
Anopheles albimanus (Diptera: Culicidae) Ensemble Distribution Modeling: Applications for Malaria Elimination

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Occurrence Records
2.2. Occurrence Data Quality Control
2.3. Pseudoabsence Points
2.4. Environmental Data
2.5. Data Sources and Processing
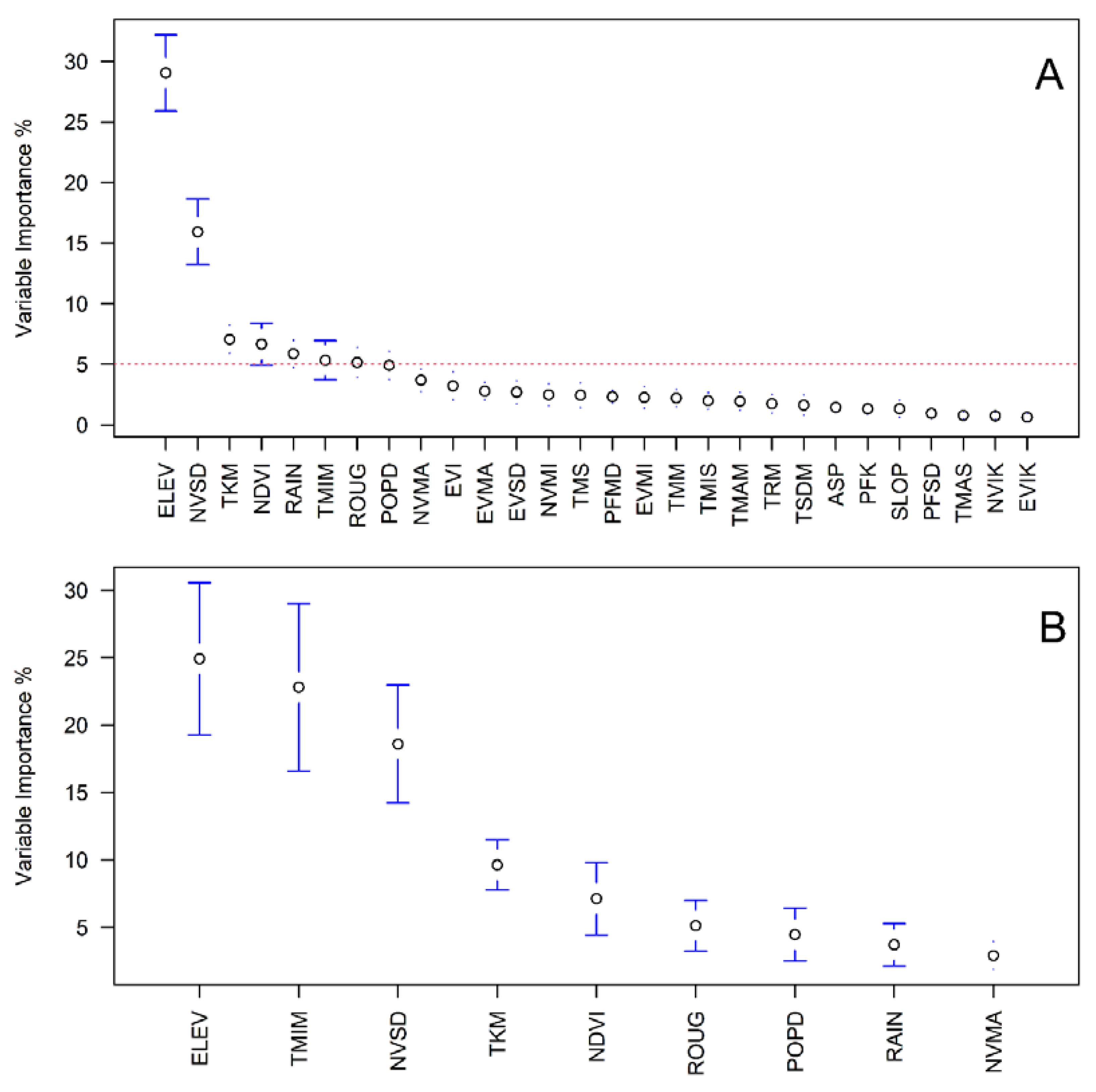
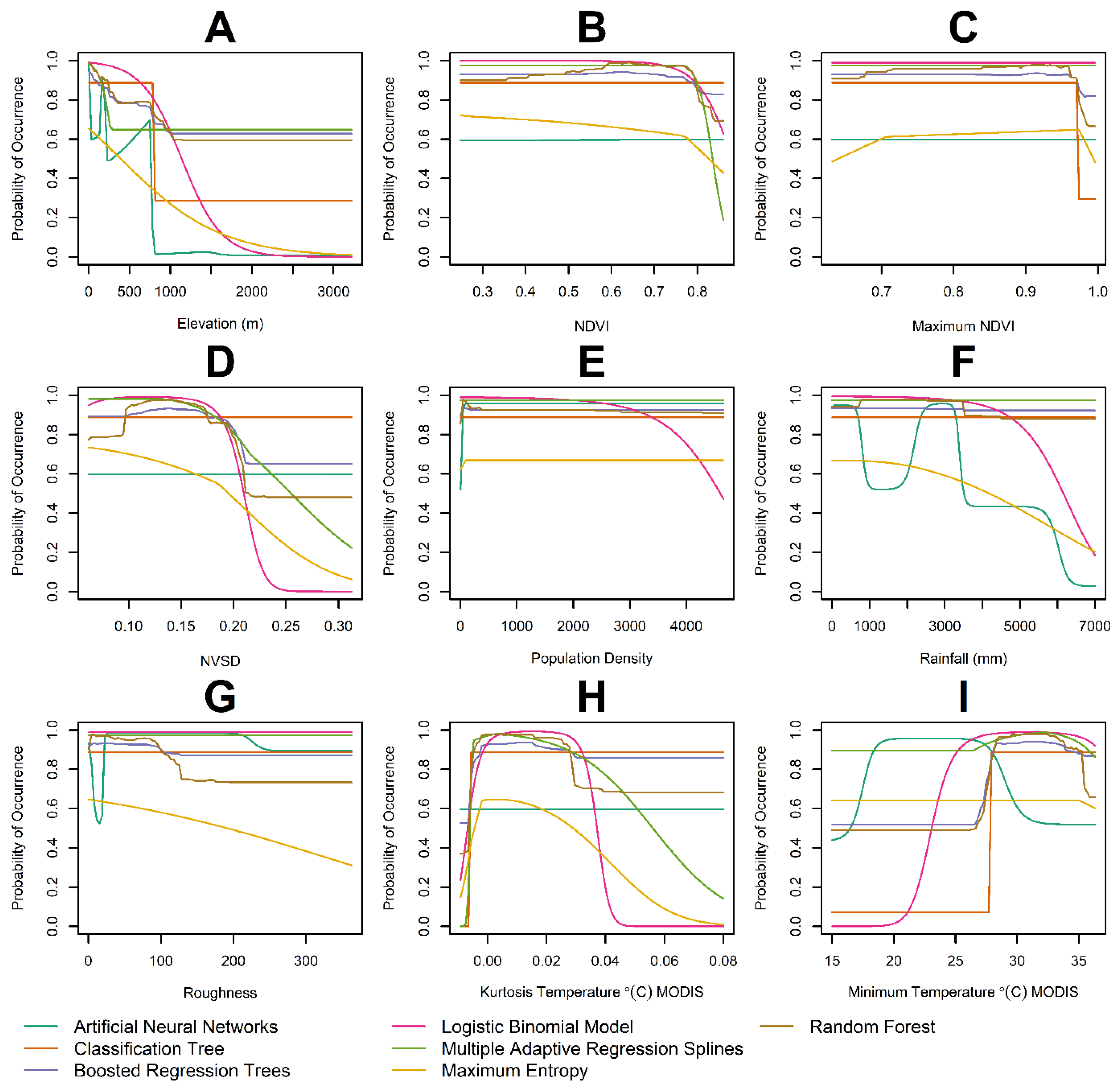
2.6. Parametric Models, Machine Learning Algorithms and Variable Selection
2.7. Applications for SDMs in the Context of Malaria Elimination
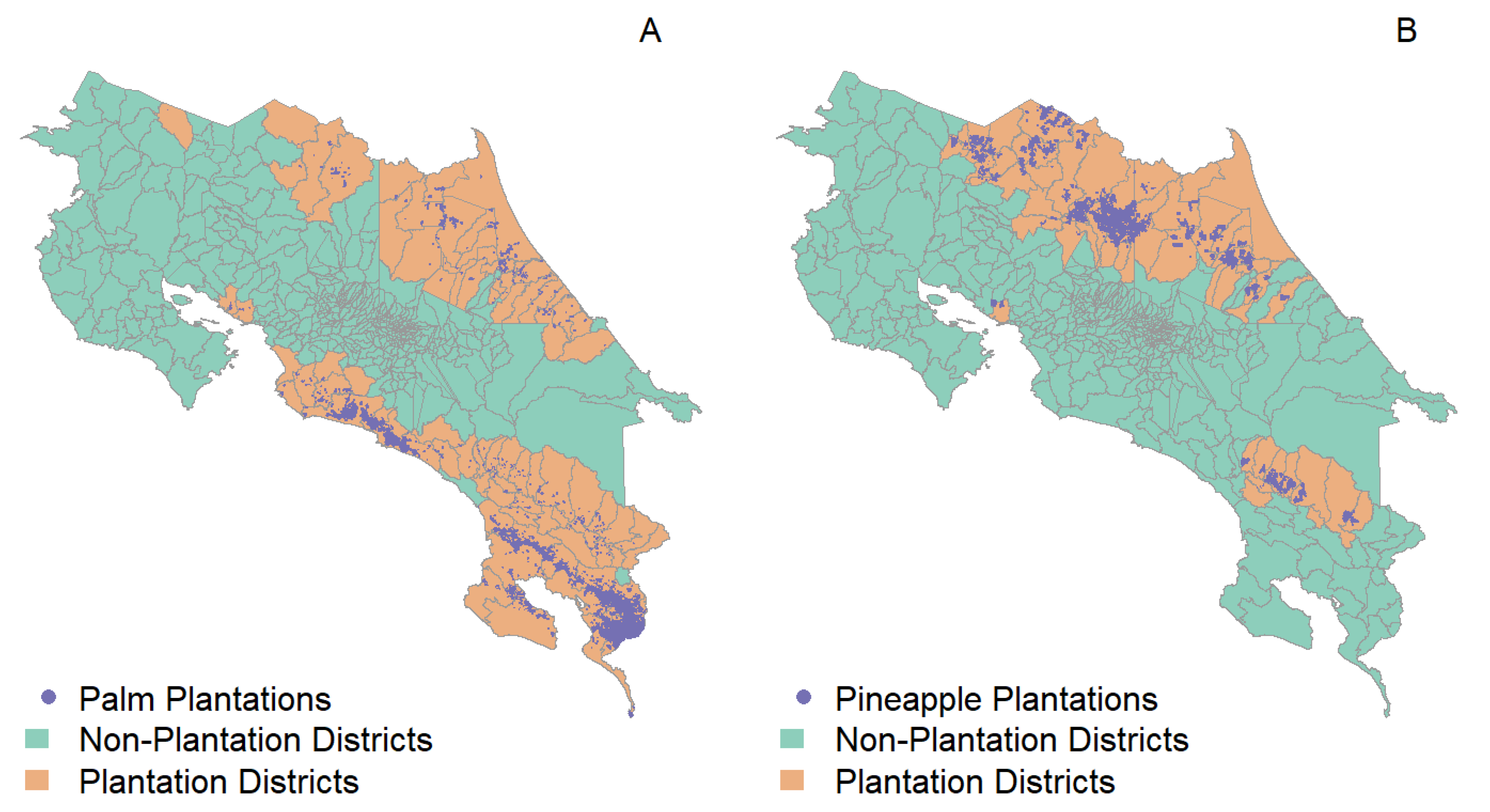
2.7.1. Background Information about Productive Landscapes in Costa Rica
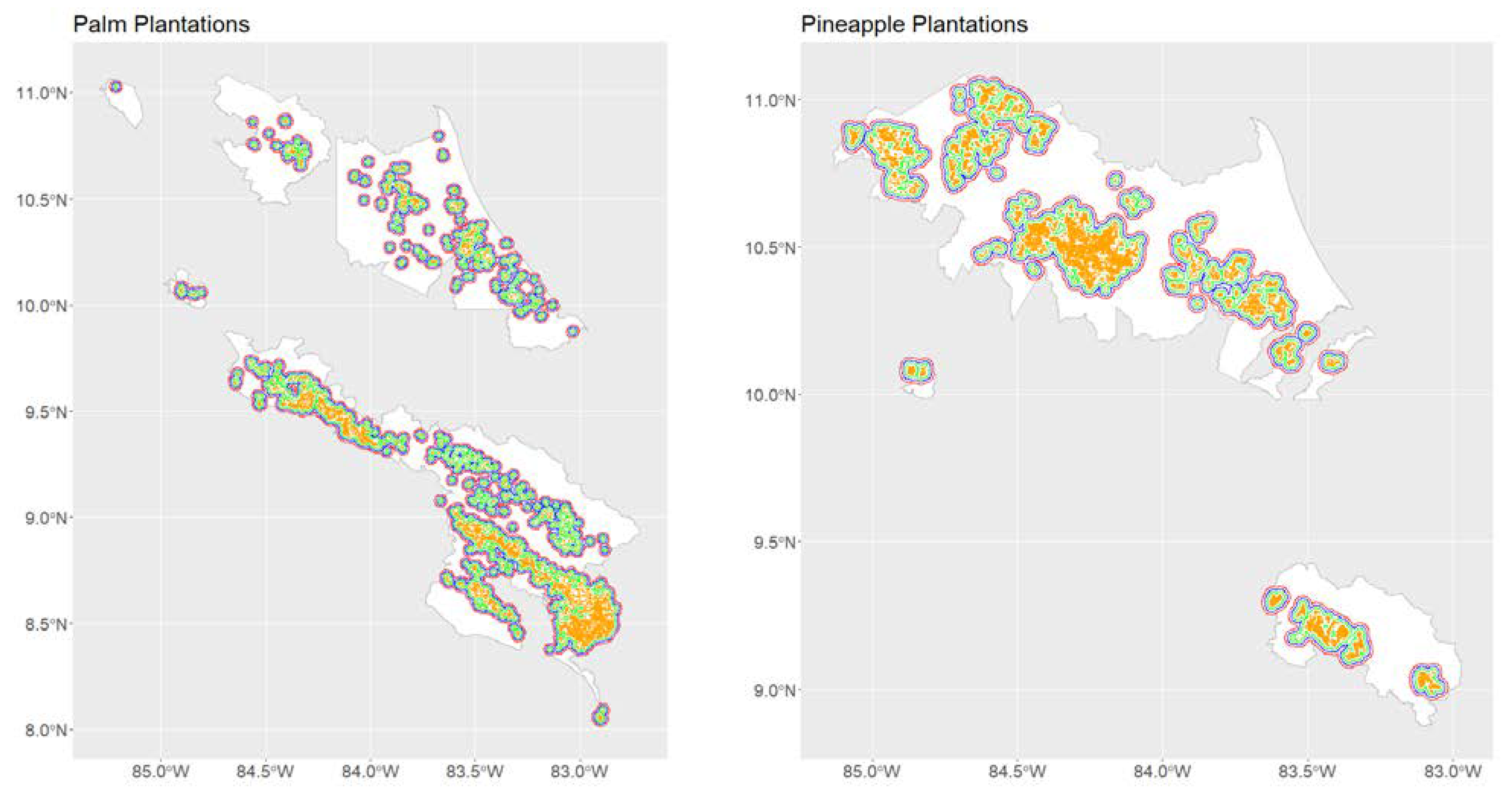
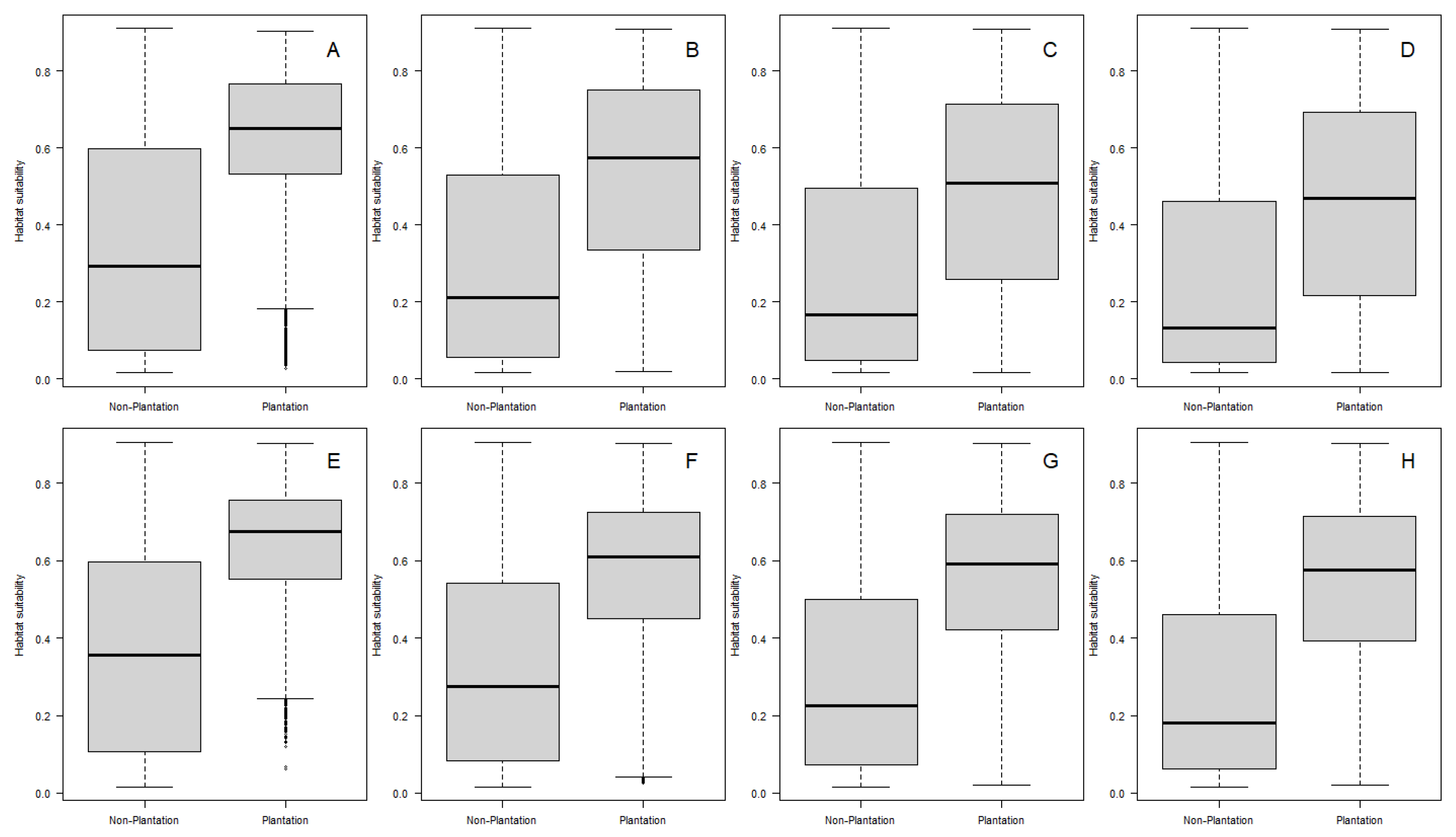
2.7.2. Anopheles albimanus in Productive Landscapes of Costa Rica
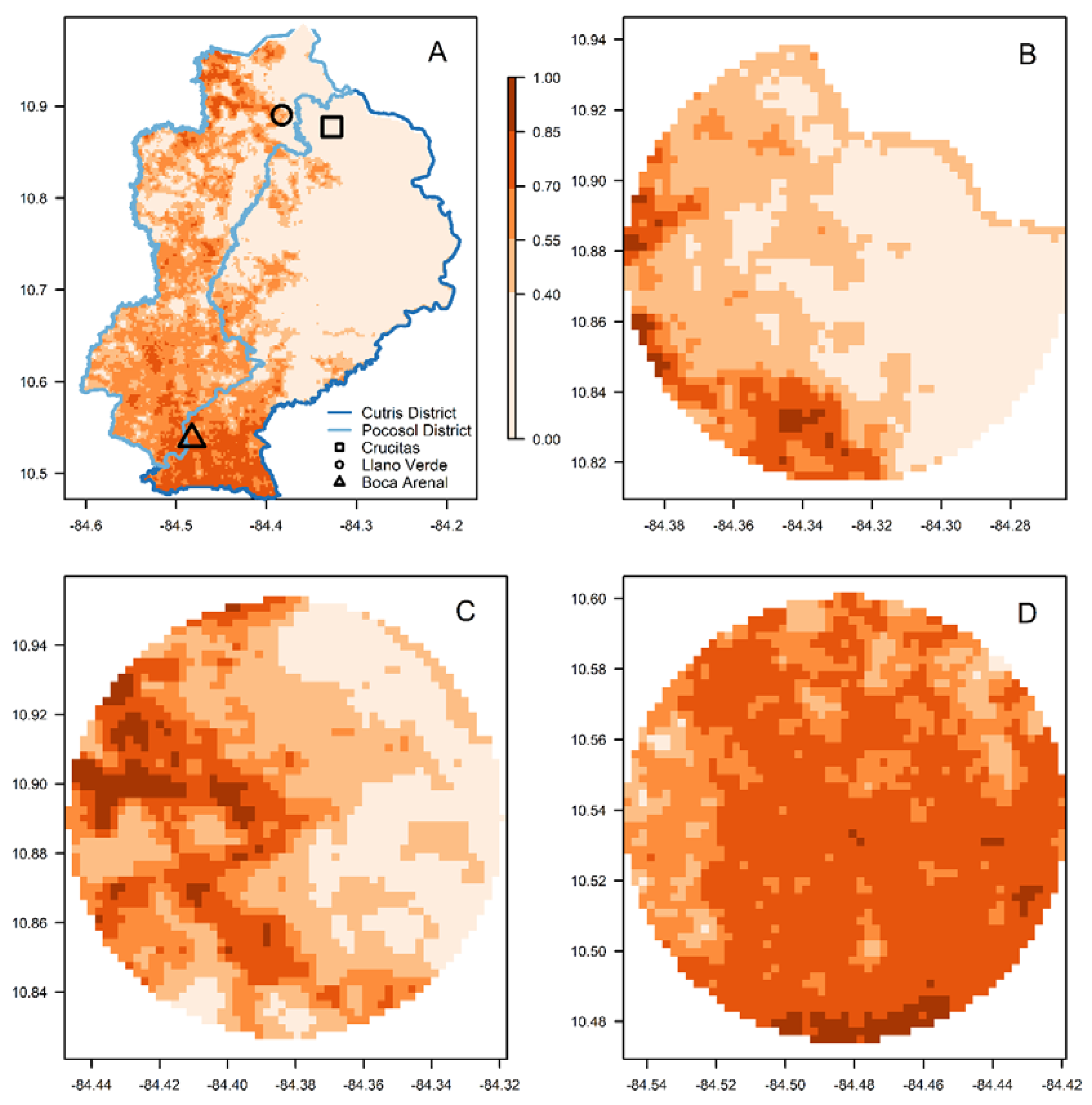
2.7.3. Anopheles albimanus in the Crucitas Outbreak of 2018–2019
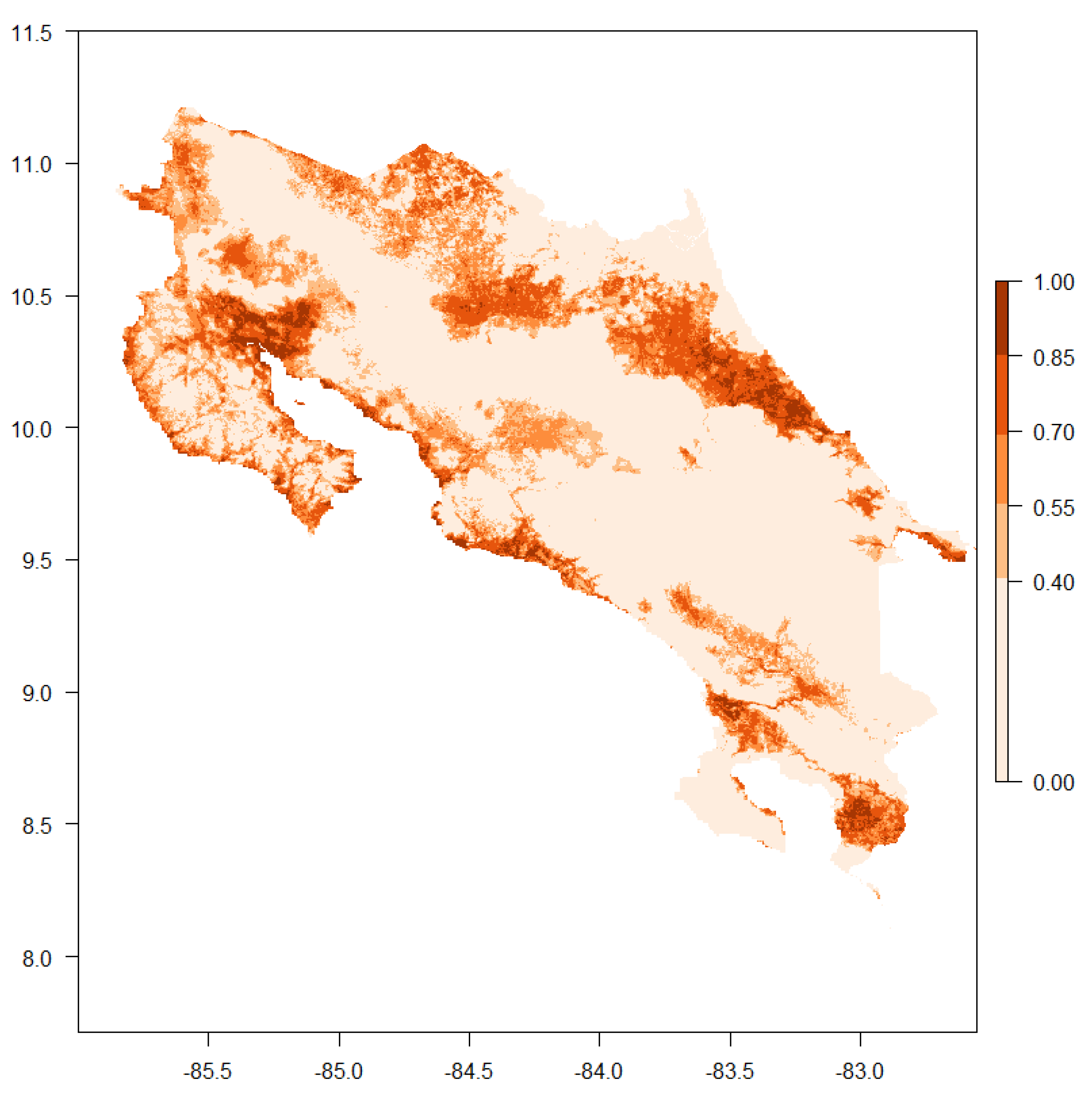
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J. Species distribution modeling and ecological niche modeling: Gettng the concepts right. Nat. Conserv. 2012, 10, 1–6. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modeling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Mendes, P.; Velazco, S.J.E.; Alves de Andrade, A.F.; De Marco, P.J. Dealing with overprediction in species distribution models: How adding distance constraints can improve model accuracy. Ecol. Model. 2020, 431, 109180. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J.; Sánchez-Cordero, V. Conservatism of ecological niches in evolutionary time. Science 1999, 285, 1265–1267. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Broennimann, O.; Guisan, A.; Wang, B.; Huang, Y.; Jiang, J. Niche conservatism in Gynandropaa frogs on the southeastern Qinghai-Tibetan Plateau. Sci. Rep. 2016, 6, 32624. [Google Scholar] [CrossRef] [Green Version]
- Mainali, K.; Hefley, T.; Ries, L.; Fagan, W.F. Matching expert range maps with species distribution model predictions. Conserv. Biol. 2020, 34, 1292–1304. [Google Scholar] [CrossRef]
- Syfert, M.M.; Joppa, L.; Smith, M.J.; Coomes, D.A.; Bachman, S.P.; Brummitt, N.A. Using species distribution models to inform IUCN Red List assessments. Biol. Conserv. 2014, 177, 174–184. [Google Scholar] [CrossRef]
- Torres, U.; Godsoe, W.; Buckley, H.L.; Parry, M.; Lusting, A.; Worner, S.P. Using niche conservatism information to prioritize hotspots of invasion by non-native freshwater invertebrates in New Zealand. Divers. Distrib. 2018, 24, 1802–1815. [Google Scholar] [CrossRef]
- Lavergne, S.; Evans, M.E.K.; Burfield, I.J.; Jiguet, F.; Thuiller, W. Are species’ responses to global change predicted by past niche evolution? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120091. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Graham, C.H. Niche Conservatism: Integrating Evolution, Ecology, and Conservation Biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Davies, T.J.; Grytnes, J.-A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Biogeogr. 2011, 38, 817–827. [Google Scholar] [CrossRef]
- Ross, R. The Prevention of Malaria, 2nd ed.; John Murray: London, UK, 1911. [Google Scholar]
- Chaves, L.F. The dialectics of malaria bednet use in sub-Saharan Africa. In The Whole Is the Truth: Essays in Honor of Richard Levins; Awerbuch, T., Clark, M.S., Taylor, P.J., Eds.; The Pumping Staiton: Arlington, MA, USA, 2018; pp. 56–80. [Google Scholar]
- Tene Fossog, B.; Ayala, D.; Acevedo, P.; Kengne, P.; Ngomo Abeso Mebuy, I.; Makanga, B.; Magnus, J.; Awono-Ambene, P.; Njiokou, F.; Pombi, M.; et al. Habitat segregation and ecological character displacement in cryptic African malaria mosquitoes. Evol. Appl. 2015, 8, 326–345. [Google Scholar] [CrossRef] [Green Version]
- White, G.B. Anopheles gambiae complex and disease transmission in Africa. Trans. R. Soc. Trop. Med. Hyg. 1974, 68, 278–301. [Google Scholar] [CrossRef]
- Derua, Y.A.; Alifrangis, M.; Hosea, K.M.; Meyrowitsch, D.W.; Magesa, S.M.; Pedersen, E.M.; Simonsen, P.E. Change in composition of the Anopheles gambiae complex and its possible implications for the transmission of malaria and lymphatic filariasis in north-eastern Tanzania. Malar. J. 2012, 11, 188. [Google Scholar] [CrossRef] [Green Version]
- Chaves, L.F.; Koenraadt, C.J.M. Climate Change and Highland Malaria: Fresh Air for a Hot Debate. Q. Rev. Biol. 2010, 85, 27–55. [Google Scholar] [CrossRef] [Green Version]
- Fuller, D.O.; Ahumada, M.L.; Quiñones, M.L.; Herrera, S.; Beier, J.C. Near-present and future distribution of Anopheles albimanus in Mesoamerica and the Caribbean Basin modeled with climate and topographci data. Int. J. Health Geogr. 2012, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Padilla, O.; Rosas, P.; Moreno, W.; Toulkeridis, T. Modeling of the ecological niches of the Anopheles spp. in Ecuador by the use of geo-informatic tools. Spat. Spatio-Temporal Epidemiol. 2017, 21, 1–11. [Google Scholar] [CrossRef]
- Carvalho, B.M.; Rangel, E.F.; Vale, M.M. Evaluation of the impacts of climate change on disease vectors through ecological niche modelling. Bull. Entomol. Res. 2017, 107, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montoya-Lerma, J.; Solarte, Y.A.; Giraldo-Calderon, G.I.; Quiñones, M.L.; Ruiz-López, F.; Wilkerson, R.C.; González, R. Malaria vector species in Colombia: A review. Mem. Inst. Oswaldo Cruz 2011, 106, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Palis, Y.; Zimmerman, R.H. Ecoregional classification of malaria vectors in the neotropics. J. Med. Entomol. 1997, 34, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Pinault, L.L.; Hunter, F.F. New highland distribution records of multiple Anopheles species in the Ecuadorian Andes. Malar. J. 2011, 10, 236. [Google Scholar] [CrossRef] [Green Version]
- Sinka, M.E.; Rubio-Palis, Y.; Manguin, S.; Patil, A.P.; Temperley, W.H.; Gething, P.W.; Van Boeckel, T.; Kabaria, C.W.; Harbach, R.E.; Hay, S.I. The dominant Anopheles vectors of human malaria in the Americas: Occurrence data, distribution maps and bionomic precis. Parasites Vectors 2010, 3, 72. [Google Scholar] [CrossRef]
- Loaiza, J.R.; Miller, M.J. Historical and contemporary forces combine to shape patterns of genetic differentiation in two species of Mesoamerican Anopheles mosquitoes. Biol. J. Linn. Soc. 2018, 126, 146–157. [Google Scholar] [CrossRef]
- Pearson, R.G.; Thuiller, W.; Araújo, M.B.; Martinez-Meyer, E.; Brotons, L.; McClean, C.; Miles, L.; Segurado, P.; Dawson, T.P.; Lees, D.C. Model-based uncertainty in species range prediction. J. Biogeogr. 2006, 33, 1704–1711. [Google Scholar] [CrossRef]
- Marin Rodriguez, R.; Chaves, L.F. Parasite Removal for Malaria Elimination in Costa Rica. Trends Parasitol. 2019, 35, 585–588. [Google Scholar] [CrossRef]
- Chaves, L.F.; Huber, J.H.; Rojas Salas, O.; Ramirez Rojas, M.; Romero, L.M.; Gutierrez Alvarado, J.M.; Perkins, T.A.; Prado, M.; Rodriguez, R.M. Malaria Elimination in Costa Rica: Changes in Treatment and Mass Drug Administration. Microorganisms 2020, 8, 984. [Google Scholar] [CrossRef]
- Chaves, L.F.; Ramírez Rojas, M.; Delgado Jiménez, S.; Prado, M.; Marín Rodríguez, R. Housing quality improvement is associated with malaria transmission reduction in Costa Rica. Socio-Econ. Plan. Sci. 2021, 74, 100951. [Google Scholar] [CrossRef]
- Villalobos, J.A. El Oro Que Contemplan Los Gusanos, Que Lo Disfruten Los Humanos”. Crucitas y La Disputa Por El Desarrollo En Costa Rica. Anu. De Estud. Centroam. 2016, 42, 133–157. [Google Scholar] [CrossRef] [Green Version]
- Rigg, C.A.; Hurtado, L.A.; Calzada, J.E.; Chaves, L.F. Malaria infection rates in Anopheles albimanus (Diptera: Culicidae) at Ipetí-Guna, a village within a region targeted for malaria elimination in Panamá. Infect. Genet. Evol. 2019, 69, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, L.A.; Rigg, C.A.; Calzada, J.E.; Dutary, S.; Bernal, D.; Koo, S.I.; Chaves, L.F. Population dynamics of Anopheles albimanus (Diptera: Culicidae) at Ipetí-Guna, a willage in a region targeted for malaria elimination in Panamá. Insects 2018, 9, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grupo Técnico Nacional de Enfermedades Vectoriales. Plan de Eliminación de la Malaria en Costa Rica, 2015–2020; Ministerio de Salud de Costa Rica: San José, Costa Rica, 2015; p. 16. [Google Scholar]
- Grupo Técnico Nacional de Enfermedades Vectoriales. Norma de Malaria; Llorca, F., Ed.; Ministerio de Salud de Costa Rica: San José, Costa Rica, 2016; p. 60. [Google Scholar]
- Chaves, L.F.; Ramírez Rojas, M.; Prado, M.; Garcés, J.L.; Salas Peraza, D.; Marín Rodríguez, R. Health policy impacts on malaria transmission in Costa Rica. Parasitology 2020, 147, 999–1007. [Google Scholar] [CrossRef]
- Chaves, L.F.; Valerín Cordero, J.A.; Delgado, G.; Aguilar-Avendaño, C.; Maynes, E.; Gutiérrez Alvarado, J.M.; Rojas, M.R.; Romero, L.M.; Rodríguez, R.M. Modeling the association between Aedes aegypti ovitrap egg counts, multi-scale remotely sensed environmental data and arboviral cases at Puntarenas, Costa Rica (2017–2018). Curr. Res. Parasitol. Vector-Borne Dis. 2021, 1, 100014. [Google Scholar] [CrossRef]
- Loaiza, J.R.; Scott, M.E.; Bermingham, E.; Sanjur, O.I.; Rovira, J.R.; Dutari, L.C.; Linton, Y.-M.; Bickersmith, S.; Conn, J.E. Novel genetic diversity within Anopheles punctimacula s.l.: Phylogenetic discrepancy between the Barcode cytochrome c oxidase I (COI) gene and the rDNA second internal transcribed spacer (ITS2). Acta Trop. 2013, 128, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Loaiza, J.R.; Scott, M.E.; Bermingham, E.; Sanjur, O.I.; Wilkerson, R.; Rovira, J.; Gutiérrez, L.A.; Correa, M.M.; Grijalva, M.J.; Birnberg, L.; et al. Late Pleistocene environmental changes lead to unstable demography and population divergence of Anopheles albimanus in the northern Neotropics. Mol. Phylogenet. Evol. 2010, 57, 1341–1346. [Google Scholar] [CrossRef] [Green Version]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radisavljevic, B.V.; Anderson, R.P.; Bjornson, R.; Weston, S. spThin: Functions for Spatial Thinning of Species Occurrence Records for Use in Ecological Models; R Package Version 0.1.0; 2014. [Google Scholar]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Instituto Geográfico Nacional. IGN Cartografía 1:5mil: Urbano 1:5mil: Sistema Nacional de Información Territorial. 2014. Available online: https://geos.snitcr.go.cr/be/IGN_5/wfs? (accessed on 10 August 2020).
- USNPS. Data Sources & Accuracy for National Park Service Maps. 2020. Available online: https://www.nps.gov/hfc/carto/data-sources.cfm (accessed on 31 August 2020).
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where, and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Monaghan, A.J.; Eisen, R.J.; Eisen, L.; Mcallister, J.; Savage, H.M.; Mutebi, J.-P.; Johansson, M.A. Consensus and uncertainty in the geographic range of Aedes aegypti and Aedes albopictus in the contiguous United States: Multi-model assessment and synthesis. PLoS Comput. Biol. 2019, 15, e1007369. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. Package Biomod2; 2016. [Google Scholar]
- Didan, K. MOD13Q1: MODIS/Terra Vegetation Indices 16-Day L3 Global 250 m SIN Grid V006 2015. Available online: https://lpdaac.usgs.gov/products/mod13q1v006/ (accessed on 8 October 2018).
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Hook, S.; Hulley, G. MOD11A1 MODIS/Terra Land Surface Temperature/Emissivity Daily L3 Global 1km SIN Grid V006. 2015, Distributed by NASA EOSDIS Land Processes DAAC. Available online: https://lpdaac.usgs.gov/products/mod11a1v006/ (accessed on 29 August 2020).
- Shimada, M.; Itoh, T.; Motooka, T.; Watanabe, M.; Shiraishi, T.; Thapa, R.; Lucas, R. New global forest/non-forest maps from ALOS PALSAR data (2007–2010). Remote Sens. Environ. 2014, 155, 13–31. [Google Scholar] [CrossRef]
- Center for International Earth Science Information Network—CIESIN—Columbia University. Gridded Population of the World, Version 4 (GPWv4): Population Density Adjusted to Match 2015 Revision UN WPP Country Totals, Revision 11 Palisades, NY: NASA Socioeconomic Data and Applications Center (SEDAC). 2018. Available online: https://sedac.ciesin.columbia.edu/data/set/gpw-v4-population-density-adjusted-to-2015-unwpp-country-totals-rev11 (accessed on 10 August 2021).
- NASA JPL. NASADEM Merged DEM Global 1 arc Second V001. Distributed by NASA EOSDIS Land Processes DAAC. 2020. Available online: https://lpdaac.usgs.gov/products/nasadem_hgtv001/ (accessed on 10 August 2021).
- Google. Earth Engine Data Catalog. 2020. Available online: https://developers.google.com/earth-engine/datasets (accessed on 1 November 2020).
- Brunsdon, C.; Comber, L. An introduction to R for Spatial Analysis and Mapping; Sage Publications LTD: London, UK, 2015; p. 343. [Google Scholar]
- Parker, J.A.; Kenyon, R.V.; Troxel, D.E. Comparison of Interpolating Methods for Image Resampling. IEEE Trans. Med. Imaging 1983, 2, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.F.; Moji, K. Density Dependence, Landscape, and Weather Impacts on Aquatic Aedes japonicus japonicus (Diptera: Culicidae) Abundance Along an Urban Altitudinal Gradient. J. Med. Entomol. 2018, 55, 329–341. [Google Scholar] [CrossRef]
- Wilson, M.F.; O’Connell, B.; Brown, C.; Guinan, J.C.; Grehan, A.J. Multiscale terrain analysis of multibeam bathymetry data for habitat mapping on the continental slope. Mar. Geod. 2007, 30, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Instituto Meteorologico Nacional. Precipitación Anual 1960–2013: Sistema Nacional de Información Territorial. 2014. Available online: http://ceniga.go.cr/geoserver/IMN/wfs? (accessed on 10 August 2020).
- Instituto Meteorologico Nacional. Temperatura Media Costa Rica 1960_2013: Sistema Nacional de Información Territorial. 2014. Available online: http://ceniga.go.cr/geoserver/IMN/wfs? (accessed on 10 August 2021).
- Instituto Meteorologico Nacional. Temperatura Mínima Costa Rica 1960_2013: Sistema Nacional de Información Territorial. 2014. Available online: http://ceniga.go.cr/geoserver/IMN/wfs? (accessed on 10 August 2021).
- Instituto Meteorologico Nacional. Temperatura Máxima Costa Rica 1960_2013: Sistema Nacional de Información Territorial. 2014. Available online: http://ceniga.go.cr/geoserver/IMN/wfs? (accessed on 10 August 2021).
- Guisan, A.; Edwards, T.C.; Hastie, T. Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 2002, 157, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Kuhn, M.; Johnson, K. Applied Predictive Modeling; Springer: New York, NY, USA, 2013; p. 600. [Google Scholar]
- Olden, J.D.; Lawler, J.J.; LeRoy-Poff, N. Machine Learning Methods Without Tears: A Primer for Ecologists. Q. Rev. Biol. 2008, 83, 171–193. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.O.; Chaves, L.F.; Hamer, G.L.; Sun, T.; Brown, W.M.; Walker, E.D.; Haramis, L.; Goldberg, T.L.; Kitron, U.D. Local impact of temperature and precipitation on West Nile virus infection in Culex species mosquitoes in northeast Illinois, USA. Parasites Vectors 2010, 3, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leathwick, J.; Elith, J.; Hastie, T. Comparative performance of generalized additive models and multivariate adaptive regression splines for statistical modelling of species distributions. Ecol. Model. 2006, 199, 188–196. [Google Scholar] [CrossRef]
- Elith, J.; Ferrier, S.; Huettmann, F.; Leathwick, J. The evaluation strip: A new and robust method for plotting predicted responses from species distribution models. Ecol. Model. 2005, 186, 280–289. [Google Scholar] [CrossRef]
- Thuiller, W. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Torres, R.; Acosta, Á.; Chinchilla, C. Proyecto comercial de compostaje de los desechos agroindustriales de la palma aceitera. Rev. Palmas 2004, 25, 377–387. [Google Scholar]
- Escobar, E.; Peralta, F. La industria de la palma aceitera en Costa Rica. ASD Oil Palm Pap. 2007, 31, 21–24. [Google Scholar]
- PRIAS. Monitoreo de Cambio de Uso en Paisajes Productivos—Palma. 2021. Available online: https://monitoreo.prias.cenat.ac.cr/geoserver/MonitoreoPalma/wfs? (accessed on 14 December 2021).
- PRIAS. Monitoreo de Cambio de Uso en Paisajes Productivos—Piña. 2021. Available online: https://monitoreo.prias.cenat.ac.cr/geoserver/MonitoreoPina/wfs? (accessed on 14 December 2021).
- León Araya, A. Desarrollo Geográfico Desigual en Costa Rica. El Ajuste Estructural Visto Desde la Región Huétar Norte (1985–2005); Editorial Universidad de Costa Rica: San Josè, Costa Rica, 2015; p. 204. [Google Scholar]
- CANAPEP. Estadísticas. 2021. Available online: https://canapep.com/estadisticas/ (accessed on 14 December 2021).
- Chang, M.; Hii, J.; Buttner, P.; Mansoor, F. Changes in abundance and behaviour of vector mosquitoes induced by land use during the development of an oil palm plantation in Sarawak. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 382–386. [Google Scholar] [CrossRef]
- Young, K.I.; Buenemann, M.; Vasilakis, N.; Perera, D.; Hanley, K.A. Shifts in mosquito diversity and abundance along a gradient from oil palm plantations to conterminous forests in Borneo. Ecosphere 2021, 12, e03463. [Google Scholar] [CrossRef]
- Hobbs, J.; Lowe, R.; Schreck, C. Studies of flight range and survival of Anopheles albimanus Wiedemann in El Salvador. I. Dispersal and survival during the dry season. Mosq. News 1974, 34, 389–393. [Google Scholar]
- Lowe, R.E.; Schreck, C.; Hobbs, J.; Dame, D.; Lofgren, C. Studies on flight range and survival of Anopheles albimanus Wiedemann in El Salvador. II. Comparisons of release methods with sterile and normal adults in wet and dry seasons. Mosq. News 1975, 35, 160–168. [Google Scholar]
- Welch, B.L. The generalization of Student’s problem when several different population variances are involved. Biometrika 1947, 34, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Machado-Allison, C.; Bown, D.N.; Nelson, M. Biología y ecología de Anopheles albimanus Wiedemann en Centroamérica. Bol. Oficina Sanit. Panam. 1996, 121, 189–220. [Google Scholar]
- Grieco, J.P.; Johnson, S.; Achee, N.L.; Masuoka, P.; Pope, K.; Rejmankova, E.; Vanzie, E.; Andre, R.; Roberts, D. Distribution of Anopheles albimanus, Anopheles vestitipennis, and Anopheles crucians associated with land use in northern Belize. J. Med. Entomol. 2006, 43, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M. Algunas observaciones sobre los hábitos de Anopheles (N.) albimanus y Anopheles (A.) punctimacula adultos, en la localidad de Matapalo (Puntarenas) Costa Rica. Rev. De Biol. Trop. 1961, 9, 153–170. [Google Scholar]
- Vargas, V.M.; Vargas, C.J.V. Male and mosquito larvae survey at the Arenal-Tempisque irrigation project, Guanacaste, Costa Rica. Rev. Biol. Trop. 2003, 51, 759–762. [Google Scholar]
- Rejmankova, E.; Roberts, D.; Harbach, R.; Pecor, J.; Peyton, E.; Manguin, S.; Krieg, R.; Polanco, J.; Legters, L. Environmental and regional determinants of Anopheles (Diptera: Culicidae) larval distribution in Belize, Central America. Environ. Entomol. 1993, 22, 978–992. [Google Scholar] [CrossRef] [Green Version]
- Rejmankova, E.; Savage, H.; Rejmanek, M.; Arredondo-Jimenez, J.; Roberts, D. Multivariate analysis of relationships between habitats, environmental factors and occurrence of anopheline mosquito larvae Anopheles albimanus and A. pseudopunctipennis in southern Chiapas, Mexico. J. Appl. Ecol. 1991, 28, 827–841. [Google Scholar] [CrossRef]
- Rejmankova, E.; Savage, H.; Rodriguez, M.; Roberts, D.; Rejmanek, M. Aquatic vegetation as a basis for classification of Anopheles albimanus Weideman (Diptera: Culicidae) larval habitats. Environ. Entomol. 1992, 21, 598–603. [Google Scholar] [CrossRef]
- Rodriguez, A.D.; Rodriguez, M.H.; Hernandez, J.E.; Dister, S.W.; Beck, L.R.; Rejmankova, E.; Roberts, D.R. Landscape surrounding human settlements and Anopheles albimanus (Diptera: Culicidae) abundance in Southern Chiapas, Mexico. J. Med. Entomol. 1996, 33, 39–48. [Google Scholar] [CrossRef]
- Hernandez, J.E.; Epstein, L.D.; Rodriguez, M.H.; Rodriguez, A.D.; Rejmankova, E.; Roberts, D.R. Use of generalized regression tree models to characterize vegetation favoring Anopheles albimanus breeding. J. Am. Mosq. Control Assoc. 1997, 13, 28–34. [Google Scholar]
- Chaves, L.F.; Friberg, M.D. Aedes albopictus and Aedes flavopictus (Diptera: Culicidae) pre-imaginal abundance patterns are associated with different environmental factors along an altitudinal gradient. Curr. Res. Insect Sci. 2021, 1, 100001. [Google Scholar] [CrossRef]
- Chaves, L.F.; Morrison, A.C.; Kitron, U.D.; Scott, T.W. Nonlinear impacts of climatic variability on the density-dependent regulation of an insect vector of disease. Glob. Change Biol. 2012, 18, 457–468. [Google Scholar] [CrossRef]
- Chaves, L.F. Climate change and the biology of insect vectors of human pathogens. In Invertebrates and Global Climate Change; Johnson, S., Jones, H., Eds.; Wiley: Chichester, UK, 2017; pp. 126–147. [Google Scholar]
- Wallace, R.; Chaves, L.F.; Bergmann, L.; Ayres Lopes, C.f.J.; Hogerwerf, L.; Kock, R.; Wallace, R.G. Clear-Cutting Disease Control: Capital-Led Deforestation, Public Health Austerity, and Vector-Borne Infection; Springer: New York, NY, USA, 2018; 72p. [Google Scholar]
- Wallace, R.G.; Bergmann, L.; Kock, R.; Gilbert, M.; Hogerwerf, L.; Wallace, R.; Holmberg, M. The dawn of Structural One Health: A new science tracking disease emergence along circuits of capital. Soc. Sci. Med. 2015, 129, 68–77. [Google Scholar] [CrossRef]
- Palmer, S. From Popular Medicine to Medical Populism: Doctors, Healers, and Public Power in Costa Rica, 1800–1940; Duke University Press Books: Durham, NC, USA, 2003; p. 352. [Google Scholar]
- Fallas, C.L. Mamita Yunai; Edit: San José, Costa Rica, 2013. [Google Scholar]
- Gutiérrez, J. Puerto Limón; Edit: San José, Costa Rica, 1977. [Google Scholar]
- Aliano, D. Curing the Ills of Central America: The United Fruit Company´s Medical Department and Corporate America´s Mission to Civilize (1900–1940). Estud. Interdiscip. Am. Lat. Y Caribe 2006, 17, 35–60. [Google Scholar]
- Wolford, W. The Plantationocene: A Lusotropical Contribution to the Theory. Ann. Am. Assoc. Geogr. 2021, 111, 1622–1639. [Google Scholar] [CrossRef]
- Pinault, L.L.; Hunter, F.F. Characterization of Larval Habitats of Anopheles albimanus, Anopheles pseudopunctipennis, Anopheles punctimacula, and Anopheles oswaldoi s.l. Populations in Lowland and Highland Ecuador. J. Vector Ecol. 2012, 37, 124–136. [Google Scholar] [CrossRef]
- Loyola, E.G.; Arredondo, J.I.; Rodríguez, M.H.; Brown, D.N.; Vaca-Marin, M.A. Anopheles vestitipennis, the probable vector of Plasmodium vivax in the Lacandon forest of Chiapas, México. Trans. R. Soc. Trop. Med. Hyg. 1991, 85, 171–174. [Google Scholar] [CrossRef]
- Hiwat, H.; Bretas, G. Ecology of Anopheles darlingi Root with respect to vector importance: A review. Parasites Vectors 2011, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Loaiza, J.; Scott, M.; Bermingham, E.; Rovira, J.; Sanjur, O.; Conn, J.E. Anopheles darlingi (Diptera: Culicidae) in Panama. Am. J. Trop. Med. Hyg. 2009, 81, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Ramaswami, R.; Bayer, R.; Galea, S. Precision medicine from a public health perspective. Annu. Rev. Public Health 2018, 39, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Galea, S.; Abdalla, S.M. Precision medicine approaches and the health of populations: Study design concerns and considerations. Persp. Biol. Med. 2018, 61, 527–536. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Covariate | Raster Original Spatial Resolution (Covariate Units) | Frequency (Period Sampled) | Derived Layers (Abbreviation) |
|---|---|---|---|
| MODIS—Enhanced Vegetation Index (EVI) | 250 m (Adimensional Ratio) | 16 days (2000-02-24 to 2019-12-31) | Standard Deviation, SD (EVSD) Kurtosis (EVIK) Maximum (EVMA) Minimum (EVMI) Median (EVI) |
| MODIS—Normalized Difference Vegetation Index (NDVI) | 250 m (Adimensional Ratio) | 16 days (2000-02-24 to 2019-12-31) | SD (NVSD) Kurtosis (NVIK) Maximum (NVMA) Minimum (NVMI) Median (NDVI) |
| MODIS—Land Surface Temperature | 1000 m (° Kelvin) | Daily (2000-02-24 to 2019-12-31) | SD (TSDM) Kurtosis (TKM) Maximum (TMAM) Minimum (TMIM) Range (TRM) Median (TMM) |
| PALSAR—Forest/Non-Forest | 25 m (1 = Forest, 2 = Non-forest, 3 = Water) | Annual (2007–2019) | Mode (PFC) SD (PFSD) Kurtosis (PFK) |
| NASA—Digital Elevation Model | 30 m (Meters Above Sea Level) | 2000 s * | Elevation (ELEV) Aspect (ASP) Roughness (ROUG) Slope (SLOP) |
| GPWv4—Population Density | 1000 m (Population Density) | 2015 | Population Density (POPD) |
| INM—Rainfall | 1:5000 (mm) ** | Annual average based on daily records (1963-01-01 2013-12-31) | Rainfall (RAIN) |
| INM—Temperature | 1:5000 (°C) ** | Annual average based on daily records (1963-01-01 2013-12-31) | Mean (TMS) Minimum (TMIS) Maximum (TMAS) |
| Algorithm | AUC | TSS |
|---|---|---|
| L-GLM | 0.91 ± 0.05 | 0.75 ± 0.11 |
| MARS | 0.89 ± 0.04 | 0.68 ± 0.09 |
| CAT | 0.79 ± 0.07 | 0.58 ± 0.13 |
| RF | 0.92 ± 0.04 | 0.76 ± 0.10 |
| GBM | 0.91 ± 0.05 | 0.72 ± 0.10 |
| ANN | 0.85 ± 0.07 | 0.63 ± 0.12 |
| MAXENT | 0.92 ± 0.04 | 0.76 ± 0.10 |
| Location | Spatial Buffer Radius (HS Mean ± S.D.) | ||
|---|---|---|---|
| 3 km | 5 km | 7 km | |
| Crucitas | 0.10 ± 0.04 | 0.12 ± 0.09 | 0.17 ± 0.15 |
| Llano Verde | 0.33 ± 0.18 | 0.33 ± 0.19 | 0.31 ± 0.19 |
| Boca Arenal | 0.70 ± 0.05 | 0.69 ± 0.06 | 0.67 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rhodes, C.G.; Loaiza, J.R.; Romero, L.M.; Gutiérrez Alvarado, J.M.; Delgado, G.; Rojas Salas, O.; Ramírez Rojas, M.; Aguilar-Avendaño, C.; Maynes, E.; Valerín Cordero, J.A.; et al. Anopheles albimanus (Diptera: Culicidae) Ensemble Distribution Modeling: Applications for Malaria Elimination. Insects 2022, 13, 221. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030221
Rhodes CG, Loaiza JR, Romero LM, Gutiérrez Alvarado JM, Delgado G, Rojas Salas O, Ramírez Rojas M, Aguilar-Avendaño C, Maynes E, Valerín Cordero JA, et al. Anopheles albimanus (Diptera: Culicidae) Ensemble Distribution Modeling: Applications for Malaria Elimination. Insects. 2022; 13(3):221. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030221
Chicago/Turabian StyleRhodes, Charlotte G., Jose R. Loaiza, Luis Mario Romero, José Manuel Gutiérrez Alvarado, Gabriela Delgado, Obdulio Rojas Salas, Melissa Ramírez Rojas, Carlos Aguilar-Avendaño, Ezequías Maynes, José A. Valerín Cordero, and et al. 2022. "Anopheles albimanus (Diptera: Culicidae) Ensemble Distribution Modeling: Applications for Malaria Elimination" Insects 13, no. 3: 221. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13030221