [CrossRef]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Tripet, F.; Lounibos, L.P.; Robbins, D.; Moran, J.; Nishimura, N.; Blosser, E.M. Competitive reduction by satyrization? Evidence for interspecific mating in nature and asymmetric reproductive competition between invasive mosquito vectors. Am. J. Trop. Med. Hyg. 2011, 85, 265–270. [Google Scholar] [CrossRef]
- Bargielowski, I.E.; Lounibos, L.P.; Carrasquilla, M.C. Evolution of resistance to satyrization through reproductive character displacement in populations of invasive dengue vectors. Proc. Nat. Acad. Sci. USA 2013, 110, 2888–2892. [Google Scholar] [CrossRef] [Green Version]
- Juliano, S.A. Coexistence, exclusion, or neutrality? A meta-analysis of competition between Aedes albopictus and resident mosquitoes. Isr. J. Ecol. Evol. 2010, 56, 325–351. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aedes aegypti | Aedes albopictus | |||||
|---|---|---|---|---|---|---|
| Year and Season of Sample | Mean Rainfall (mm) | Location in Village | Total Eggs Collected in Ovitraps | Total Eggs Tested in Laboratory | Number of Larvae Reared (%) | Number of Larvae Reared (%) |
| 2016 Dry | 2.1 | Indoor | 5595 | 1781 | 408 (49.5) | 417 (50.5) |
| Outdoor | 6801 | 1864 | 411 (52.4) | 373 (47.6) | ||
| 2016 Rainy | 363.5 | Indoor | 15,773 | 8437 | 1436 (42.0) | 1983 (58.0) |
| Outdoor | 14,749 | 7883 | 1145 (39.6) | 1750 (60.4) | ||
| 2016–2017 Dry | 13.0 | Indoor | 4363 | 2601 | 685 (55.5) | 550 (44.5) |
| Outdoor | 4462 | 2378 | 551 (49.6) | 559 (50.4) | ||
| 2017 Rainy | 342.6 | Indoor | 15,916 | 7614 | 2785 (48.3) | 2983 (51.7) |
| Outdoor | 17,749 | 8735 | 2558 (37.9) | 4200 (62.1) | ||
| 2017–2018 Dry | 5.0 | Indoor | 13,860 | 5482 | 1834 (46.3) | 2125 (53.7) |
| Outdoor | 11,322 | 5481 | 1335 (35.0) | 2474 (65.0) | ||
| 2018 Rainy | 315.2 | Indoor | 28,957 | 13,302 | 5800 (55.8) | 4601 (44.2) |
| Outdoor | 26,076 | 12,228 | 4464 (46.0) | 5234 (54.0) | ||
| Totals | 165,623 | 77,786 | 23,412 | 27,249 |
| Ae. aegypti | Ae. albopictus | ||||
|---|---|---|---|---|---|
| Year and Season of Sample | Location in Village | Total Eggs Collected in Ovitraps | Total Eggs Tested in Laboratory | Number of Larvae Reared (%) | Number of Larvae Reared (%) |
| 2016 Dry | Indoor | 6882 | 2001 | 751 (78.0) | 212 (22.0) |
| Outdoor | 4924 | 1411 | 582 (77.7) | 167 (22.3) | |
| 2016 Rainy | Indoor | 19,166 | 9161 | 2549 (69.5) | 1116 (30.5) |
| Outdoor | 20,996 | 8388 | 1711 (52.8) | 1529 (47.2) | |
| 2016–2017 Dry | Indoor | 5107 | 3090 | 920 (67.2) | 449 (32.8) |
| Outdoor | 7978 | 3314 | 853 (59.2) | 587 (40.8) | |
| 2017 Rainy | Indoor | 21,906 | 9640 | 4540 (62.1) | 2775 (37.9) |
| Outdoor | 35,046 | 12,223 | 4339 (46.4) | 5003 (53.6) | |
| 2017–2018 Dry | Indoor | 16,386 | 6767 | 2939 (64.5) | 1621 (35.5) |
| Outdoor | 20,474 | 7112 | 2342 (49.2) | 2414 (50.8) | |
| 2018 Rainy | Indoor | 38,440 | 13,449 | 7912 (76.7) | 2406 (23.3) |
| Outdoor | 60,407 | 14,018 | 6548 (65.5) | 3442 (34.5) | |
| Totals | 257,712 | 90,574 | 35,986 | 21,721 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marina, C.F.; Bond, J.G.; Hernández-Arriaga, K.; Valle, J.; Ulloa, A.; Fernández-Salas, I.; Carvalho, D.O.; Bourtzis, K.; Dor, A.; Williams, T.; et al. Population Dynamics of Aedes aegypti and Aedes albopictus in Two Rural Villages in Southern Mexico: Baseline Data for an Evaluation of the Sterile Insect Technique. Insects 2021, 12, 58. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010058
Marina CF, Bond JG, Hernández-Arriaga K, Valle J, Ulloa A, Fernández-Salas I, Carvalho DO, Bourtzis K, Dor A, Williams T, et al. Population Dynamics of Aedes aegypti and Aedes albopictus in Two Rural Villages in Southern Mexico: Baseline Data for an Evaluation of the Sterile Insect Technique. Insects. 2021; 12(1):58. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010058
Chicago/Turabian StyleMarina, Carlos F., J. Guillermo Bond, Kenia Hernández-Arriaga, Javier Valle, Armando Ulloa, Ildefonso Fernández-Salas, Danilo O. Carvalho, Kostas Bourtzis, Ariane Dor, Trevor Williams, and et al. 2021. "Population Dynamics of Aedes aegypti and Aedes albopictus in Two Rural Villages in Southern Mexico: Baseline Data for an Evaluation of the Sterile Insect Technique" Insects 12, no. 1: 58. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010058