
Morphological Structure and Distribution of Hairiness on Different Body Parts of Apis mellifera with an Implication on Pollination Biology and a Novel Method to Measure the Hair Length



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
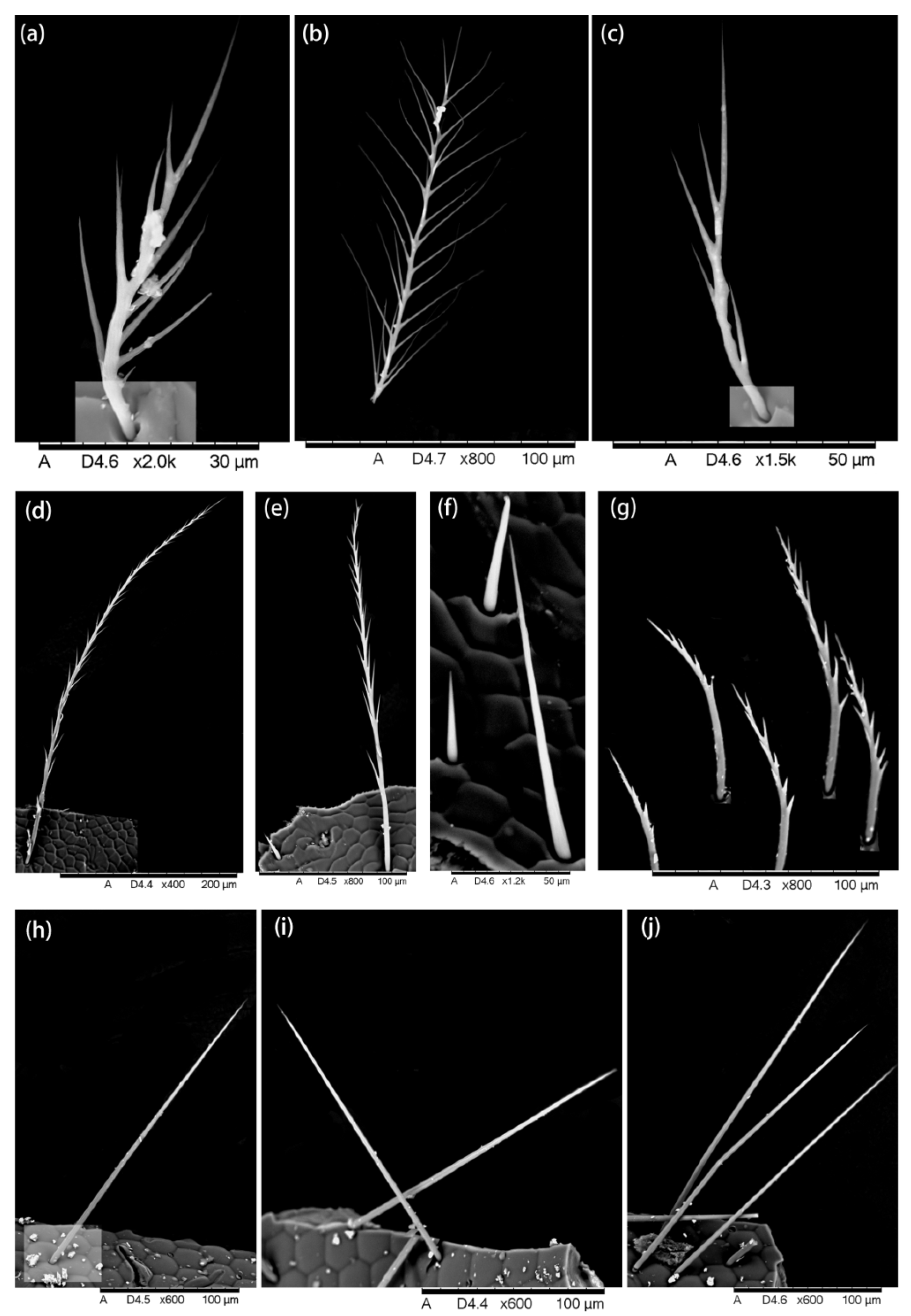
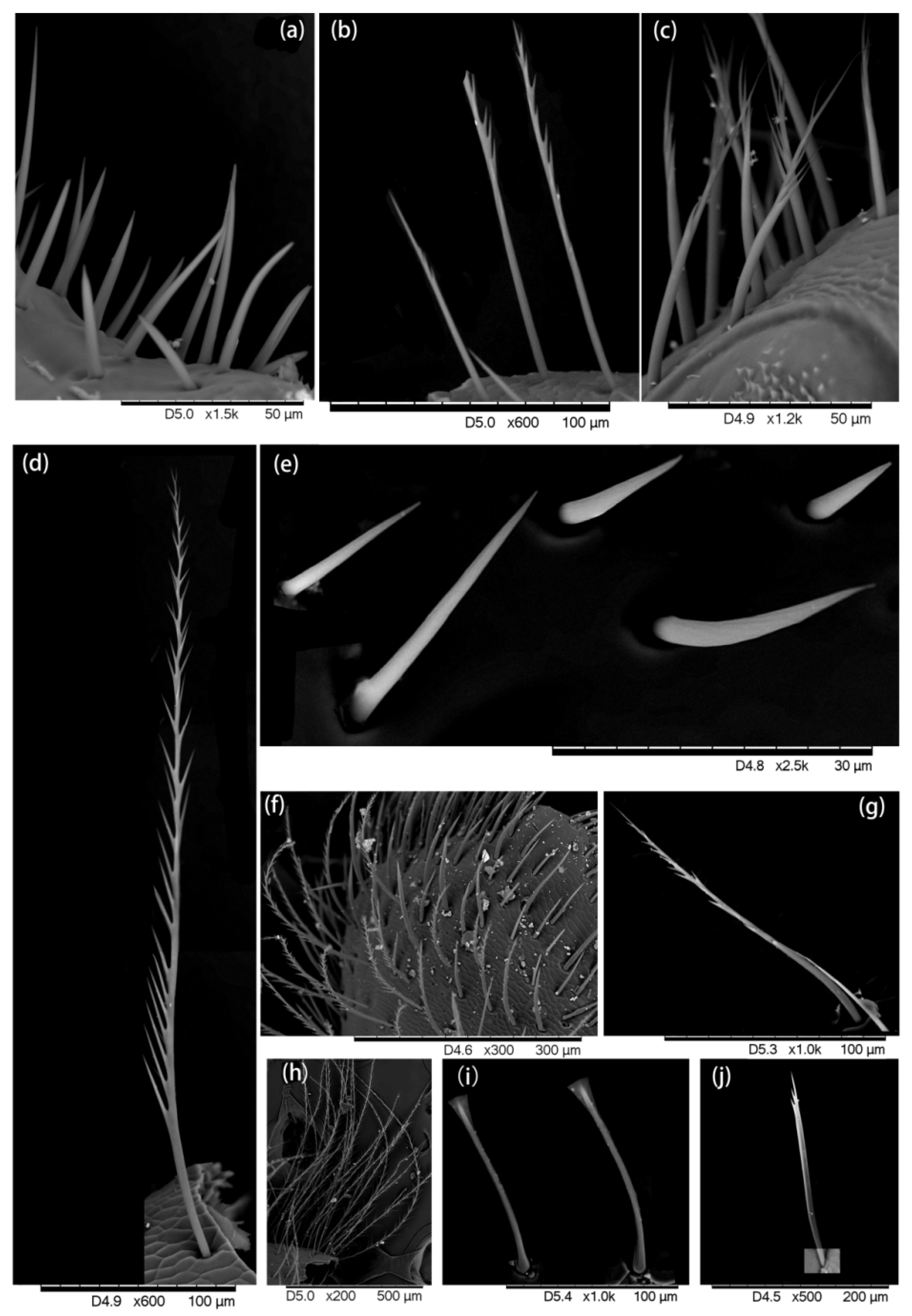
2.1. Hair Morphology
2.2. Slide Preparation Method
2.3. Hair Length
2.4. Statistical Analysis
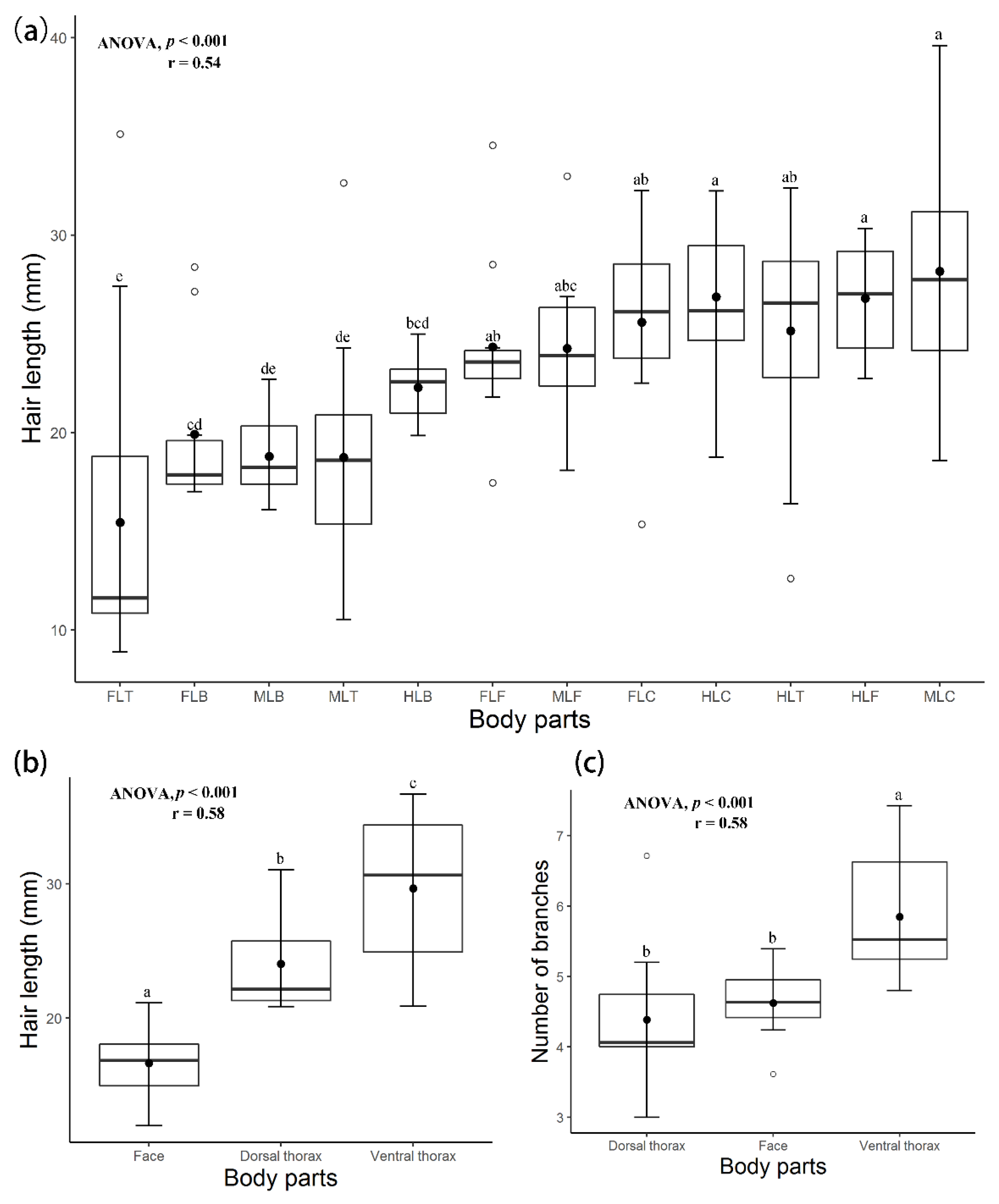
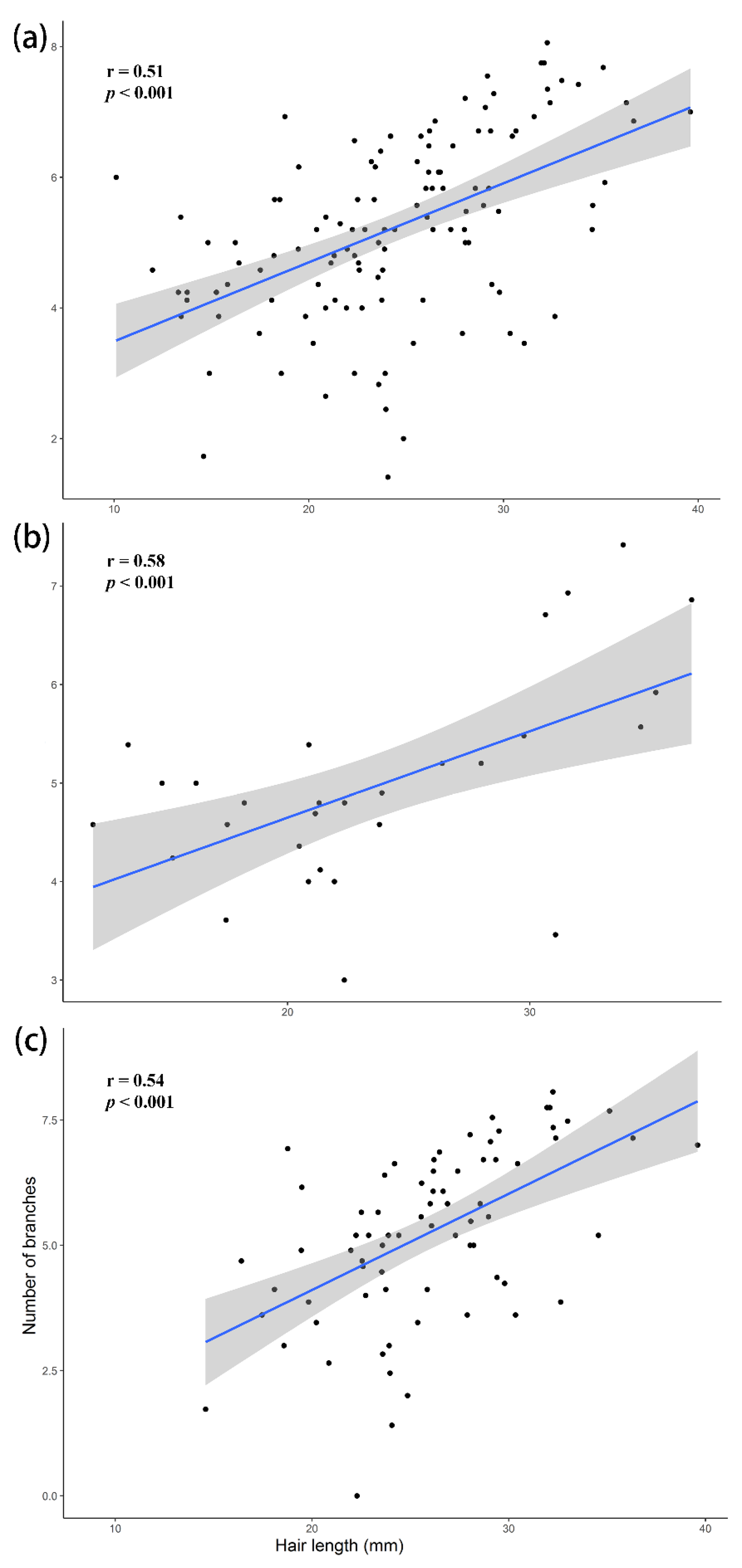
3. Results
3.1. Hair Morphology
3.2. Correlation between Hair Length and Number of Branches
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roquer-Beni, L.; Rodrigo, A.; Arnan, X.; Klein, A.M.; Fornoff, F.; Boreux, V.; Bosch, J. A novel method to measure hairiness in bees and other insect pollinators. Ecol. Evol. 2020, 10, 2979–2990. [Google Scholar] [CrossRef] [Green Version]
- Amador, G.J.; Matherne, M.; Waller, D.; Mathews, M.; Gorb, S.N.; Hu, D.L. Honey bee hairs and pollenkitt are essential for pollen capture and removal. Bioinspir. Biomim. 2017, 12, 026015. [Google Scholar] [CrossRef]
- Percival, M. Floral Biology; Elsevier Science: Burlington, MA, USA, 1965. [Google Scholar]
- Southwick, E.E. Bee hair structure and the effect of hair on metabolism at low temperature. J. Apic. Res. 1985, 24, 144–149. [Google Scholar] [CrossRef]
- Heinrich, B. The Hot-Blooded Insects: Strategies and Mechanisms of Thermoregulation; Springer: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- May, M.L. Insect Thermoregulation. Annu. Rev. Entomol. 1979, 24, 313–349. [Google Scholar] [CrossRef]
- Nock, C.A.; Vogt, R.J.; Beisner, B.E. Functional Traits; John Wiley & Sons: Chichester, UK, 2016; pp. 1–8. [Google Scholar]
- Kühsel, S. Pollinator Trait Diversity: Functional Implications at Different Land-Use Intensities and Environmental Conditions. Ph.D. Thesis, Technische Universität Darmstadt, Chichester, Germany, 2015. [Google Scholar]
- Moretti, M.; Dias, A.T.; De Bello, F.; Altermatt, F.; Chown, S.L.; Azcarate, F.M.; Bell, J.R.; Fournier, B.; Hedde, M.; Hortal, J. Handbook of protocols for standardized measurement of terrestrial invertebrate functional traits. Funct. Ecol. 2017, 31, 558–567. [Google Scholar] [CrossRef]
- Liu, R.; Chen, D.; Luo, S.; Xu, S.; Xu, H.; Shi, X.; Zou, Y. Quantifying pollination efficiency of flower-visiting insects and its application in estimating pollination services for common buckwheat. Agric. Ecosyst. Environ. 2020, 301, 107011. [Google Scholar] [CrossRef]
- Vergara-Amado, J.; Manzi, C.; Franco, L.M.; Contecha, S.C.; Marquez, S.J.; Solano-Iguaran, J.J.; Haro, R.E.; Silva, A.X. Effects of residual doses of neonicotinoid (imidacloprid) on metabolic rate of queen honey bees Apis mellifera (Hymenoptera: Apidae). Apidologie 2020, 51, 1091–1099. [Google Scholar] [CrossRef]
- Bartomeus, I.; Winfree, R. Pollinator declines: Reconciling scales and implications for ecosystem services. F1000Research 2013, 2, 146. [Google Scholar] [CrossRef] [Green Version]
- Powney, G.D.; Carvell, C.; Edwards, M.; Morris, R.K.A.; Roy, H.E.; Woodcock, B.A.; Isaac, N.J.B. Widespread losses of pollinating insects in Britain. Nat. Commun. 2019, 10, 1018. [Google Scholar] [CrossRef]
- Ekroos, J.; Kleijn, D.; Batáry, P.; Albrecht, M.; Báldi, A.; Blüthgen, N.; Knop, E.; Kovács-Hostyánszki, A.; Smith, H.G. High land-use intensity in grasslands constrains wild bee species richness in Europe. Biol. Conserv. 2020, 241, 108255. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Colla, S.R.; Gadallah, F.; Richardson, L.; Wagner, D.; Gall, L. Assessing declines of North American bumble bees (Bombus spp.) using museum specimens. Biodivers. Conserv. 2012, 21, 3585–3595. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Clough, Y.; Ekroos, J.; Baldi, A.; Batary, P.; Bommarco, R.; Gross, N.; Holzschuh, A.; Hopfenmuller, S.; Knop, E.; Kuussaari, M.; et al. Density of insect-pollinated grassland plants decreases with increasing surrounding land-use intensity. Ecol. Lett. 2014, 17, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Cirkovic, D.; Stevanovic, J.; Glavinic, U.; Aleksic, N.; Djuric, S.; Aleksic, J.; Stanimirovic, Z. Honey bee viruses in Serbian colonies of different strength. PeerJ 2018, 6, e5887. [Google Scholar] [CrossRef] [Green Version]
- Procházková, M.; Škubník, K.; Füzik, T.; Mukhamedova, L.; Přidal, A.; Plevka, P. Virion structures and genome delivery of honeybee viruses. Curr. Opin. Virol. 2020, 45, 17–24. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Münzenberg, U.; Bürger, C.; Thies, C.; Tscharntke, T. Scale-dependent effects of landscape context on three pollinator guilds. Ecology 2002, 83, 1421–1432. [Google Scholar] [CrossRef]
- Gisder, S.; Genersch, E. Viruses of commercialized insect pollinators. J. Invertebr. Pathol. 2017, 147, 51–59. [Google Scholar] [CrossRef]
- Ullah, A.; Tlak Gajger, I.; Majoros, A.; Dar, S.A.; Khan, S.; Kalimullah; Haleem Shah, A.; Nasir Khabir, M.; Hussain, R.; Khan, H.U.; et al. Viral impacts on honey bee populations: A review. Saudi J. Biol. Sci. 2021, 28, 523–530. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, J.M.; Hogendoorn, K. How protection of honey bees can help and hinder bee conservation. Curr. Opin. Insect Sci. 2021, 46, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.B.; Williams, A.; Osborne, J.L.; Shaw, R.F. Shared traits make flies and bees effective pollinators of oilseed rape (Brassica napus L.). Basic Appl. Ecol. 2018, 32, 66–76. [Google Scholar] [CrossRef]
- Morse, R.A.; Calderone, N.W. The value of honey bees as pollinators of US crops in 2000. Bee Cult. 2000, 128, 1–15. [Google Scholar]
- Southwick, E.E.; Southwick, L., Jr. Estimating the economic value of honey bees (Hymenoptera: Apidae) as agricultural pollinators in the United States. J. Econ. Entomol. 1992, 85, 621–633. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Partap, U.; Ya, T. The Human Pollinators of Fruit Crops in Maoxian County, Sichuan, China. Mt. Res. Dev. 2012, 32, 176–186. [Google Scholar] [CrossRef]
- Stavert, J.R.; Linan-Cembrano, G.; Beggs, J.R.; Howlett, B.G.; Pattemore, D.E.; Bartomeus, I. Hairiness: The missing link between pollinators and pollination. PeerJ 2016, 4, e2779. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Sun, J.; Cai, B.; Wu, Y. Phyllocnistis podocarpa sp. nov. (Lepidoptera, Gracillariidae), a buddhist pine leaf-miner from Japan: Taxonomy, DNA barcodes, damage and parasitoids. Zootaxa 2018, 4422, 558–568. [Google Scholar] [CrossRef]
- Willmer, P. Pollination and Floral Ecology; Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Massey, F.J., Jr. The Kolmogorov-Smirnov test for goodness of fit. J. Am. Stat. Assoc. 1951, 46, 68–78. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Peat, J.; Darvill, B.; Ellis, J.; Goulson, D. Effects of climate on intra-and interspecific size variation in bumble-bees. Funct. Ecol. 2005, 19, 145–151. [Google Scholar] [CrossRef]
- Roberts, R.B.; Vallespir, S.R. Specialization of hairs bearing pollen and oil on the legs of bees (Apoidea: Hymenoptera). Ann. Entomol. Soc. Am. 1978, 71, 619–627. [Google Scholar] [CrossRef]
- Haider, M.; Dorn, S.; Sedivy, C.; Müller, A. Phylogeny and floral hosts of a predominantly pollen generalist group of mason bees (Megachilidae: Osmiini). Biol. J. Linn. Soc. 2014, 111, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Mcgregor, S. Beekeeping in the United States; Publisher Agricultural Research Service, United States Department of Agriculture: Washington, DC, USA, 1967.
- Wasserman, D.; Nash, D.J. Variation in body size, hair length, and hair density in the deer mouse Peromyscus maniculatus along an altitudinal gradient. Ecography 1979, 2, 115–118. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Parts | Hair Length (mm) | Number of Branches | |
|---|---|---|---|
| Mean ± Standard Error | Mean ± Standard Error | ||
| Face | 16.65 ± 0.92 | 4.63 ± 0.15 | |
| Thorax | Dorsal surface | 24.05 ± 1.25 | 4.38 ± 0.33 |
| Ventral surface | 29.67 ± 1.80 | 5.85 ± 0.29 | |
| Abdomen | Dorsal surface | 15.69 ± 1.25 | 4.64 ± 0.33 |
| Ventral surface | 23.49 ± 1.30 | 5.87 ± 0.21 | |
| Foreleg | Coxa | 25.59 ± 1.48 | 4.74 ± 0.91 |
| Trochanter | 18.76 ± 2.85 | 3.03 ± 1.04 | |
| Femur | 24.34 ± 1.42 | 2.70 ± 0.78 | |
| Tibia | 12.16 ± 0.84 | 0 | |
| Basitarsus | 19.91 ± 1.34 | 0 | |
| Mid-leg | Coxa | 28.17 ± 2.028 | 4.73 ± 0.80 |
| Trochanter | 21.06 ± 1.57 | 0.85 ± 0.57 | |
| Femur | 24.27 ± 1.31 | 4.42 ± 0.65 | |
| Tibia | 16.43 ± 1.18 | 1.27 ± 0.58 | |
| Basitarsus | 18.90 ± 0.68 | 0 | |
| Hind leg | Coxa | 26.88 ± 1.29 | 6.86 ± 0.23 |
| Trochanter | 26.06 ± 1.54 | 5.69 ± 0.56 | |
| Femur | 26.81 ± 0.89 | 4.61 ± 0.25 | |
| Tibia | 24.25 ± 1.88 | 0 | |
| Basitarsus | 22.28 ± 0.51 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, K.A.; Liu, T. Morphological Structure and Distribution of Hairiness on Different Body Parts of Apis mellifera with an Implication on Pollination Biology and a Novel Method to Measure the Hair Length. Insects 2022, 13, 189. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020189
Khan KA, Liu T. Morphological Structure and Distribution of Hairiness on Different Body Parts of Apis mellifera with an Implication on Pollination Biology and a Novel Method to Measure the Hair Length. Insects. 2022; 13(2):189. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020189
Chicago/Turabian StyleKhan, Kamal Ahmed, and Tengteng Liu. 2022. "Morphological Structure and Distribution of Hairiness on Different Body Parts of Apis mellifera with an Implication on Pollination Biology and a Novel Method to Measure the Hair Length" Insects 13, no. 2: 189. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020189