
Does Bacillus thuringiensis Affect the Stress and Immune Responses of Rhynchophorus ferrugineus Larvae, Females, and Males in the Same Way?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing
2.2. Insect Bioassay: Lethal Concentration and Lethal Time
2.3. Bleeding Procedure
2.4. Bacillus Thuringiensis Effects on Hemocyte Number and Type
2.5. Protein Extraction
2.5.1. Hemocyte Lysate Supernatant
2.5.2. Brain Lysate Supernatant
2.6. Hsp 70 Estimation
2.7. Statistical Analyses
3. Results
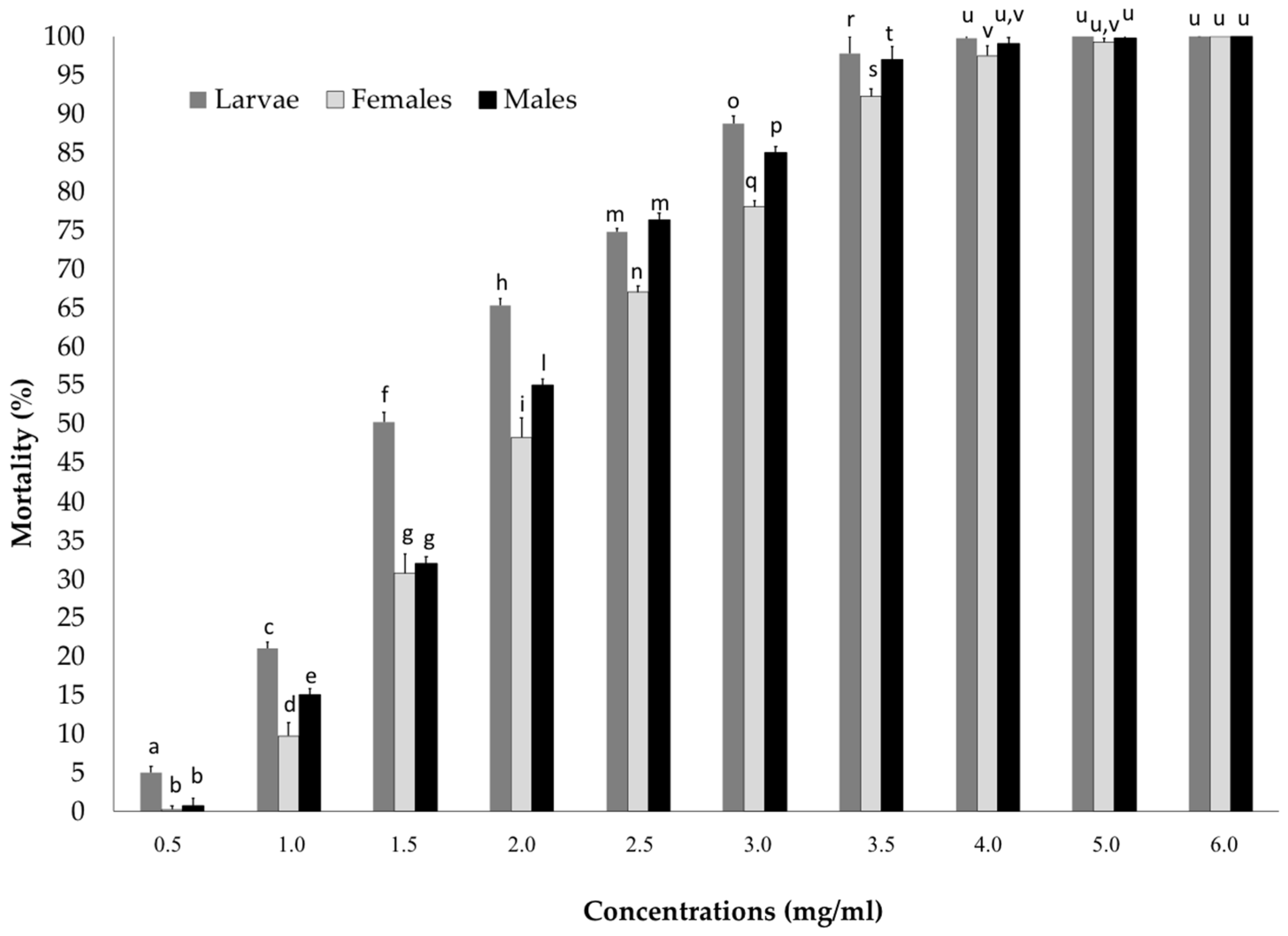
3.1. Laboratory Bioassay: Lethal Concentration and Lethal Time
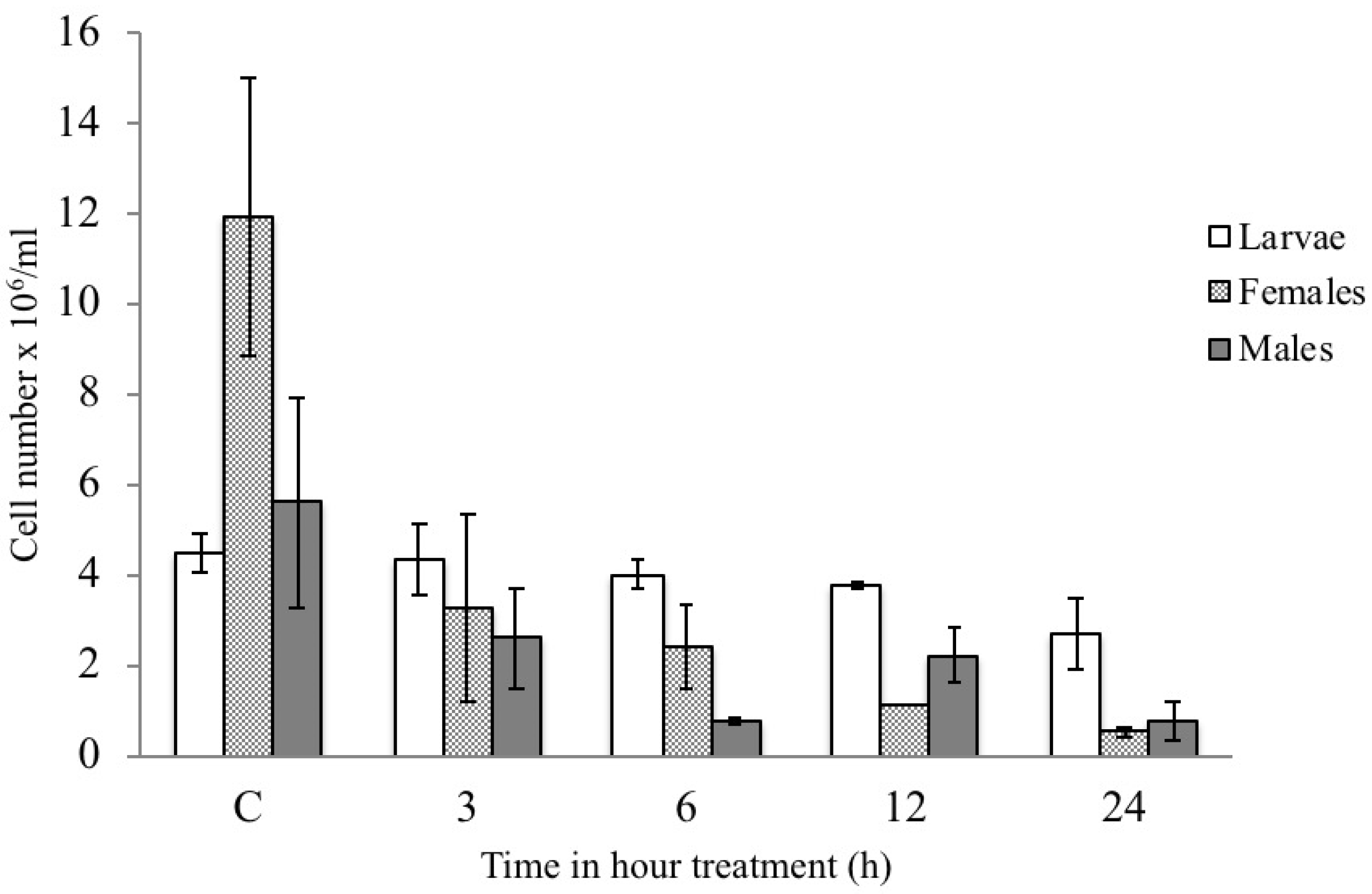
3.2. Effects of Bt on Hemocytes
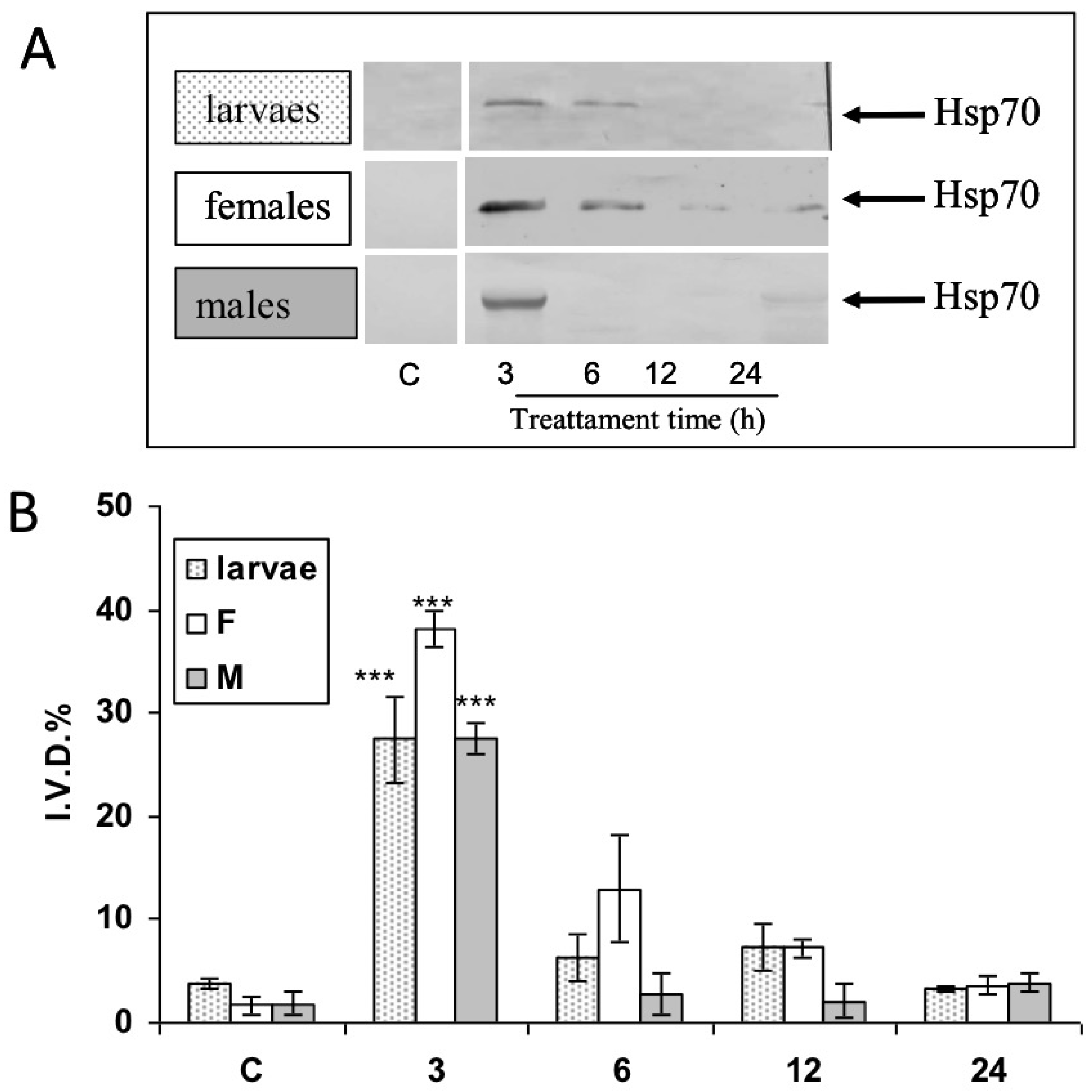
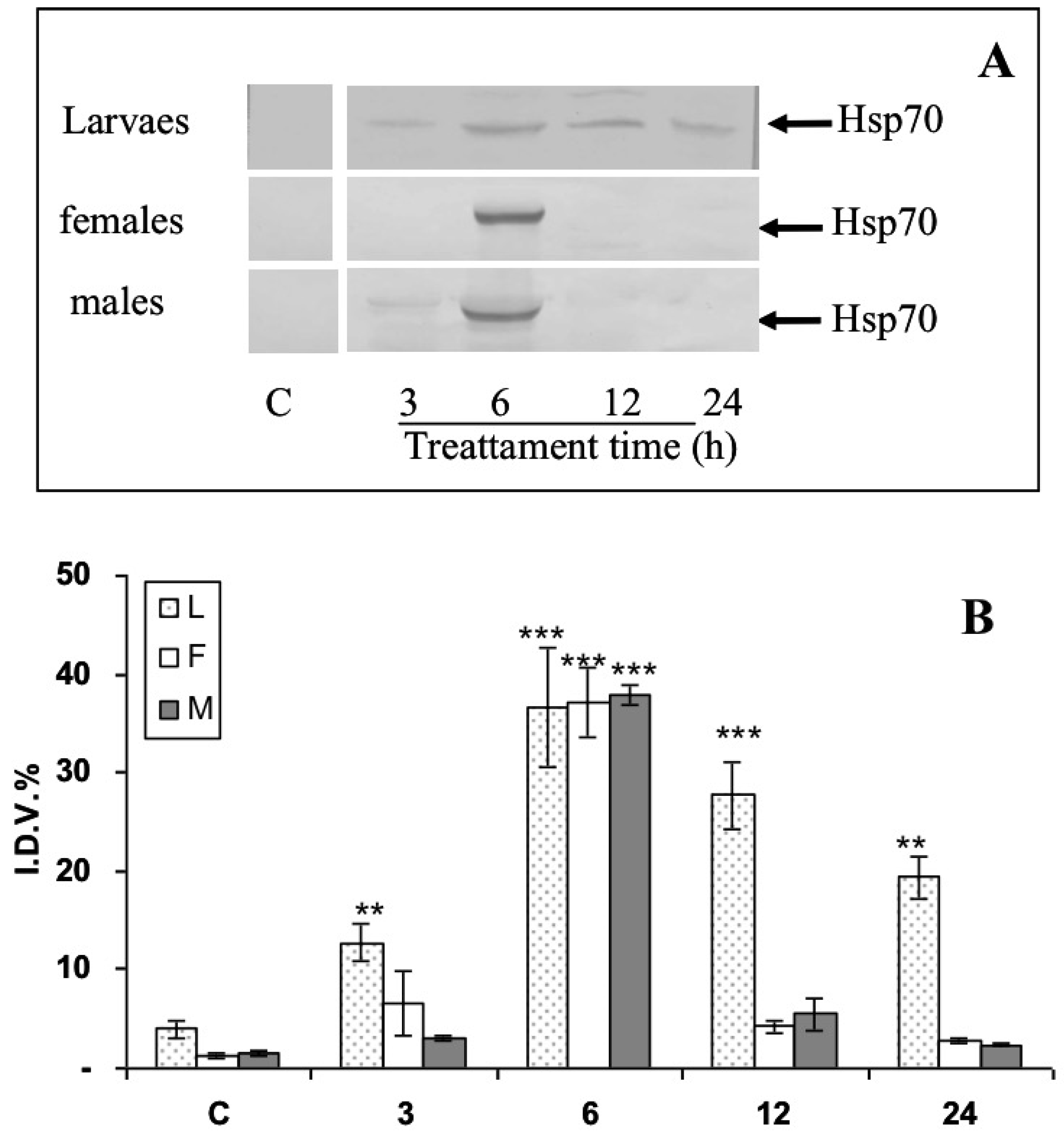
3.3. Modulation of Hsp70 from RPW Larvae, Females, and Males in Hemocytes and the Brain after Bt Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Feng, M.; Xia, J.; Fei, S.; Peng, R.; Wang, X.; Zhou, Y.Z.; Wang, P.; Swevers, L.; Sun, J. Identification of silkworm hemocyte subsets and analysis of their response to baculovirus infection based on single-cell RNA sequencing. Front. Immunol. 2021, 12, 645359. [Google Scholar] [CrossRef] [PubMed]
- Manachini, B.R.I.; Bue, P.L.; Peri, E.; Colazza, S. Potential effects of Bacillus thuringiensis against adults and older larvae of Rhynchophorus ferrugineus. IOBC/WPRS Bull. 2009, 45, 239–242. [Google Scholar]
- Manachini, B.; Arizza, V.; Parrinello, D.; Parrinello, N. Hemocytes of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) and their response to Saccharomyces cerevisiae and Bacillus thuringiensis. J. Invertebr. Pathol. 2011, 106, 360–365. [Google Scholar] [CrossRef]
- Wang, G.; Zhou, Y.; Tang, B.; Ali, H.; Hou, Y. Immune function differences between two color morphs of the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae) at different life stages. Ecol. Evol. 2021, 11, 5702–5712. [Google Scholar] [CrossRef]
- Cappa, F.; Torrini, G.; Mazza, G.; Inghilesi, A.F.; Benvenuti, C.; Villani, L.; Roversi, P.F.; Cervo, R. Assessing immunocompetence in red palm weevil adult and immature stages in response to bacterial challenge and entomopathogenic nematode infection. Insect Sci. 2020, 27, 1031–1042. [Google Scholar] [CrossRef]
- Ibrahim, M.; Loulou, A.; Brouk, A.; Muller, A.; Machado, R.A.; Kallel, S. Parasites rather than phoronts: Teratorhabditis synpapillata nematodes reduce lifespan of their Rhynchophorus ferrugineus host in a life stage-dependent manner. Ecol. Evol. 2021, 11, 12596–12604. [Google Scholar] [CrossRef]
- Manachini, B.; Schillaci, D.; Arizza, V. Biological Responses of Rhynchophorus ferrugineus (Coleoptera: Curculionidae) to Steinernema carpocapsae (Nematoda: Steinernematidae). J. Econ. Entomol. 2013, 106, 8. [Google Scholar] [CrossRef]
- Mastore, M.; Arizza, V.; Manachini, B.; Brivio, M.F. Modulation of immune responses of Rhynchophorus ferrugineus (Insecta, Coleoptera) induced by the entomopathogenic nematode Steinernema carpocapsae (Nematoda, Rhabditida). Insect Sci. 2015, 22, 748–760. [Google Scholar] [CrossRef]
- Sadawy, H.A.E.; Namaky, A.H.E.; Omari, F.A.; Bahareth, O.M. Susceptibility of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) to entomopathogenic nematodes with regard to its immune response. Biol. Control 2020, 148, 104308. [Google Scholar] [CrossRef]
- Banerjee, A.; Dangar, T.K. Pseudomonas aeruginosa, a facultative pathogen of red palm weevil, Rhynchophorus ferrugineus. World J. Microbiol. Biotechnol. 1995, 11, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Salama, H.S.; Foda, M.S.; Bendary, M.A.E.; Razek, A.A. Infection of red palm weevil, Rhynchophorus ferrugineus, by spore-forming bacilli indigenous to its natural habitat in Egypt. J. Pest Sci. 2004, 77, 27–31. [Google Scholar] [CrossRef]
- Raio, A.; Roversi, P.F.; Francardi, V. Bacteria associated to Rhynchophorus ferrugineus (Olivier) (Coleoptera Dryophthoridae) in Italy. Redia 2016, 99, 53–57. [Google Scholar]
- Bravo, A.; Gill, S.S.; Soberon, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojda, I.; Taszłow, P. Heat shock affects host–pathogen interaction in Galleria mellonella infected with Bacillus thuringiensis. J. Insect Physiol. 2013, 59, 894–905. [Google Scholar] [CrossRef]
- Manachini, B.; Billeci, N.; Palla, F. Exotic insect pests: The impact of the Red Palm Weevil on natural and cultural heritage in Palermo (Italy). J. Cult. Herit. 2013, 145, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, L.A.; Poulin, R.; Ward, D.F.; Zuk, M. Sex differences in parasitic infections among arthropod hosts: Is there a male bias? Oikos 2000, 88, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Sułek, M.; Kordaczuk, J.; Wojda, I. Current understanding of immune priming phenomena in insects. J Invertebr. Pathol. 2021, 185, 107656. [Google Scholar] [CrossRef]
- Hoffmann, J.A. Innate immunity of insects. Curr. Opin. Immunol. 1995, 7, 4–10. [Google Scholar] [CrossRef]
- Tsakas, S.; Marmaras, V.J. Insect immunity and its signalling: An overview. Invertebr. Surviv. J. 2010, 7, 228–238. [Google Scholar]
- Li, S.; Xu, X.; Shakeel, M.; Xu, J.; Zheng, Z.; Zheng, J.; Yu, X.; Zhao, Q.; Jin, F. Bacillus thuringiensis suppresses the humoral immune system to overcome defense mechanism of Plutella xylostella. Front. Physiol. 2018, 9, 1478. [Google Scholar] [CrossRef] [PubMed]
- Kojour, M.A.M.; Han, Y.S.; Jo, Y.H. An overview of insect innate immunity. Entomol. Res. 2020, 50, 282–291. [Google Scholar] [CrossRef]
- Critchlow, J.T.; Norris, A.; Tate, A.T. The legacy of larval infection on immunological dynamics over metamorphosis. Philos. Trans. R. Soc. B 2019, 374, 20190066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson-Rich, N.; Dres, S.T.; Starks, P.T. The ontogeny of immunity: Development of innate immune strength in the honey bee (Apis mellifera). J. Insect Physiol. 2008, 54, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Roitberg, B.D.; Rosengaus, R.B. Intersection between parental investment, transgenerational immunity, and termite sociality in the face of disease: A theoretical approach. Behav. Ecol. Sociobiol. 2022, 76, 1–28. [Google Scholar] [CrossRef]
- Stoepler, T.M.; Castillo, J.C.; Lill, J.T.; Eleftherianos, I. Hemocyte density increases with developmental stage in an immune-challenged forest caterpillar. PLoS ONE 2013, 8, e70978. [Google Scholar] [CrossRef] [Green Version]
- Pandey, J.P.; Tiwari, R.K. An Overview of Insect Hemocyte Science and its Future Application in Applied and Biomedical Fields. Am. J. Biochem. Mol. Biol. 2012, 2, 82–105. [Google Scholar] [CrossRef] [Green Version]
- Brivio, M.F.; Mastore, M.; Nappi, A.J. A pathogenic parasite interferes with phagocytosis of insect immunocompetent cells. Dev. Comp. Immunol. 2010, 34, 991–998. [Google Scholar] [CrossRef]
- Zhang, C.R.; Zhang, S.; Xia, J.; Li, F.F.; Xia, W.Q.; Shu-Sheng, L.; Xiao, W.W. The immune strategy and stress response of the Mediterranean species of the Bemisia tabaci complex to an orally delivered bacterial pathogen. PLoS ONE 2014, 9, e94477. [Google Scholar] [CrossRef]
- Ritossa, F. A new puffing pattern induced by temperature shock and DNP in drosophila. Experientia 1962, 18, 571–573. [Google Scholar] [CrossRef]
- Sonoda, S.; Tsumuki, H. Induction of heat shock protein genes by chlorfenapyr in cultured cells of the cabbage armyworm, Mamestra brassicae. Pestic. Biochem. Physiol. 2007, 89, 185–189. [Google Scholar] [CrossRef]
- Yoshimi, T.; Minowa, K.; Renier, N.K.K.; Watanabe, C.; Sugaya, Y.; Miura, T. Activation of a stress-induced gene by insecticides in the midge, Chironomus yoshimatsui. J. Biochem. Mol. Toxicol. 2002, 16, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Chen, X.; Liu, W.; Zhang, Z.; Wang, Y.; You, K.; Li, Y.; Zhang, R.; Zhou, Q. Characterization of heat shock protein 70 transcript from Nilaparvata lugens (Stål): Its response to temperature and insecticide stresses. Pestic. Biochem. Physiol. 2017, 142, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Kausar, S.; Abbas, M.N.; Yang, L.; Cui, H. Biotic and abiotic stress induces the expression of Hsp70/90 organizing protein gene in silkworm, Bombyx mori. Int. J. Biol. Macromol. 2020, 143, 610–618. [Google Scholar] [CrossRef]
- Lyupina, Y.V.; Dmitrieva, S.B.; Timokhova, A.V.; Beljelarskaya, S.N.; Zatsepina, O.G.; Evgen’ev, M.B.; Mikhailov, V.S. An important role of the heat shock response in infected cells for replication of baculoviruses. Virology 2010, 406, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Wojda, I.; Jakubowicz, T. Humoral immune response upon mild heat-shock conditions in Galleria mellonella larvae. J. Insect Physiol. 2007, 53, 1134–1144. [Google Scholar] [CrossRef]
- Celi, M.; Vazzana, M.; Manachini, B.R.I.; Parrinello, N.; Arizza, V. In vivo modulation of Hsp70 in Rhynchophorus ferrugineus hemocytes after Bacillus thuringiensis treatment. In Proceedings of the 43th Annual Meeting of the Society for Invertebrate Pathology, 10th International Colloquium on Invertebrate Pathology and Microbial Control, and The Final Meeting of COST862: Bacterial Toxins for Insect Control, Karadeniz Technical University, Trabzon, Turkey, 11–15 July 2010; p. 120. [Google Scholar]
- King, A.M.; MacRae, T.H. Insect heat shock proteins during stress and diapause. Annu. Rev. Entomol. 2015, 60, 59–75. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Celi, M.; Filiciotto, F.; Parrinello, D.; Buscaino, G.; Damiano, A.; Cuttitta, A.; D’Angelo, S.; Mazzola, S.; Vazzana, M. Physiological and agonistic behavioural response of Procambarus clarkii to an acoustic stimulus. J. Exp. Biol. 2013, 216, 709–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finney, D.J. Probit Analysis: A Statistical Treatment of the Sigmoid Response Curve; Cambridge University Press: Cambridge, UK, 1952. [Google Scholar]
- Antony, B.; Johny, J.; Montagné, N.; Jacquin-Joly, E.; Capoduro, R.; Cali, K.; Persaud, K.; Al-Saleh, M.A.; Pain, A. Pheromone receptor of the globally invasive quarantine pest of the palm tree, the red palm weevil (Rhynchophorus ferrugineus). Mol. Ecol. 2021, 30, 2025–2039. [Google Scholar] [CrossRef] [PubMed]
- Milosavljević, I.; El-Shafie, H.A.F.; Faleiro, J.R.; Hoddle, C.D.; Lewis, M.; Hoddle, M.S. Palmageddon: The wasting of ornamental palms by invasive palm weevils, Rhynchophorus spp. J. Pest Sci. 2019, 92, 143–156. [Google Scholar] [CrossRef]
- Meftaul, I.M.; Venkateswarlu, K.; Dharmarajan, R.; Annamalai, P.; Megharaj, M. Pesticides in the urban environment: A potential threat that knocks at the door. Sci. Total Environ. 2020, 711, 134612. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; Freed, S. Biochemical resistance mechanisms against chlorpyrifos, imidacloprid and lambda-cyhalothrin in Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae). Crop Prot. 2021, 143, 105568. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Quintero, H.A.; Serrão, J.E.; Martínez, L.C. Insecticidal activity of Bacillus thuringiensis strains on the nettle caterpillar, Euprosterna elaeasa (Lepidoptera: Limacodidae). Insects 2020, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Cho, S. Characterization of the immune hemocyte in larvae of Dorcus titanus castanicolor (Motschulsky, 1861) (Lucanidae, Coleoptera). Entomol. Res. 2021, 51, 445–452. [Google Scholar] [CrossRef]
- Makki, R.; Cinnamon, E.; Gould, A.P. The Development and functions of oenocytes. Annu. Rev. Entomol. 2014, 59, 405–425. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, T.; Krejcova, G.; Bajgar, A.; Nedbalova, P.; Strasser, P. Molecular regulations of metabolism during immune response in insects. Insect Biochem. Mol. Biol. 2019, 109, 31–42. [Google Scholar] [CrossRef]
- Martins, G.F.; Ramalho-Ortigao, J.M. Oenocytes in insects. Invertebr. Surviv. J. 2013, 9, 139–152. [Google Scholar]
- Schmitz, A.; Anselme, C.; Ravallec, M.; Rebuf, C.; Simon, J.C.; Gatti, J.L.; Poirié, M. The cellular immune response of the pea aphid to foreign intrusion and symbiotic challenge. PLoS ONE 2012, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyetta, N.R.; Ramos, F.O.; Leyria, J.; Canavoso, L.E.; Fruttero, L.L. Morphological and ultrastructural characterization of hemocytes in an insect model, the hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae). Insects 2021, 12, 640. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindquist, S. Regulation of protein synthesis during heat shock. Nature 1981, 293, 311–314. [Google Scholar] [CrossRef]
- Li, M.Y.; Huang, Y.; Lei, X.; Xu, C.T.; Li, B.; Chen, D.X.; Liu, S. Identification of six heat shock protein 70 genes in Lasioderma serricorne (Coleoptera: Anobiidae) and their responses to temperature stress. J. Asia-Pac. Entomol. 2021, 24, 597–605. [Google Scholar] [CrossRef]
- Breitenbach, J.E.; Shelby, K.S.; Popham, H.J. Baculovirus induced transcripts in hemocytes from the larvae of Heliothis virescens. Viruses 2011, 3, 2047–2064. [Google Scholar] [CrossRef]
- Breitenbach, J.E.; Popham, H.J. Baculovirus replication induces the expression of heat shock proteins in vivo and in vitro. Arch. Virol. 2013, 158, 1517–1522. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Wu, G.X.; Ye, G.Y.; Hu, C. Heat shock protein genes (hsp20, hsp75 and hsp90) from Pieris rapae: Molecular cloning and transcription in response to parasitization by Pteromalus puparum. Insect Sci. 2013, 20, 183–193. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Denlinger, D.L. Heat-shock protein 90 is down-regulated during pupal diapause in the flesh fly, Sarcophaga crassipalpis, but remains responsive to thermal stress. Insect Mol. Biol. 2000, 9, 641–645. [Google Scholar] [CrossRef]
- Yocum, G.D.; Joplin, K.H.; Denlinger, D.L. Upregulation of a 23 kDa small heat shock protein transcript during pupal diapause in the flesh fly, Sarcophaga crassipalpis. Insect Biochem. Mol. Biol. 1998, 28, 677–682. [Google Scholar] [CrossRef]
- Nakasu, E.Y.; Firmino, A.A.; Dias, S.C.; Rocha, T.L.; Ramos, H.B.; Oliveira, G.R.; Lucena, W.; Carlini, C.R.; Grossi-De-Sá, M.F. Analysis of Cry8Ka5-binding proteins from Anthonomus grandis (Coleoptera: Curculionidae) midgut. J. Invertebr. Pathol. 2010, 104, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Z.; Liang, G.M.; Zhang, J.; Wu, K.M.; Guo, Y.Y.; Rector, B.G. Proteomic analysis of novel Cry1Ac binding proteins in Helicoverpa armigera (Hübner). Arch. Insect Biochem. Physiol. 2010, 73, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Wojda, I.; Kowalski, P. Galleria mellonella infected with Bacillus thuringiensis involves Hsp90. Cent. Eur. J. Biol. 2013, 8, 561–569. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. I | LC | EC | 95% CI | Slope ± SE | X2 (p-Value) | |
|---|---|---|---|---|---|---|
| Larva | 330 | LC50 | 1.22 | 0.23–3.18 | 2.03 ± 0.09 | 7.44 (0.10) |
| Females | 330 | LC50 | 2.29 | 1.89–4.92 | 1.99 ± 0.11 | 3.77 (0.44) |
| Males | 330 | LC50 | 1.85 | 0.78–7.49 | 2.35 ± 0.10 | 6.54 (0.16) |
| No. I | LT50 | Intercept ± SE | Slope ± SE | CC | LT50 | 95% CI | |
|---|---|---|---|---|---|---|---|
| Larvae | 40 | 36.82 | 5.96 ± 0.09 | 3.80± 0.45 | 0.940 | 36.82 | 32.76–40.95 |
| Females | 40 | 44.06 | 4.90 ± 0.10 | 2.98 ± 0.39 | 0.963 | 44.06 | 38.63–50.47 |
| Males | 40 | 37.34 | 5.23 ± 0.11 | 3.28 ± 0.41 | 0.984 | 37.34 | 32.78–42.90 |
| Times | 0 h | 3 h | 6 h | 12 h | 24 h |
|---|---|---|---|---|---|
| Plasmatocytes (×106/mL) | |||||
| L | 2.35 ± 0.45 a | 1.99 ± 0.78 a,k | 1.19 ± 0.33 t * | 0.78 ± 0.35 ** α | 0.26 ± 0.04 ** ι |
| F | 3.42 ± 1.64 a | 1.53 ± 2.35 a,k | 0.95 ± 0.83 a,t,u | 0.3 ± 0.00 * α,β | 0.18 ± 0.08 * ι |
| M | 0.87 ± 0.25 b | 0.37 ± 0.13 * l | 0.28 ± 0.03 u * | 0.5 ± 0.36 b,β | 0.55 ± 0.3 b,ι |
| Granulocytes (×106/mL) | |||||
| L | 1.52 ± 0.44 c | 1.53 ± 0.38 c,m | 1.52 ± 0.33 c,v | 1.55 ± 0.09 χ | 1.64 ± 0.2 c |
| F | 7.97 ± 1.70 d | 1.7 ± 0.33 ** m | 1.38 ± 1.05 ** m,v,z | 0.8 ± 0.01 δ ** | 0.32 ± 0.13 ** λ |
| M | 4.38 ± 2.3 e | 2.12 ± 0.97 e,m | 0.5 ± 0.05 * w | 1.65 ± 0.83 e,χ | 0.21 ± 0.10 * λ |
| Oenocytes (×106/mL) | |||||
| L | 0.19 ± 0.04 f | 0.36 ± 0.17 f,n | 0.59 ± 0.28 f,x | 0.67 ± 0.12 ** δ | 0.44 ± 0.09 * μ |
| F | 0.15 ± 0.02 f | 0.05 ± 0.01 *** o | 0.07 ± 0.02 ** y | 0.05 ± 0.02 *** ε | 0.03 *** μ |
| M | 0.23 ± 0.19 f | 0.08 ± 0.02 f,o | 0 | 0.08 ± 0.06 f,ε | 0 |
| Prohemocytes (×106/mL) | |||||
| L | 0.32 ± 0.14 g | 0.23 ± 0.06 g,p | 0.37 ± 0.09 g,y | 0.44 ± 0.09 g,φ | 0.22 ± 0.06 g |
| F | 0.43 ± 0.05 g | 0 *** | 0.002 ± 0.000 *** z | 0 *** γ | 0 *** |
| M | 0.05 ± 0.05 h | 0.03 ± 0.02 b,q | 0 | 0 | 0.03 ± 0.00 ν |
| Spherulocytes (×106/mL) | |||||
| L | 0.14 ± 0.08 i | 0.23 ± 0.05 i,r | 0.35 ± 0.19 i,r, | 0.38 ± 0.08 * η | 0.18 ± 0.08 i |
| F | 0 | 0 | 0 | 0 | 0 |
| M | 0.09 ± 0.01 j | 0.001 ± 0.000 j,s | 0 | 0.0015 ± 0.000 j | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Celi, M.; Russo, D.; Vazzana, M.; Arizza, V.; Manachini, B. Does Bacillus thuringiensis Affect the Stress and Immune Responses of Rhynchophorus ferrugineus Larvae, Females, and Males in the Same Way? Insects 2022, 13, 437. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050437
Celi M, Russo D, Vazzana M, Arizza V, Manachini B. Does Bacillus thuringiensis Affect the Stress and Immune Responses of Rhynchophorus ferrugineus Larvae, Females, and Males in the Same Way? Insects. 2022; 13(5):437. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050437
Chicago/Turabian StyleCeli, Monica, Debora Russo, Mirella Vazzana, Vincenzo Arizza, and Barbara Manachini. 2022. "Does Bacillus thuringiensis Affect the Stress and Immune Responses of Rhynchophorus ferrugineus Larvae, Females, and Males in the Same Way?" Insects 13, no. 5: 437. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050437