
The Effect of Trap Color on Catches of Monochamus galloprovincialis and Three Most Numerous Non-Target Insect Species


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Testing the Response of Immature and Mature M. galloprovincialis to Different Colors under Laboratory Conditions
2.2. Testing the Response of Mature M. galloprovincialis to Different Colors under Field Conditions
2.3. Statistical Analyses
3. Results
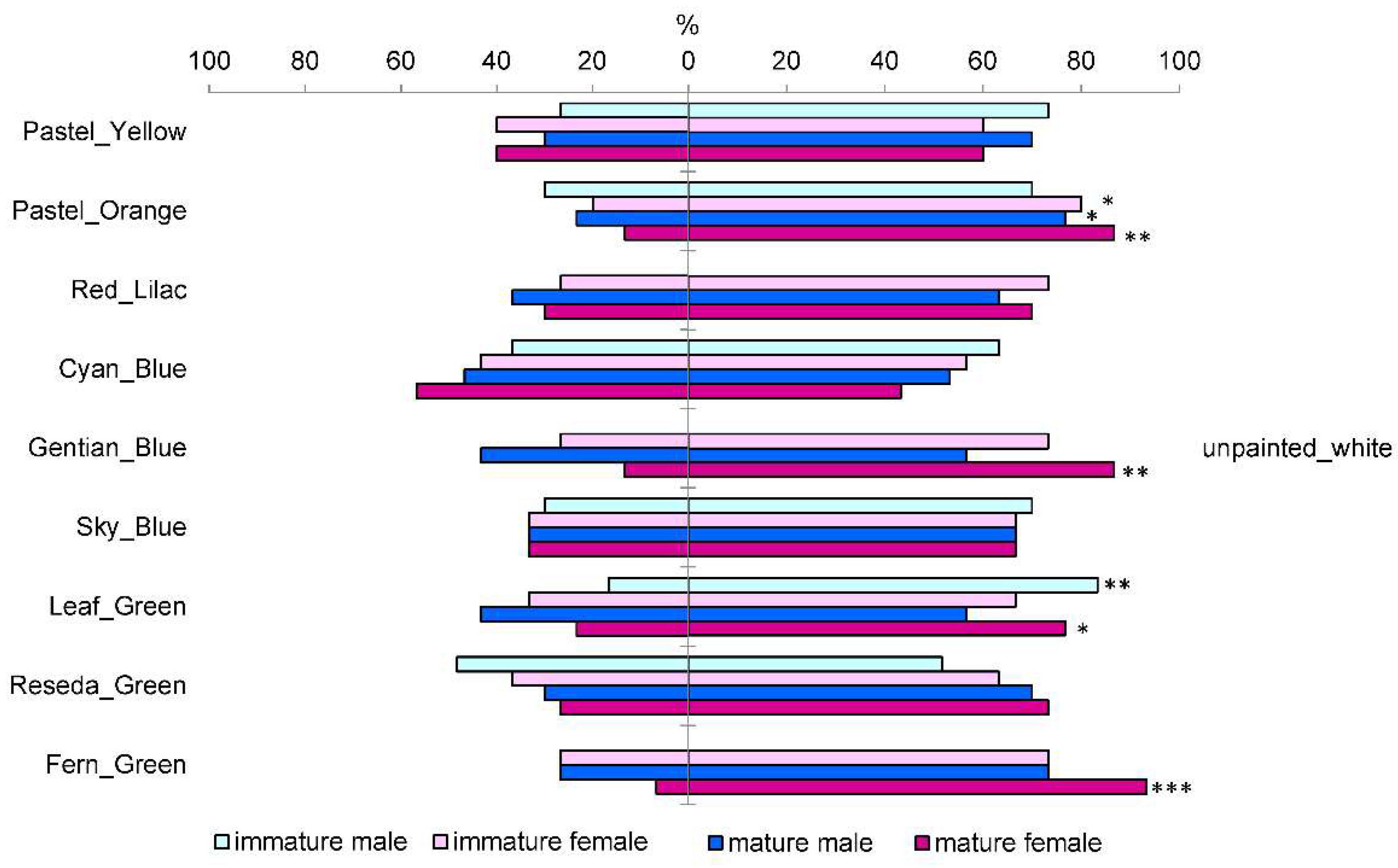
3.1. Response of Immature and Mature M. galloprovincialis to Colors in the Laboratory Studies
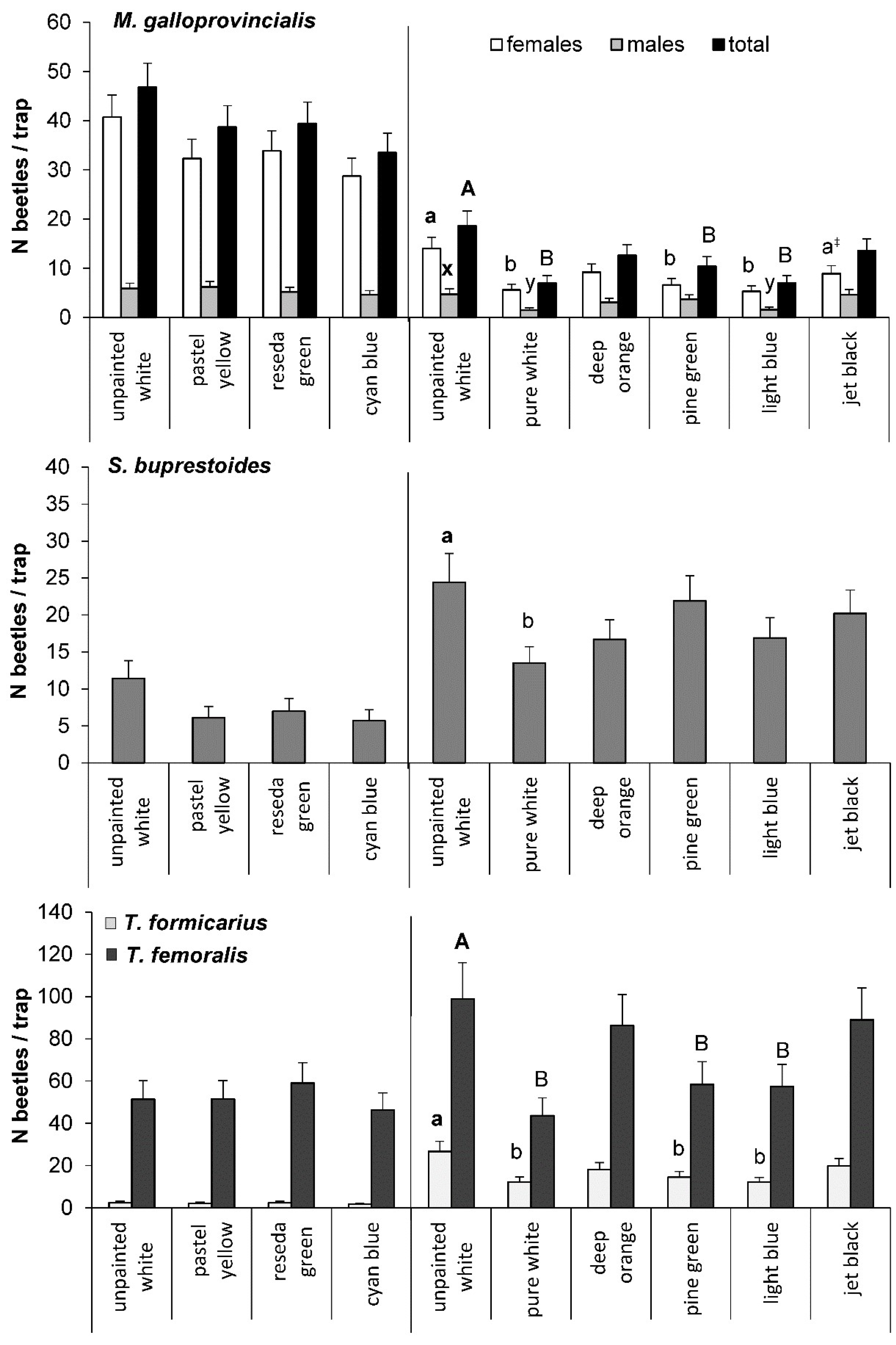
3.2. Effect of Trap Color in the Field Studies
3.2.1. M. galloprovincialis
3.2.2. S. buprestoides
3.2.3. Thanasimus spp.
4. Discussion
4.1. Response of M. galloprovincialis to Color
4.2. Response of Non-Target Insect Species to Color
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sousa, E.; Bravo, M.A.; Pires, J.; Naves, P.; Penas, A.C.; Bonifácio, L.; Mota, M. Bursaphelenchus xylophilus (Nematoda; Aphelenchoididae) associated with Monochamus galloprovincialis (Coleoptera; Cerambycidae) in Portugal. Nematology 2001, 3, 89–91. [Google Scholar]
- Mamiya, Y. The pine wood nematode. In Plant and Insect Nematodes; Nickle, W.R., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1984; pp. 589–627. [Google Scholar]
- Rutherford, T.A.; Webster, J.M. Distribution of pine wilt disease with respect to temperature in North America, Japan, and Europe. Can. J. For. Res. 1987, 17, 1050–1059. [Google Scholar] [CrossRef]
- Evans, H.; McNamara, D.; Braasch, H.; Chadouef, J.; Magnusson, C. Pest risk analysis (PRA) for the territories of the European Union (as PRA area) on Bursaphelenchus xylophilus and its vectors in the genus Monochamus. Bull. OEPP/EPPO Bull. 1996, 26, 199–249. [Google Scholar] [CrossRef]
- Mota, M.; Vieira, P. Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Springer: Dordrecht, Germany, 2008. [Google Scholar]
- European Union. 2012/535/EU: Commission Implementing Decision of 26 September 2012 on emergency measures to prevent the spread within the Union of Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle et al. (the pine wood nematode) (notified under document C(2012) 6543). Off. J. EU 2012, L 266, 42, (with later amendments). [Google Scholar]
- EPPO. EPPO Standards—Diagnostics. PM 7/4 (3) Bursaphelenchus xylophilus. Bull. OEPP/EPPO Bull. 2013, 43, 105–118. [Google Scholar] [CrossRef]
- De Groot, P.; Nott, R. Evaluation of traps of six different designs to capture pine sawyer beetles (Coleoptera: Cerambycidae). Agric. For. Entomol. 2001, 3, 107–111. [Google Scholar] [CrossRef]
- McIntosh, R.L.; Katinic, P.J.; Allison, J.D.; Borden, J.H.; Downey, D.L. Comparative efficacy of five types of trap for woodborers in the Cerambycidae, Buprestidae and Siricidae. Agric. For. Entomol. 2001, 3, 113–120. [Google Scholar] [CrossRef] [Green Version]
- De Groot, P.; Nott, R.W. Response of Monochamus (Col., Cerambycidae) and some Buprestidae to flight intercept traps. J. Appl. Entomol. 2003, 127, 548–552. [Google Scholar] [CrossRef]
- Allison, J.D.; Johnson, C.W.; Meeker, J.R.; Strom, B.L.; Butler, S.M. Effect of aerosol surface lubricants on the abundance and richness of selected forest insects captured in multiple-funnel and panel traps. J. Econ. Entomol. 2011, 104, 1258–1264. [Google Scholar] [CrossRef] [PubMed]
- Allison, J.D.; Bhandari, B.D.; McKenney, J.L.; Millar, J.G. Design factors that influence the performance of flight intercept traps for the capture of longhorned beetles (Coleoptera: Cerambycidae) from the subfamilies Lamiinae and Cerambycinae. PLoS ONE 2014, 9, e93203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, J.D.; Graham, E.E.; Poland, T.M.; Strom, B.L. Dilution of fluon before trap surface treatment has no effect on longhorned beetle (Coleoptera: Cerambycidae) captures. J. Econ. Entomol. 2016, 109, 1215–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, G.; Etxebeste, I.; Gallego, D.; Hall, D.R.; Jactel, H.; Pajares, J.A. Optimization of traps for live trapping of Pine Wood Nematode vector Monochamus galloprovincialis. J. Appl. Entomol. 2015, 139, 618–626. [Google Scholar] [CrossRef]
- Jaworski, T.; Plewa, R.; Dziuk, A.; Sukovata, L. Effect of trap type and PTFE treatment on catches of Monochamus galloprovincialis and three most numerous non-target insect species. Ann. For. Res. 2022; submitted. [Google Scholar]
- Campbell, S.A.; Borden, J.H. Additive and synergistic integration of multimodal cues of both hosts and non-hosts during host selection by woodboring insects. Oikos 2009, 118, 553–563. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Giannone, F.; Moino, S.; Rassati, D. Exploiting trap color to improve surveys of longhorn beetles. J. Pest Sci. 2021, 94, 871–883. [Google Scholar] [CrossRef]
- Moericke, V. Hostplant specific colour behaviour by Hyalopterus pruni (Aphididae). Entomol. Exp. Appl. 1969, 12, 524–534. [Google Scholar] [CrossRef]
- Bernays, E.A.; Chapman, R.F. Host-Plant Selection by Phytophagous Insects; Chapman & Hall: New York, NY, USA, 1994. [Google Scholar]
- Doering, T.F.; Chittka, L. Visual ecology of aphids—A critical review on the role of colours in host finding. Arthropod Plant Interact. 2007, 1, 3–16. [Google Scholar] [CrossRef] [Green Version]
- McCall, A.C.; Murphy, S.J.; Venner, C.; Brown, M. Florivores prefer white versus pink petal color morphs in wild radish, Raphanus sativus. Oecologia 2013, 172, 189–195. [Google Scholar] [CrossRef]
- Yang, H.; Wang, B.-X.; Yang, W.; Yang, C.-P.; Lu, L.; Chen, Z.-M. The role of vision in host-orientation and mate-finding behaviors of the white-striped longhorned beetle, Batocera lineolata. Entomol. News 2017, 126, 358–371. [Google Scholar] [CrossRef]
- Fukaya, M.; Akino, T.; Yasuda, T.; Yasui, H.; Wakamura, S. Visual and olfactory cues for mate orientation behaviour in male whitespotted longicorn beetle, Anoplophora malasiaca. Entomol. Exp. Appl. 2004, 111, 111–115. [Google Scholar] [CrossRef]
- Fukaya, M.; Akino, T.; Yasui, H.; Yasuda, T.; Wakamura, S.; Yamamura, K. Effect of size and color of female models for male mate orientation in the white spotted longicorn beetle Anoplophora malasiaca (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 2005, 40, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Kemp, D.J.; Rutowski, R.L. (2011) Chapter 2—The role of coloration in mate choice and sexual interactions in butterflies. In Advances in the Study of Behavior; Brockmann, H.J., Roper, T.J., Naguib, M., Mitani, J.C., Simmons, L.W., Eds.; Academic Press: Waltham, MA, USA, 2011; Volume 43, pp. 55–92. [Google Scholar] [CrossRef]
- Domingue, M.J.; Lelito, J.P.; Myrick, A.J.; Csóka, G.; Szöcs, L.; Imrei, Z.; Baker, T.C. Differences in spectral selectivity between stages of visually guided mating approaches in a buprestid beetle. J. Exp. Biol. 2016, 219, 2837–2843. [Google Scholar] [CrossRef] [Green Version]
- Tan, E.J.; Reid, C.A.M.; Symonds, M.R.E.; Juradi-Rivera, J.A.; Elgar, M.A. The role of life-history and ecology in the evolution of color patterns in Australian chrysomeline beetles. Front. Ecol. Evol. 2017, 5, 140. [Google Scholar] [CrossRef] [Green Version]
- Loeffler-Henry, K.; Kang, C.K.; Sherratt, T.N. Consistent associations between body size and hidden contrasting color signals across a range of insect taxa. Am. Nat. 2019, 194, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Soukupová, T.; Veselý, P.; Fuchs, R. Wariness to colour patterns: Birds versus European red-and-black insects. Ecol. Entomol. 2021, 46, 1157–1164. [Google Scholar] [CrossRef]
- Briscoe, A.D.; Chittka, L. The evolution of color vision in insects. Ann. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelber, A. Invertebrate colour vision. In Invertebrate Vision; Warrant, E., Nilsson, D.E., Eds.; Cambridge University Press: New York, NY, USA, 2006; pp. 250–290. [Google Scholar]
- Lord, N.P.; Plimpton, R.L.; Sharkey, C.R.; Suvorov, A.; Lelito, J.P.; Willardson, B.M.; Bybee, S.M. A cure for the blues: Opsin duplication and subfunctionalization for short-wavelength sensitivity in jewel beetles (Coleoptera: Buprestidae). BMC Evol. Biol. 2016, 16, 107. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, C.R.; Fujimoto, M.S.; Lord, N.P.; Shin, S.; McKeena, D.D.; Suvorov, A.; Martin, G.J.; Bybee, S.M. Overcoming the loss of blue sensitivity through opsin duplication in the largest animal group, beetles. Sci. Rep. 2017, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crook, D.J.; Francese, J.A.; Zylstra, K.E.; Fraser, I.; Sawyer, A.J.; Bartels, D.W.; Lance, D.R.; Mastro, V.C. Laboratory and field response of the emerald ash borer (Coleoptera: Buprestidae), to selected regions of the electromagnetic spectrum. J. Econ. Entomol. 2009, 102, 2160–2169. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-Y.; Stuart-Fox, D.; Wlaker, G.; Roberts, N.W.; Franklin, A.M. Insect visual sensitivity to long wavelengths enhances color contrast of insects against vegetation. Sci. Rep. 2022, 12, 982. [Google Scholar] [CrossRef]
- Domingue, M.J.; Csóka, G.; Tóth, M.; Vétek, G.; Pénzes, B.; Mastro, V.; Baker, T.C. Field observations of visual attraction of three European oak buprestid beetles toward conspecific and heterospecific models. Entomol. Exp. Appl. 2011, 140, 112–121. [Google Scholar] [CrossRef]
- Kim, K.-N.; Huang, Q.-Y.; Lei, C.-L. Advances in insect phototaxis and application to pest management: A review. Pest Manag. Sci. 2019, 75, 3135–3143. [Google Scholar] [CrossRef]
- Van der Kooi, C.J.; Stavenga, D.G.; Arikawa, K.; Belušič, G.; Kelber, A. Evolution of insect color vision: From spectral sensitivity to visual ecology. Ann. Rev. Entomol. 2021, 66, 435–461. [Google Scholar] [CrossRef]
- Shen, S.; Fan, Z.; Zhang, X.; Kong, X.; Liu, F.; Zhang, Z.; Zhang, X.; Hu, X.; Zhang, S. The characteristics of chemosensory and opsin genes in newly emerged and sexually mature Agrilus planipennis, an important quarantine forest beetle. Front. Genet. 2021, 11, 604757. [Google Scholar] [CrossRef]
- Koyama, N.; Gokan, N.; Matsui, M. The compound eye of the longicorn beetles (Coleoptera: Cerambycidae) with estimation of the activities by its structure. J. Fac. Tex. Sci. Tech. Shinshu Univ. 1975, 66, 1–16. [Google Scholar]
- Gokan, N.; Hosobuchi, K. Fine structure of the compound eyes of longicorn beetles (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1979, 14, 12–27. [Google Scholar] [CrossRef]
- Warrant, E.; Dacke, M. Visual navigation in nocturnal insects. Physiology 2016, 31, 182–192. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Jung, Y.H.; Lee, S.-M. Diel rhythmicity of field responses to synthetic pheromone lures in the pine sawyer Monochamus saltuarius. Insects 2021, 12, 441. [Google Scholar] [CrossRef]
- Nishimura, M. Daily observation on behaviors of Japanese pine sawyer adult, Monochamus alternatus Hope. J. Jpn. For. Soc. 1973, 55, 100–104. [Google Scholar] [CrossRef]
- Wen, C.; Ma, T.; Deng, Y.; Liu, C.; Liang, S.; Wen, J.; Wang, C.; Wen, X. Morphological and optical features of the apposition compound eye of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Micron 2020, 128, 102769. [Google Scholar] [CrossRef]
- Liu, B.; Xu, H.C.; Meng, J.G.; Sun, J.H.; Fan, J.T. Visual cues for the host-finding and mating locations of Monochamus alternatus (Coleoptera: Cerambycidae). J. Zhejiang Agr. For. Univ. 2012, 29, 617–620. [Google Scholar]
- Giffard, B.; David, G.; Joubard, B.; Piou, D.; Jactel, H. How do sex and sexual maturation influence the response of Monochamus galloprovincialis to host odours? J. Appl. Entomol. 2017, 141, 551–560. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Q.-H.; Wang, Y.; Liu, G.-T.; Zhou, X.; Niu, J.; Schlyter, F. Catching Ips duplicatus (Sahlberg) (Coleoptera: Scolytidae) with pheromone-baited traps: Optimal trap type, colour, height and distance to infestation. Pest Manag. Sci. 2010, 66, 2013–2219. [Google Scholar] [CrossRef]
- Francese, J.A.; Crook, D.J.; Fraser, I.; Lance, D.R.; Sawyer, A.J.; Mastro, V.C. Optimization of trap color for emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2010, 103, 1235–1241. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, T.; Su, Y.; Wang, Z.; Zhang, X.; Wang, S.; Liu, Y. Influence of trap color, type, and placement on capture efficacy for Protaetia brevitarsis (Coleoptera: Scarabaeidae). J. Econ. Entomol. 2021, 114, 225–230. [Google Scholar] [CrossRef]
- Rassati, D.; Toffolo, E.P.; Battisti, A.; Faccoli, M. Monitoring of the pine sawyer beetle Monochamus galloprovincialis by pheromone traps in Italy. Phytoparasitica 2012, 40, 329–336. [Google Scholar] [CrossRef]
- Encycolorpedia. Available online: https://encycolorpedia.com (accessed on 29 December 2021).
- Pajares, J.A.; Ibeas, F.; Diez, J.J.; Gallego, D. Attractive responses by Monochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. J. Appl. Entomol. 2004, 128, 633–638. [Google Scholar] [CrossRef]
- Stanisz, A. Przystępny Kurs Statystyki z Zastosowaniem STATISTICA PL na Przykładach z Medycyny Tom. 1. Statystyki Podstawowe; StatSoft: Kraków, Poland, 2006. [Google Scholar]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- StatSoft Inc. STATISTICA (Data Analysis Software System), version 10; StatSoft Inc.: Tulsa, OK, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- R Studio Team. R Studio: Integrated Development for R, version 1.3.1093; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.5.2-1. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 20 November 2020).
- Zuur, A.; Ieno, E.; Walker, N.; Saveliev, A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Mangiafico, S.S. Summary and Analysis of Extension Program Evaluation in R, Version 1.15.0. 2016. Available online: rcompanion.org/documents/RHandbookProgramEvaluation.pdf (accessed on 13 December 2018).
- Dodds, K.J.; Dubois, G.D.; Hoebeke, R.R. Trap type, lure placement, and habitat effects on Cerambycidae and Scolytinae (Coleoptera) catches in the Northeastern United States. J. Econ. Entomol. 2010, 103, 698–707. [Google Scholar] [CrossRef]
- Morewood, W.D.; Hein, K.E.; Katinic, P.J.; Borden, J.H. An improved trap for large wood-boring insects, with special reference to Monochamus scutellatus (Coleoptera: Cerambycidae). Can. J. For. Res. 2002, 32, 519–525. [Google Scholar] [CrossRef]
- Graham, E.E.; Poland, T.M. Efficacy of fluon conditioning for capturing cerambycid beetles in different trap designs and persistence on panel traps over time. J. Econ. Entomol. 2012, 105, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M. Trapping strategy for Monochamus sutor and Monochamus galloprovincialis: Potential vectors of the pine wood nematode in Scandinavia. Agric. For. Entomol. 2019, 21, 372–378. [Google Scholar] [CrossRef]
- Graham, E.E.; Mitchell, R.F.; Reagel, P.F.; Barbour, J.D.; Millar, J.G.; Hanks, L.M. Treating panel traps with a fluoropolymer enhances their efficiency in capturing cerambycid beetles. J. Econ. Entomol. 2010, 103, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Rassati, D.; Haack, R.A.; Knížek, M.; Faccoli, M. National trade can drive range expansion of bark- and wood-boring beetles. J. Econ. Entomol. 2018, 111, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Boone, C.K.; Sweeney, J.; Silk, P.; Hughes, C.; Webster, R.P.; Stephen, F.; MacLauchlan, L.; Bentz, B.; Drumont, A.; Zhao, B.; et al. Monochamus species from different continents can be effectively detected with the same trapping protocol. J. Pest Sci. 2019, 92, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Hoch, G.; Connell, J.; Roques, A. Testing multi-lure traps for surveillance of native and alien longhorn beetles (Coleoptera, Cerambycidae) at ports of entry and in forests in Austria. Manag. Biol. Invasions 2020, 11, 677–688. [Google Scholar] [CrossRef]
- Allison, J.D.; Redak, R.A. The impact of trap type and design features on survey and detection of bark and woodboring beetles and their associates: A review and meta-analysis. Annu. Rev. Entomol. 2017, 62, 127–146. [Google Scholar] [CrossRef] [PubMed]
- Petrice, T.R.; Haack, R.A. Comparison of different trap colors and types for capturing adult Agrilus (Coleoptera: Buprestidae) and other buprestids. Great Lakes Entomol. 2015, 48, 45–66. Available online: https://scholar.valpo.edu/tgle/vol48/iss1/4 (accessed on 23 December 2021).
- Mäntylä, E.; Kleier, S.; Lindstedt, C.; Kipper, S.; Hilker, M. Insectivorous birds are attracted by plant traits induced by insect egg deposition. J. Chem. Ecol. 2018, 44, 1127–1138. [Google Scholar] [CrossRef]
- Lang, M.; Kuusk, A.; Nilson, T.; Lükk, T.; Pehk, M.; Alm, G. Reflectance Spectra of Ground Vegetation in Sub-Boreal Forests [online]. Tartu Observatory, Tõravere, Estonia. Available online: http://www.aai.ee/bgf/ger2600/ (accessed on 29 December 2021).
- Campbell, S.A.; Borden, J.H. Bark reflectance spectra of conifers and angiosperms: Implications for host discrimination by coniferophagous bark and timber beetles. Can. Entomol. 2005, 137, 719–722. [Google Scholar] [CrossRef]
- Kerr, J.L.; Kelly, D.; Bader, M.K.-F.; Brockerhoff, E.G. Olfactory cues, visual cues, and semiochemical diversity interact during host location by invasive forest beetles. J. Chem. Ecol. 2017, 43, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Juola, J.; Hovi, A.; Rautiainen, M. Multiangular spectra of tree bark for common boreal tree species in Europe. Silva Fenn. 2020, 54, 10331. [Google Scholar] [CrossRef]
- Silva, D.; Salamanca, J.; Kyryczenko-Roth, V.; Alborn, H.T.; Rodriguez-Saona, C. Comparison of trap types, placement, and colors for monitoring Anthonomus musculus (Coleoptera: Curculionidae) adults in highbush blueberries. J. Insect Sci. 2018, 18, 19. [Google Scholar] [CrossRef] [PubMed]
- Beresford, D.V.; Sutcliffe, J.F. Studies on the effectiveness of coroplast sticky traps for sampling stable flies (Diptera: Muscidae), including a comparison to alsynite. J. Econ. Entomol. 2006, 99, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q. Sexual selection of Zorion guttigerum Westwood (Coleoptera: Cerambycidae: Cerambycinae) in relation to body size and color. J. Insect Behav. 2002, 15, 675–687. [Google Scholar] [CrossRef]
- Pajares, J.A.; Alvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.R.; Farman, D.I. Identification and field activity of a male-produced aggregation pheromone in the Pine Sawyer Beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, G.; Gallego, D.; David, G.; Bonifacio, L.; Jactel, H.; Sousa, E.; Pajares, J.A. Combining pheromone and kairomone for effective trapping of the pine sawyer beetle Monochamus galloprovincialis. J. Appl. Entomol. 2016, 140, 58–71. [Google Scholar] [CrossRef]
- Jurc, M.; Bojovic, S.; Fernández, M.F.; Jurc, D. The attraction of cerambycids and other xylophagous beetles, potential vectors of Bursaphelenchus xylophilus, to semio-chemicals in Slovenia. Phytoparasitica 2012, 40, 337–349. [Google Scholar] [CrossRef]
- Thomaes, A.; Drumont, A.; Warzée, N.; Grégoire, J.C.; Stassen, E.J.; Crevecoeur, L.; Berckvens, N.; Casteels, H.; Vijver, D.V.; Raemdonck, H. Ecology and distribution of Thanasimus formicarius (Linnaeus, 1758) and the newly discovered Thanasimus femoralis (Zetterstedt, 1828) in Belgium (Coleoptera: Cleridae). Bull. Ann. Soc. R. Belge Entomolog. 2017, 153, 206–214. [Google Scholar]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Magnani, G.; Rassati, D. Effect of trap color on captures of bark- and wood-boring beetles (Coleoptera; Buprestidae and Scolytinae) and associated predators. Insects 2020, 11, 749. [Google Scholar] [CrossRef] [PubMed]
- Shibata, E.; Sato, S.; Sakuratani, Y.; Sugimoto, T.; Kimura, F.; Ito, F. Cerambycid beetles (Coleoptera) lured to chemicals in forests of Nara prefecture, Central Japan. Ann. Entomol. Soc. Am. 1996, 89, 835–842. [Google Scholar] [CrossRef]
- Sweeney, J.; De Groot, P.; MacDonald, L.; Smith, S.; Cocquempot, C.; Kenis, M.; Gutowski, J.M. Host volatile attractants and traps for detection of Tetropium fuscum (F.), Tetropium castaneum L., and other longhorned beetles (Coleoptera: Cerambycidae). Environ. Entomol. 2004, 33, 844–854. [Google Scholar] [CrossRef] [Green Version]
- Kelsey, R.G.; Joseph, G. Ethanol and water in Pseudotsuga menziesii and Pinus ponderosa stumps. J. Chem. Ecol. 1999, 25, 2779–2792. [Google Scholar] [CrossRef]
- Strom, B.L.; Roton, L.M.; Goyer, R.A.; Meeker, J.R. Visual and semiochemical disruption of host finding in the southern pine beetle. Ecol. Appl. 1999, 9, 1028–1038. [Google Scholar] [CrossRef]
- Akkuzu, E.; Şahin, M.; Ugiş, A.; Bal, E. Assessment of trap color and trap height above the ground on the capture of Ips sexdentatus and Thanasimus formicarius. Šumar. List 2021, 145, 169–174. [Google Scholar] [CrossRef]
- Bakke, A.; Kvamme, T. Kairomone response in Thanasimus predators to pheromone components of Ips typographus. J. Chem. Ecol. 1981, 7, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M. Differences in responses to α-pinene and ethanol, and flight periods between the bark beetle predators Thanasimus femoralis and T. formicarius (Col.: Cleridae). For. Ecol. Manag. 2003, 177, 301–311. [Google Scholar] [CrossRef]
- Hulcr, J.; Ubik, K.; Vrkoč, J. The role of semiochemicals in tritrophic interactions between the spruce bark beetle Ips typographus, its predators and infested spruce. J. Appl. Entomol. 2006, 130, 275–283. [Google Scholar] [CrossRef]
- Wehnert, M.; Müller, M. ‘Allochthonous Kairomones’ in stands of European beech (Fagus sylvatica)—Approach for nature-based bark beetle management with clerid beetles (Thanasimus spp.). Biol. Control 2012, 62, 16–23. [Google Scholar] [CrossRef]
- Lubojacký, J.; Holuša, J. Effect of insecticide-treated trap logs and lure traps for Ips typographus (Coleoptera: Curculionidae) management on nontarget arthropods catching in Norway spruce stands. J. For. Sci. 2014, 60, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Gómez de Dios, M.Á.; Baena Ruiz, M.; Carrasco Gotarredona, Á.; Rodríguez Reviriego, S. Contribution to the knowledge of the checkered beetles (Coleoptera: Cleridae) of Almeria (Andalusia, Spain). Zool. Baetica 2015, 26, 101–144. [Google Scholar]
- Strom, B.L.; Goyer, R.A. Effect of silhouette color on trap catches of Dendroctonus frontalis (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 2001, 94, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Sakalian, V.; Langourov, M. Colour traps a method for distributional and ecological investigations of Buprestidae (Coleoptera). Acta Soc. Zool. Bohem. 2004, 68, 53–59. [Google Scholar]
- Rodriguez-Saona, C.R.; Byers, J.A.; Schiffhauer, D. Effect of color and height on captures of blunt-nosed and sharp-nosed leafhoppers (Hemiptera: Cicadellidae) and non-target arthropods in cranberry bogs. Crop Prot. 2012, 40, 132–144. [Google Scholar] [CrossRef]
- Sukovata, L.; Dziuk, A.; Parratt, M.; Bystrowski, C.; Dainton, K.; Polaszek, A.; Moore, R. The importance of trap type, trap colour and capture liquid for catching Dendrolimus pini and their impact on by-catch of beneficial insects. Agric. For. Entomol. 2020, 22, 319–327. [Google Scholar] [CrossRef]
- Bonifácio, L.; Henriques, J.; Inácio, M.L.; Naves, P.; Sousa, E.; Barroso, J.M.; Gonçalves, E.; Figueiredo, A.C. Monitoring and control of the pinewood nematode and its insect vector populations. Constraints and innovations. In Book of Abstracts, Proceedings of the International Symposium IUFRO on Pine Wilt Disease “PWD2020”, Orleans, France, 22–26 November 2021; p. 50. Available online: https://symposium.inrae.fr/pwd2020/content/download/3971/40932/version/3/file/Proceedings%20template_vf3.pdf (accessed on 5 January 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Color | Codes According to RAL or NCS 1 Color Systems | Approx. Wave- Length 2 | Light Reflectance Value 2 | Laboratory Tests | Field Studies | |||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 2019 | 2020 | ||||
| Pastel Yellow | 1034 | 585.4 | 42.1 | x 3 | x | x | ||
| Pastel Orange | 2003 | 592.7 | 33.2 | x | x | |||
| Deep Orange | 2011 | 591.7 | 27.4 | x | ||||
| Flame Red | 3000 | 608.0 | 9.9 | x | x | |||
| Light Pink | 3015 | 669.7 | 42.5 | x | ||||
| Red Lilac | 4001 | 551.5 | 14.9 | x | x | |||
| Pastel Violet | 4009 | 510.0 | 26.3 | x | x | |||
| Cyan Blue | S 0520-R90B 1 | 481.8 | 73.0 | x | x | x | ||
| Gentian Blue | 5010 | 478.3 | 7 | x | x | x | ||
| Light Blue | 5012 | 481.9 | 21.3 | x | ||||
| Sky Blue | 5015 | 480.4 | 17.5 | x | x | |||
| Cyan Green | S 4030-B90G 1 | 506.5 | 14.4 | x | ||||
| Leaf Green | 6002 | 553.5 | 8 | x | x | |||
| Reseda Green | 6011 | 562.3 | 18.3 | x | x | x | ||
| Fern Green | 6025 | 564.5 | 13.8 | x | x | |||
| Pine Green | 6028 | 518.4 | 7.4 | x | ||||
| Nut Brown | 8011 | 589.4 | 5.1 | x | ||||
| Jet Black | 9005 | 464.2 | 0.5 | x | ||||
| Pure White | 9010 | 575.7 | 83–96 | x | ||||
| White (unpainted) coroplast | x | x | x | |||||
| Test | Pair of Tested Colors | Immature Beetles | Mature Beetles | ||
|---|---|---|---|---|---|
| Males | Females | Males | Females | ||
| 1 | Pairs of contrasting colors | ||||
| Pastel Orange/Gentian Blue | 50 | 88 | 30 | 30 | |
| Pastel Orange/Flame Red | 20 | 20 | - | - | |
| Leaf Green/Gentian Blue | 22 | 20 | - | - | |
| Leaf Green/Flame Red | - | 20 | - | - | |
| Nut Brown/Flame Red | 30 | 30 | 30 | 30 | |
| 2 | Pairs of closely related colors | ||||
| Pastel Orange/Pastel Yellow | 30 | 30 | 30 | 30 | |
| Light Pink/Flame Red | 29 | 30 | 30 | 30 | |
| Pastel Violet/Red Lilac | 30 | 30 | 30 | 30 | |
| Sky Blue/Gentian Blue | 30 | 30 | 30 | 30 | |
| Leaf Green/Cyan green | 30 | 30 | 30 | 30 | |
| Fern Green/Reseda Green | 30 | 30 | 30 | 30 | |
| 3 | Colors tested against white (unpainted) coroplast | ||||
| Pastel Yellow | 30 | 30 | 30 | 30 | |
| Pastel Orange | 30 | 30 | 30 | 30 | |
| Red Lilac | - | 30 | 30 | 30 | |
| Cyan Blue | 30 | 30 | 30 | 30 | |
| Gentian Blue | - | 30 | 30 | 30 | |
| Sky Blue | 30 | 30 | 30 | 30 | |
| Leaf Green | 30 | 30 | 30 | 30 | |
| Reseda Green | 29 | 30 | 30 | 30 | |
| Fern Green | - | 30 | 30 | 30 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sukovata, L.; Dziuk, A.; Plewa, R.; Jaworski, T. The Effect of Trap Color on Catches of Monochamus galloprovincialis and Three Most Numerous Non-Target Insect Species. Insects 2022, 13, 220. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030220
Sukovata L, Dziuk A, Plewa R, Jaworski T. The Effect of Trap Color on Catches of Monochamus galloprovincialis and Three Most Numerous Non-Target Insect Species. Insects. 2022; 13(3):220. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030220
Chicago/Turabian StyleSukovata, Lidia, Aleksander Dziuk, Radosław Plewa, and Tomasz Jaworski. 2022. "The Effect of Trap Color on Catches of Monochamus galloprovincialis and Three Most Numerous Non-Target Insect Species" Insects 13, no. 3: 220. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030220