
Characterization of Potential Molecular Markers in Lac Insect Kerria lacca (Kerr) Responsible for Lac Production


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
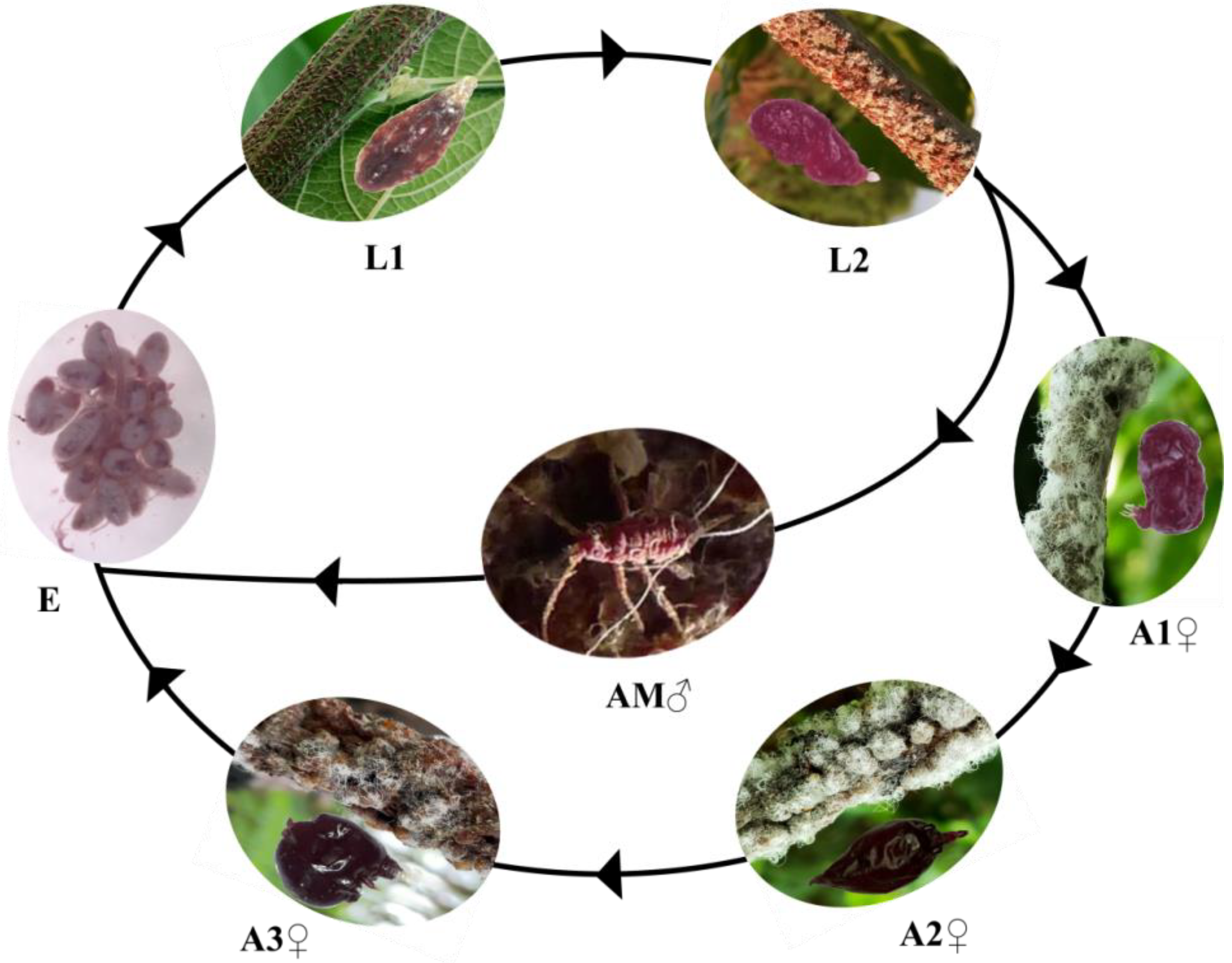
2.1. Insect Material
2.2. Sequencing and Variant Identification
2.3. Annotation of Single-Nucleotide Polymorphisms and Insertion/Deletion Polymorphism
2.4. Identification of Lac Production-Related Genes with Linked Variants
3. Results
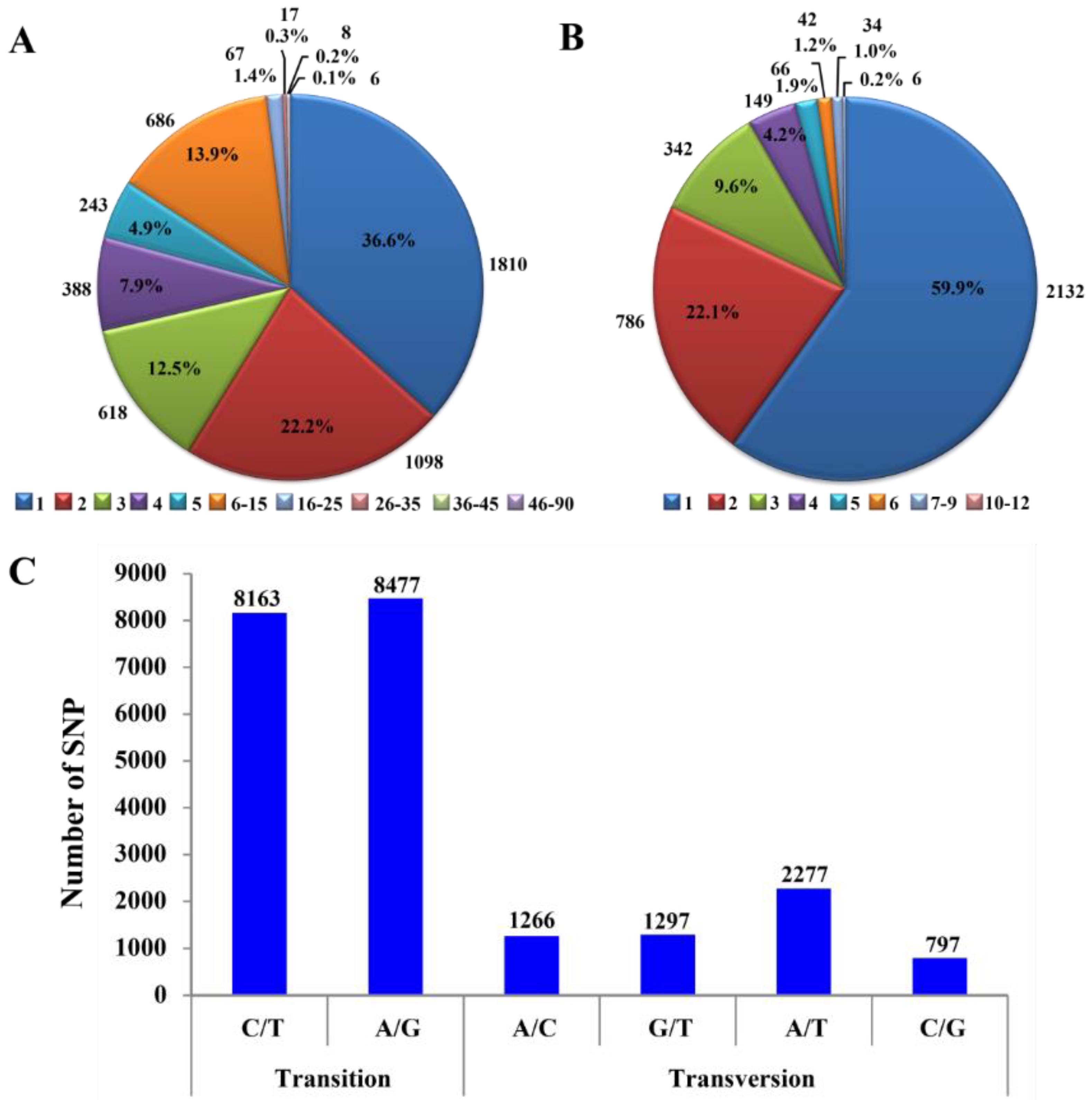
3.1. SNP and InDel Variant Identification
3.2. SNP Substitutions at Different Stages of Kerria Lacca
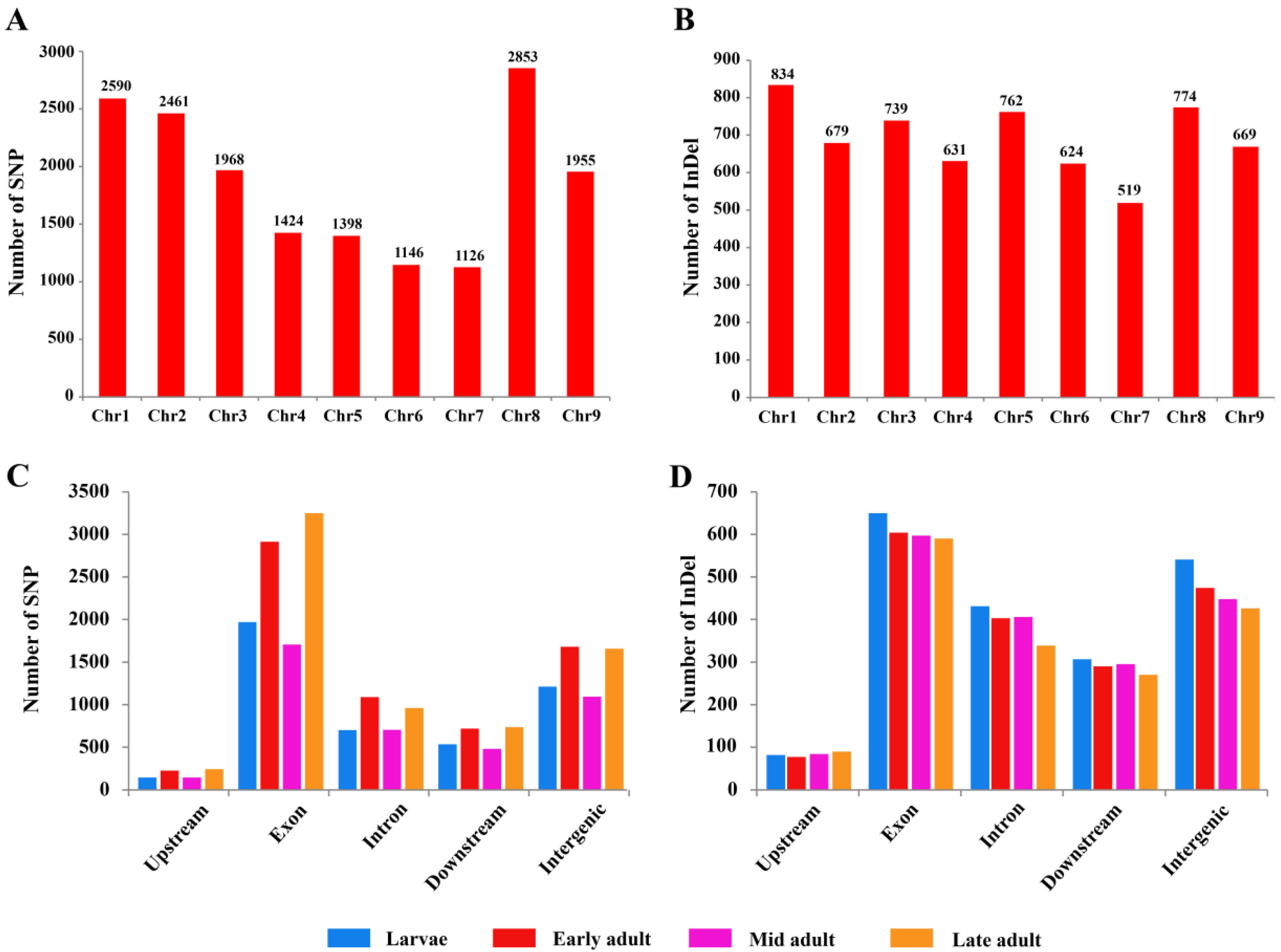
3.3. SNP and InDel Chromosomal Distribution
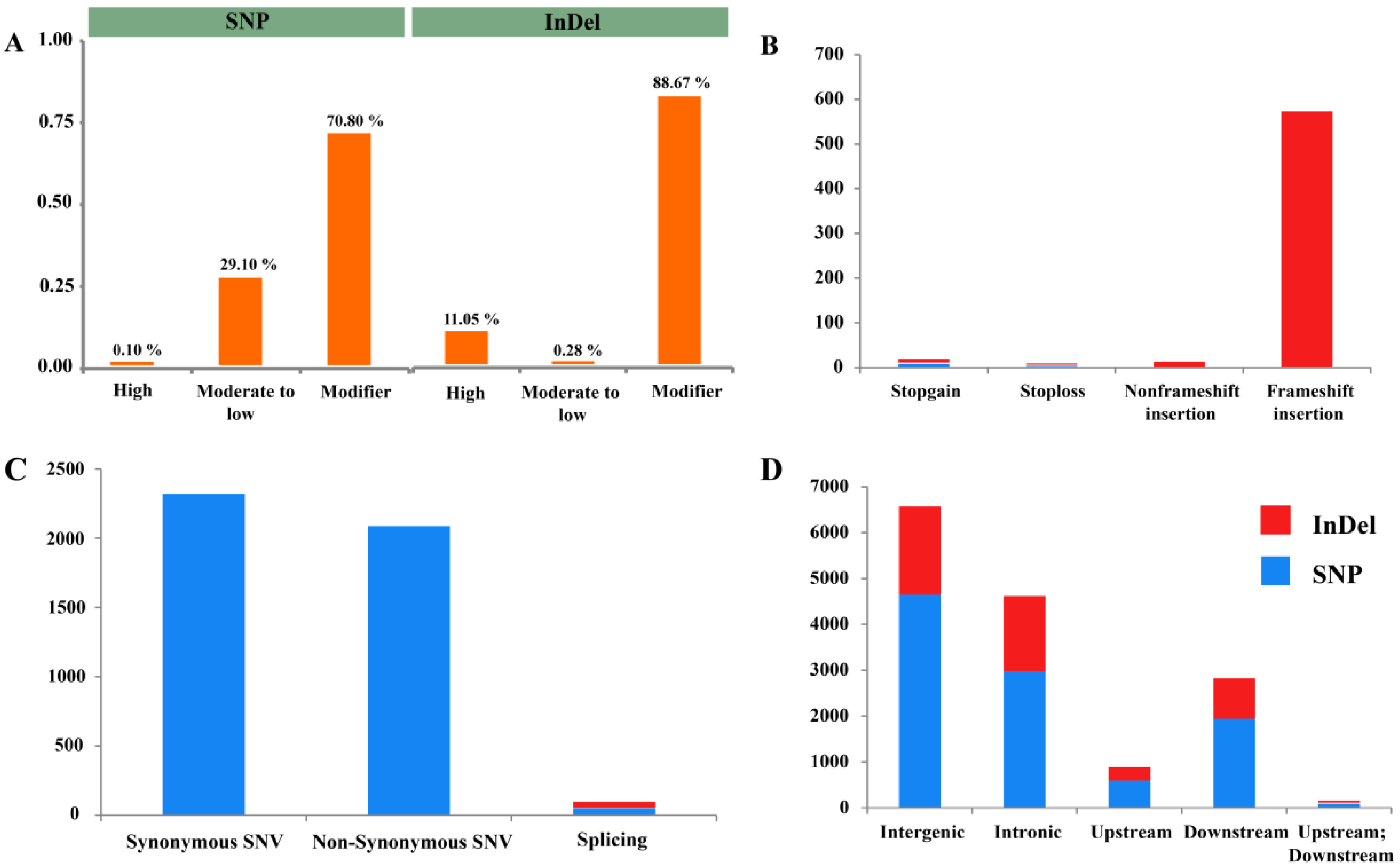
3.4. SNP and InDel Annotation
3.5. Functional Classification Variants
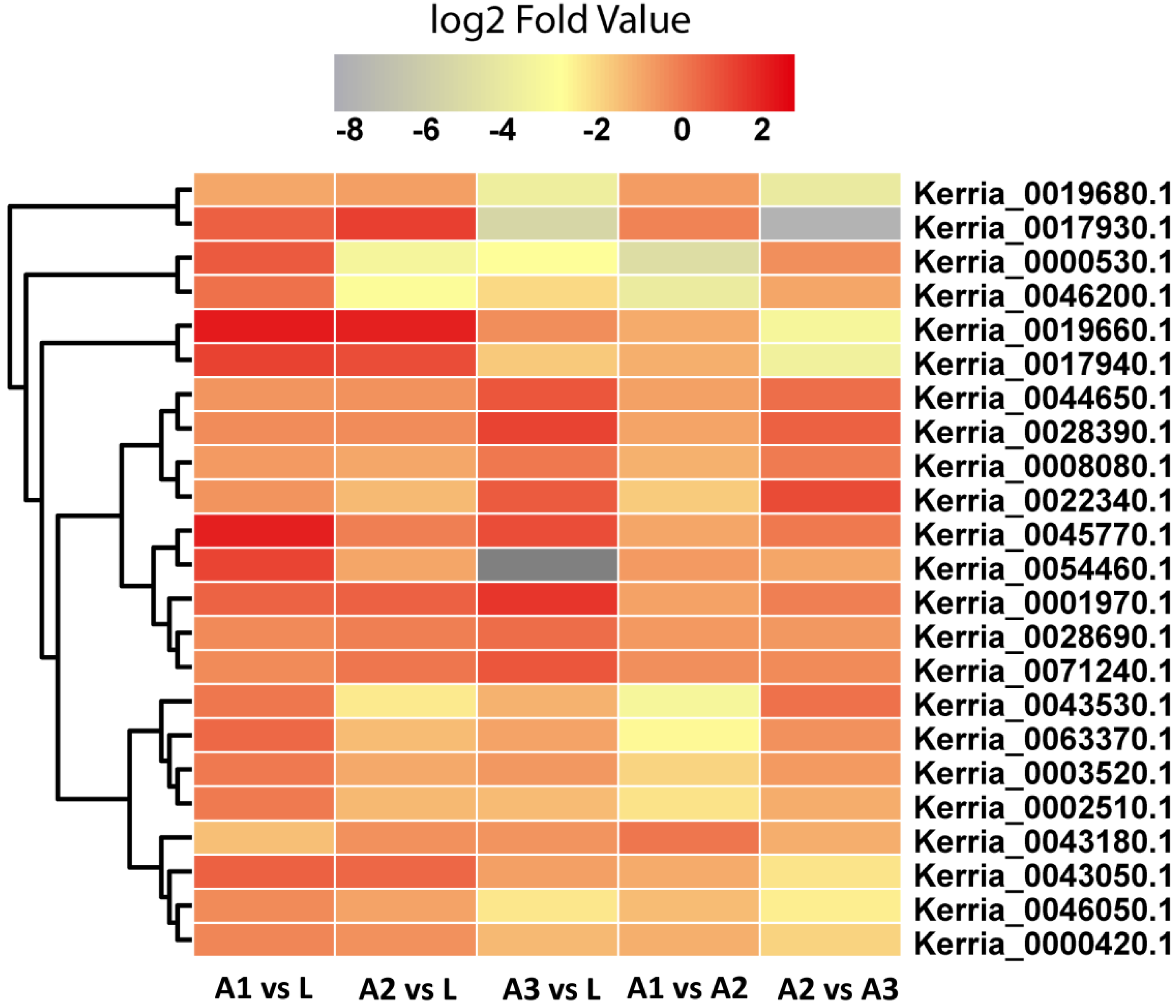
3.6. Identification of Lac Synthesis Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kandasamy, T.; Ekbal, S.; Kumari, K.; Lohot, V.D.; Mohanasundaram, A.; Sharma, K.K. Unraveling bacterial diversity of the Indian Lac Insect Kerria lacca (Kerr) using next generation sequencing. Int. J. Trop. Insect Sci. 2022, 42, 2365–2372. [Google Scholar] [CrossRef]
- Netam, P.K.; Netam, P.K.; Markam, P.S. Population dynamics of associated fauna of lac insect Kerria lacca (Kerr.) in kusum host plant at Kanker district of Chhattisgarh. Pharm. Innov. J. 2021, 10, 359–362. [Google Scholar]
- Borah, N.; Garkoti, S.C. Indigenous lac culture and local livelihood: A case study of Karbi community of Assam, North-Eastern India. Indian J. Tradit. Knowl. 2020, 19, 197–207. [Google Scholar]
- Wang, W.W.; Liu, P.F.; Lu, Q.; Ling, X.F.; Zhang, J.W.; Chen, M.S.; Chen, H.; Chen, X.M. Potential Pathways and Genes Involved in Lac Synthesis and Secretion in Kerria chinensis (Hemiptera: Kerriidae) Based on Transcriptomic Analyses. Insects 2019, 10, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.M. Biodiversity of Lac Insects; Yunnan Science and Technology Press: Kunming, China, 2005; p. 96. [Google Scholar]
- Rohmer, M. The mevalonate-independent methylerythritol 4-phosphate (MEP) pathway for isoprenoid biosynthesis, including carotenoids. Pure Appl. Chem. 1999, 71, 2279–2284. [Google Scholar] [CrossRef]
- Bede, J.C.; Teal, P.E.; Goodman, W.G.; Tobe, S.S. Biosynthetic Pathway of Insect Juvenile Hormone III in Cell Suspension Cultures of the Sedge Cyperus iria. Plant Physiol. 2001, 127, 584–593. [Google Scholar] [CrossRef]
- Hsieh, M.-H.; Goodman, H.M. The Arabidopsis IspH Homolog Is Involved in the Plastid Nonmevalonate Pathway of Isoprenoid Biosynthesis. Plant Physiol. 2005, 138, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Shamim, G.; Pandey, D.M.; Ramani, R.; Sharma, K.K. Identification of genes related to resin biosynthesis in the Indian lac insect, Kerria lacca (Hemiptera: Tachardiidae). Int. J. Trop. Insect Sci. 2014, 34, 149–155. [Google Scholar] [CrossRef]
- Chen, H.; Wang, W.W.; Bashir, N.H. The Heredity and Evolution of Lac Insects; Science Press: Beijing, China, 2021; p. 108. [Google Scholar]
- Bashir, N.H.; Wang, W.W.; Liu, J.; Wang, W.; Chen, H. First record of the lac-producing species Kerria nepalensis Varshney (Hemiptera, Kerriidae) from China, with a key to Chinese species. ZooKeys 2021, 1061, 1–9. [Google Scholar] [CrossRef]
- Shah, T.H.; Thomas, M.; Bhandari, R. Lac production, constraints and management: A review. Int. J. Curr. Res. 2015, 7, 13652–13659. [Google Scholar]
- Varshney, R.K.; Sharma, K.K. Lac Insects of the World: An Updated Catalogue and Bibliography; Indian Council of Agricultural Research, Indian Institute of Natural Resins and Gums: Ranchi, India, 2020; p. 84. [Google Scholar]
- Chen, H.; Chen, X.M.; Feng, Y.; Yang, H.; He, R.; Zhang, W.F.; Yang, Z.X. Molecular phylogeny and biogeography of lac insects (Hemiptera: Kerriidae) inferred from nuclear and mitochondrial gene sequences. Mol. Biol. Rep. 2013, 40, 5943–5952. [Google Scholar] [CrossRef] [PubMed]
- Ramani, R.; Sharma, K.K. Lac Insects. In Beneficial Insects Farming; Sharma, K.K., Monobrullah, M., Mohanasundaram, A., Ramani, R., Eds.; ICAR-Indian Institute of Natural Resins and Gums: Ranchi, India, 2016; pp. 53–65. [Google Scholar]
- Mohanta, J.; Dey, D.G.; Mohanty, N. Life cycle of lac insect, Kerria lacca kerr in Similipal Biosphere Reserve. Indian J. Entomol. 2013, 75, 26–30. [Google Scholar]
- Chen, X.M.; Chen, Y.Q.; Zhang, H.; Shi, L. Lac Insects Breeding and Lac Processing; Chinese Forestry Press: Beijing, China, 2008. [Google Scholar]
- Xu, Y.Q.; Tian, S.Y.; Li, R.Q.; Huang, X.F.; Li, F.Q.; Ge, F.; Huang, W.Z.; Zhou, Y. Transcriptome Characterization and Identification of Molecular Markers (SNP, SSR, and Indels) in the Medicinal Plant Sarcandra glabra spp. Biomed. Res. Int. 2021, 9990910. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, M.K.; Kale, S.M.; Sriswathi, M.; Naresh, T.; Shasidhar, Y.; Garg, V.; Pandey, M.K.; Varshney, R.K. Genome-Wide Discovery and Deployment of Insertions and Deletions Markers Provided Greater Insights on Species, Genomes, and Sections Relationships in the Genus Arachis. Front. Plant Sci. 2017, 8, 2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Korol, A.B.; Fahima, T.; Beiles, A.; Nevo, E. Microsatellites: Genomic distribution, putative functions and mutational mechanisms: A review. Mol. Ecol. 2002, 11, 2453–2465. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.; Das, P.; Hazarika, L.K.; Sharma, K.K.; Mohanasundaram, A.; Roy, S. Evaluation of certain plant hosts for the lac insect. Int. J. Trop. Insect Sci. 2021, 41, 2585–2592. [Google Scholar] [CrossRef]
- Mishra, Y.; Kumar, P. Lac Culture. In Industrial Entomology; Omkar, Ed.; Springer: Singapore, 2017; pp. 109–155. [Google Scholar]
- Andrews, S. FastQC: A Quality Control Application for High Throughput Sequence Data. Babraham Institute Project Page. 2012. Available online: http://www.bioinformatics.bbsrc.ac.uk/projects/fastqc (accessed on 15 December 2021).
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Brouard, J.-S.; Schenkel, F.; Marete, A.; Bissonnette, N. The GATK joint genotyping workflow is appropriate for calling variants in RNA-seq experiments. J. Anim. Sci. Biotechnol. 2019, 10, 44. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.E.; Sheehan, M.J. Sex differences in deleterious genetic variants in a haplodiploid social insect. BioRxiv 2021, 1–22. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; He, Y.; Xia, R. TBtools, a Toolkit for Biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Wang, M.; Li, H.Y.; Zheng, H.Q.; Zhao, L.W.; Xue, X.F.; Wu, L.M. De Novo Transcriptomic Resources in the Brain of Vespa velutina for Invasion Control. Insects 2020, 11, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.; Liu, F.; Lu, H.; Huang, Y. Characterization and Analysis of a de novo Transcriptome from the Pygmy Grasshopper Tetrix Japonica. Mol. Ecol. Resour. 2017, 17, 381–392. [Google Scholar] [CrossRef]
- Behura, S.K. Molecular marker systems in insects: Current trends and future avenues. Mol. Ecol. 2006, 15, 3087–3113. [Google Scholar] [CrossRef]
- Jain, A.; Roorkiwal, M.; Kale, S.; Garg, V.; Yadala, R.; Varshney, R.K. InDel markers: An extended marker resource for molecular breeding in chickpea. PLoS ONE 2019, 14, e0213999. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.L.; Kong, L.P.; Yu, K.C.; Zhang, F.G.; Shi, X.Y.; Wang, Y.P.; Nan, H.Y.; Zhao, X.H.; Lu, S.J.; Cao, D. Development and validation of InDel markers for identification of QTL underlying flowering time in soybean. Crop J. 2018, 6, 126–135. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | InDel | |||
|---|---|---|---|---|
| Type | Count | Ratio | Count | Ratio |
| 3′UTR | 1601 | 14.15% | 587 | 17.31% |
| 5′UTR | 579 | 5.12% | 288 | 8.50% |
| Exon | 4423 | 39.10% | 592 | 17.46% |
| Intergenic | 4663 | 41.22% | 1909 | 56.30% |
| Splice sites | 47 | 0.42% | 15 | 0.44% |
| Stages | Transitions (Ts) | Transversions (Tv) | Ts/Tv Ratio |
|---|---|---|---|
| Larvae | 3275 | 1314 | 2.49 |
| Early Adult | 4832 | 1828 | 2.64 |
| Mid Adult | 3041 | 1110 | 2.74 |
| Late Adult | 5492 | 1385 | 3.97 |
| Gene ID | Description | Function | Chr | Variant |
|---|---|---|---|---|
| Kerria_0043050.1 | Decaprenyl-diphosphate synthase | TBS | 5 | SNP_0043050.1 [A/G] |
| Kerria_0001970.1 | Decaprenyl-diphosphate synthase | TBS | 1 | SNP_0001980.1 [A/T] |
| Kerria_0000530.1 | Farnesyl pyrophosphate synthase | TBS | 1 | SNP_0000530.1 [G/A] |
| Kerria_0046200.1 | Farnesyl diphosphate synthase | TBS | 5 | SNP_0046200.1 [T/G] |
| Kerria_0043530.1 | Acetyl-CoA acetyltransferase | TBS | 5 | SNP_0043540.1 [A/C] |
| Kerria_0003520.1 | Hydroxymethylglutaryl-CoA reductase | TBS | 1 | SNP_0003520.1 [T/C] |
| Kerria_0063370.1 | Acyl-CoA dehydrogenase | FAM | 9 | SNP_0063370.1 [G/A] |
| Kerria_0044650.1 | Acyl-CoA dehydrogenase | FAM | 5 | SNP_0044650.1 [T/G] |
| Kerria_0028690.1 | Acyl-coenzyme A oxidase | FAM | 8 | SNP_0028690.1 [T/C] |
| Kerria_0019680.1 | Acyl-CoA delta (11) desaturase | FAM | 3 | SNP_0019690.1 [C/T] |
| Kerria_0019660.1 | Very long chain fatty acid elongase | FAM | 3 | SNP_0019660.1 [A/G] |
| Kerria_0043180.1 | Fatty acid synthase | FAB, FAM | 5 | SNP_0043180.1 [A/G] |
| Kerria_0046050.1 | Fatty acid synthase | FAB, FAM | 5 | SNP_0046050.1 [A/G] |
| Kerria_0045770.1 | Fatty acid synthase | FAB, FAM | 5 | SNP_0045780.1 [A/G] |
| Kerria_0028390.1 | Long-chain acyl-CoA synthetase | FAB, FAM | 8 | SNP_0028390.1 [C/T] |
| Kerria_0071240.1 | Long-chain acyl-CoA synthetase | FAB, FAM | 2 | SNP_0071240.1 [G/A] |
| Kerria_0008080.1 | S-malonyltransferase | FAB, FAM | 1 | SNP_0008070.1 [G/A] |
| Kerria_0017940.1 | Elongation-very long chain fatty acids-protein-4 | FAE, FAM | 3 | SNP_0017950.1 [C/T] |
| Kerria_0017930.1 | Elongation-very long chain fatty acids-protein-4 | FAE, FAM | 3 | SNP_0017930.1 [C/T] |
| Kerria_0000420.1 | Elongation-very long chain fatty acids-protein-4 | FAE, FAM | 1 | SNP_0000410.1 [A/C] |
| Kerria_0022340.1 | Elongation-very long chain fatty acids-protein-6 | FAE, FAM | 3 | SNP_0022340.1 [A/G] |
| Kerria_0054460.1 | Elongation-very long chain fatty acids-protein-7 | FAE, FAM | 4 | SNP_0054470.1 [C/T] |
| Kerria_0002510.1 | 3-ketoacyl-CoA thiolase | FAE, FAM | 1 | SNP_0002510.1 [C/T] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashir, N.H.; Wang, W.; Ling, X.; Zhang, J.; Lu, Q.; He, R.; Chen, H. Characterization of Potential Molecular Markers in Lac Insect Kerria lacca (Kerr) Responsible for Lac Production. Insects 2022, 13, 545. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060545
Bashir NH, Wang W, Ling X, Zhang J, Lu Q, He R, Chen H. Characterization of Potential Molecular Markers in Lac Insect Kerria lacca (Kerr) Responsible for Lac Production. Insects. 2022; 13(6):545. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060545
Chicago/Turabian StyleBashir, Nawaz Haider, Weiwei Wang, Xiaofei Ling, Jinwen Zhang, Qin Lu, Rui He, and Hang Chen. 2022. "Characterization of Potential Molecular Markers in Lac Insect Kerria lacca (Kerr) Responsible for Lac Production" Insects 13, no. 6: 545. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060545