
Morphology of the Female Reproductive System of the Soybean Thrips, Neohydatothrips variabilis (Beach, 1896) (Thysanoptera: Thripidae)


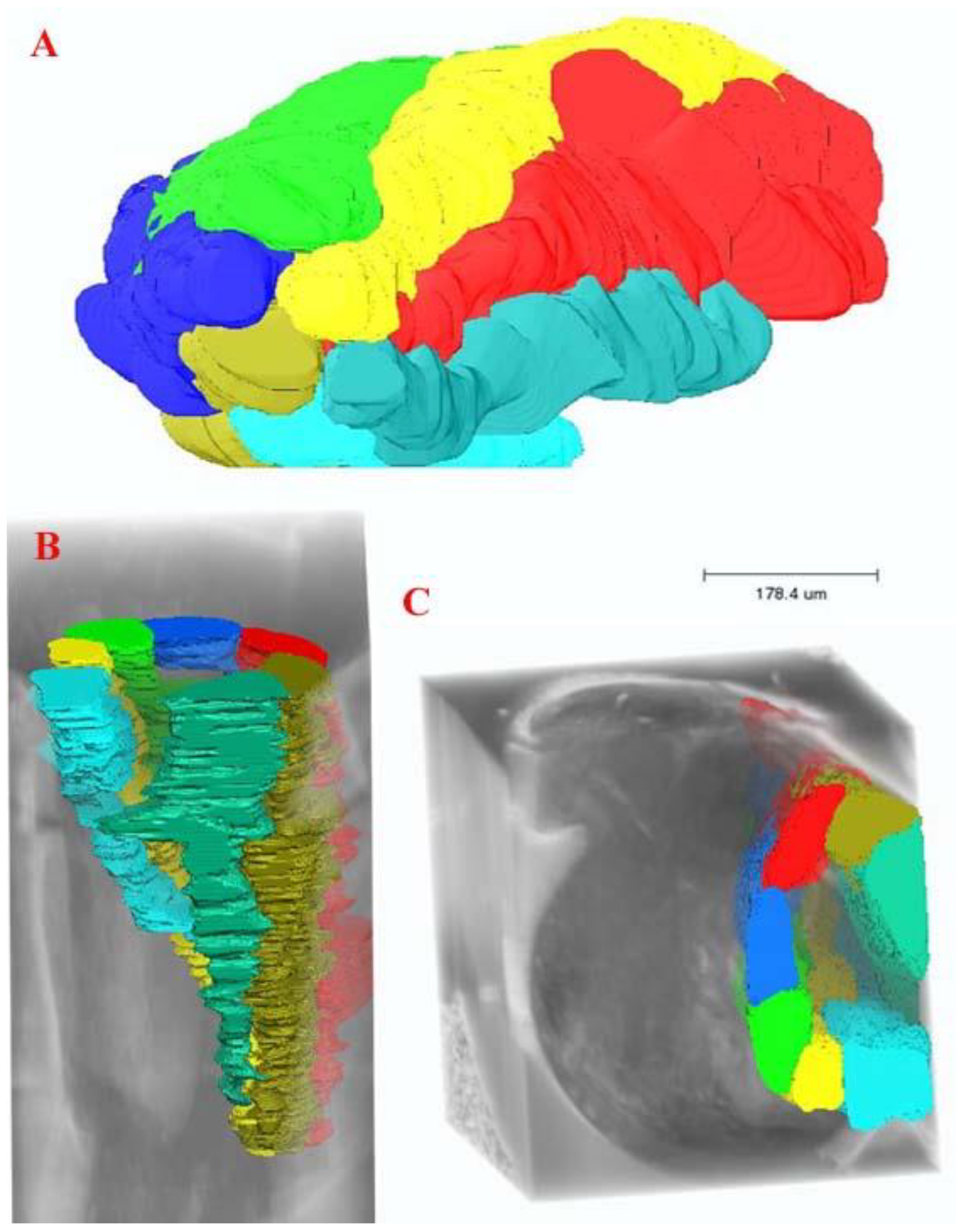{kind=link}
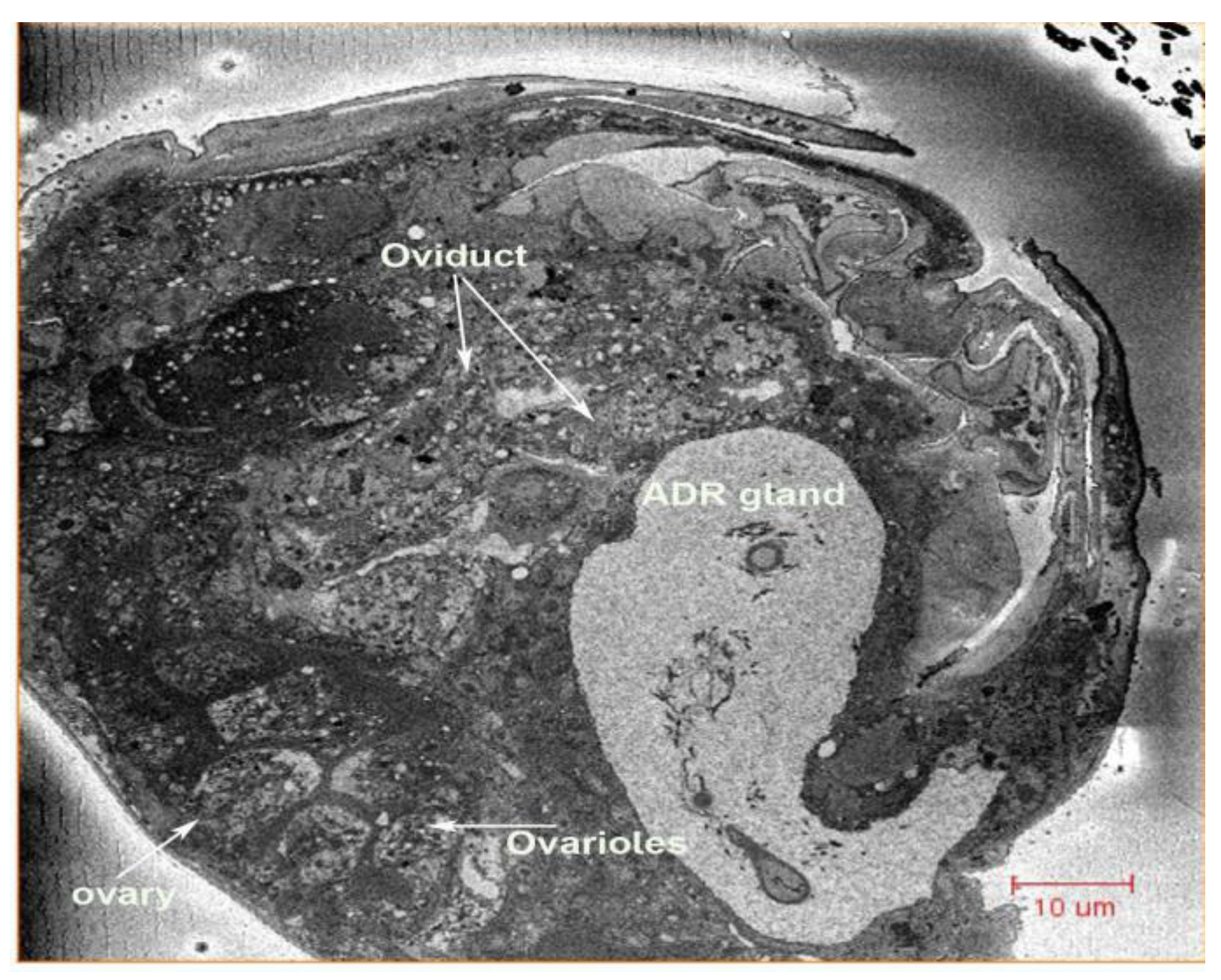{kind=link}
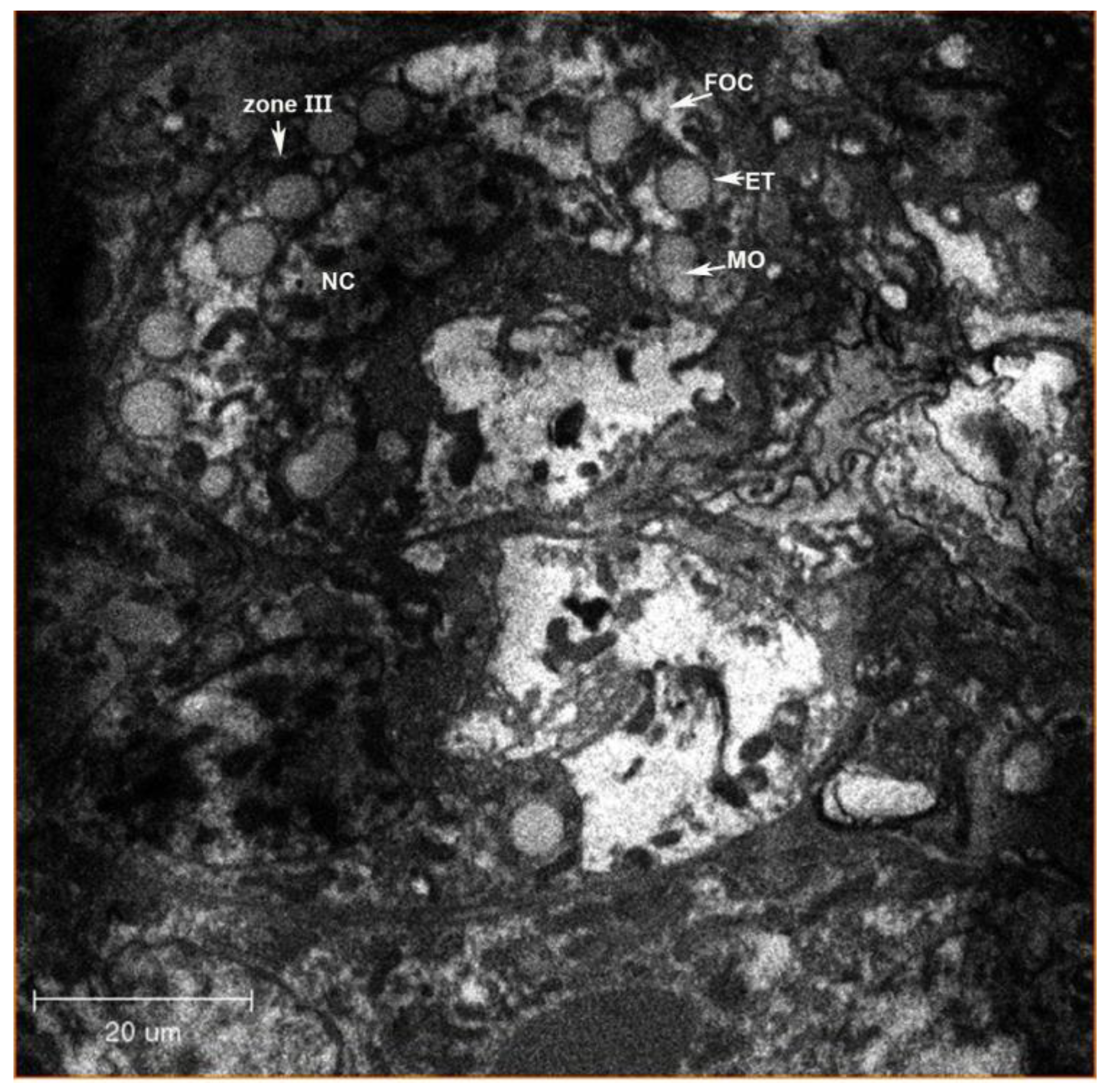{kind=link}
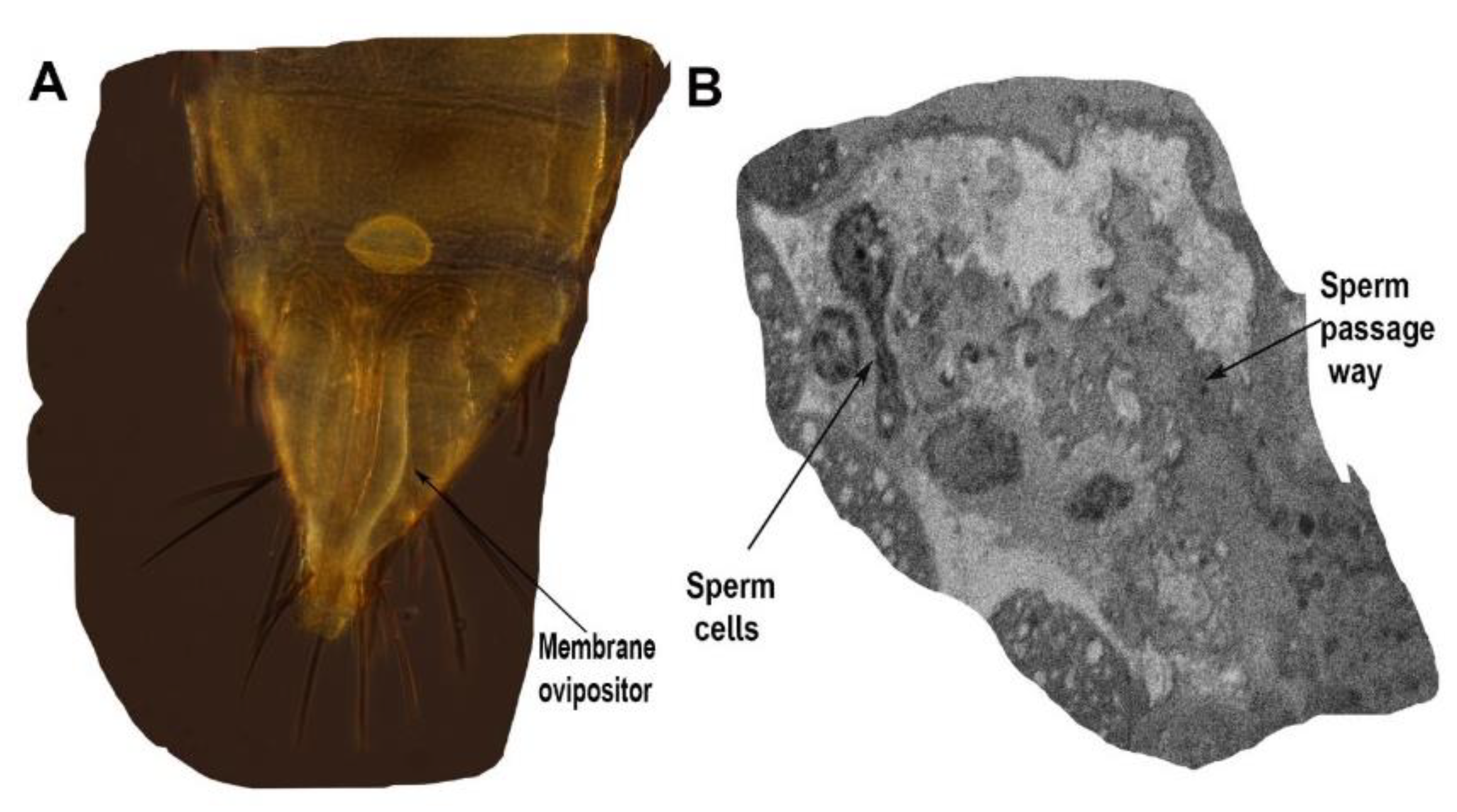{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. Egg-Laying Behavior
2.2.1. Fuchsin Staining of Plant Tissues
2.2.2. Histology of Egg-Containing Leaves
2.3. Morphology of Female Terminalia
2.3.1. Terminology
2.3.2. Dissections
2.3.3. CLSM
2.3.4. SBFSEM
3. Results
3.1. Female Terminalia of N. variabilis
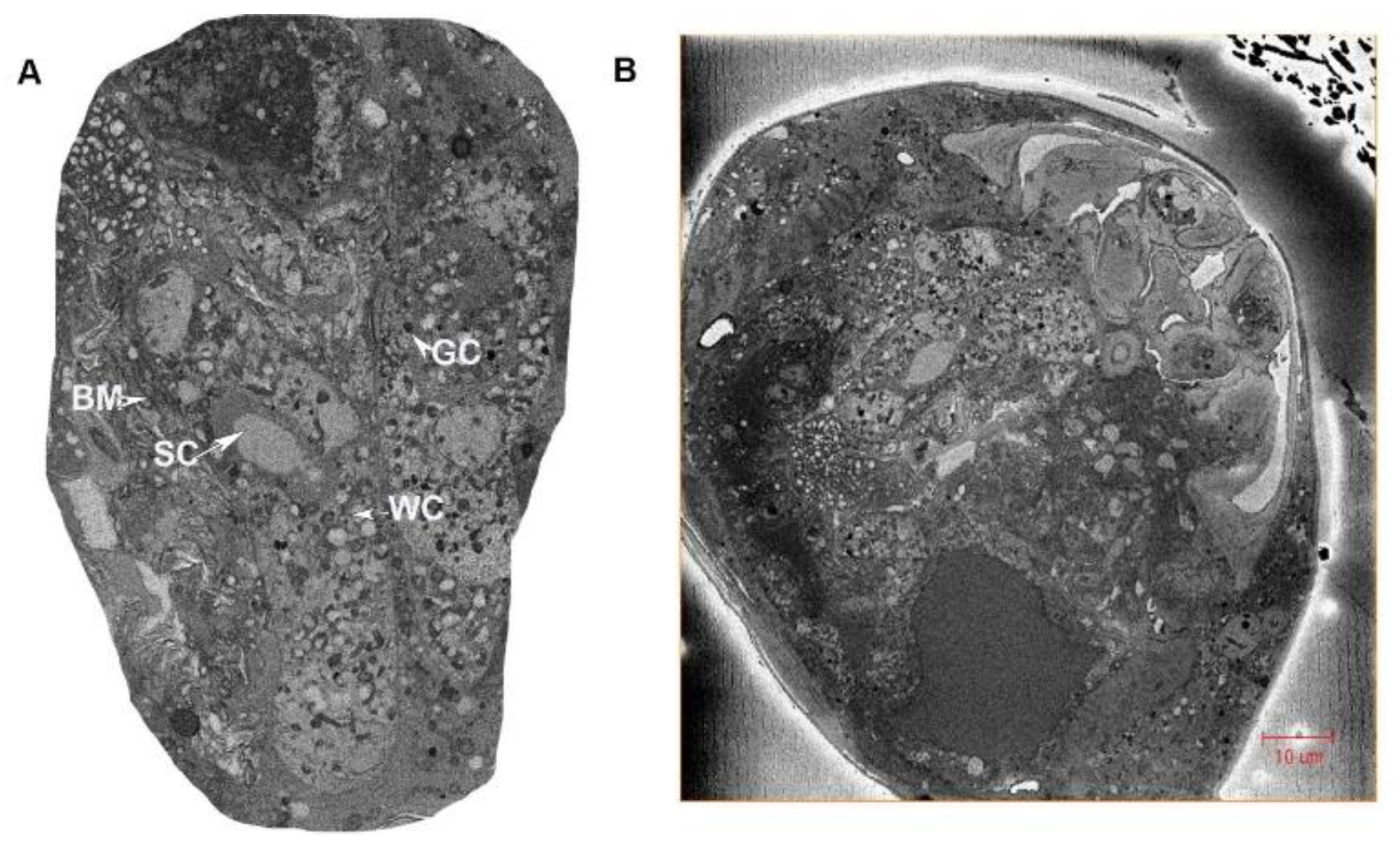
3.1.1. Ovaries
3.1.2. Oviducts
3.1.3. Spermatheca
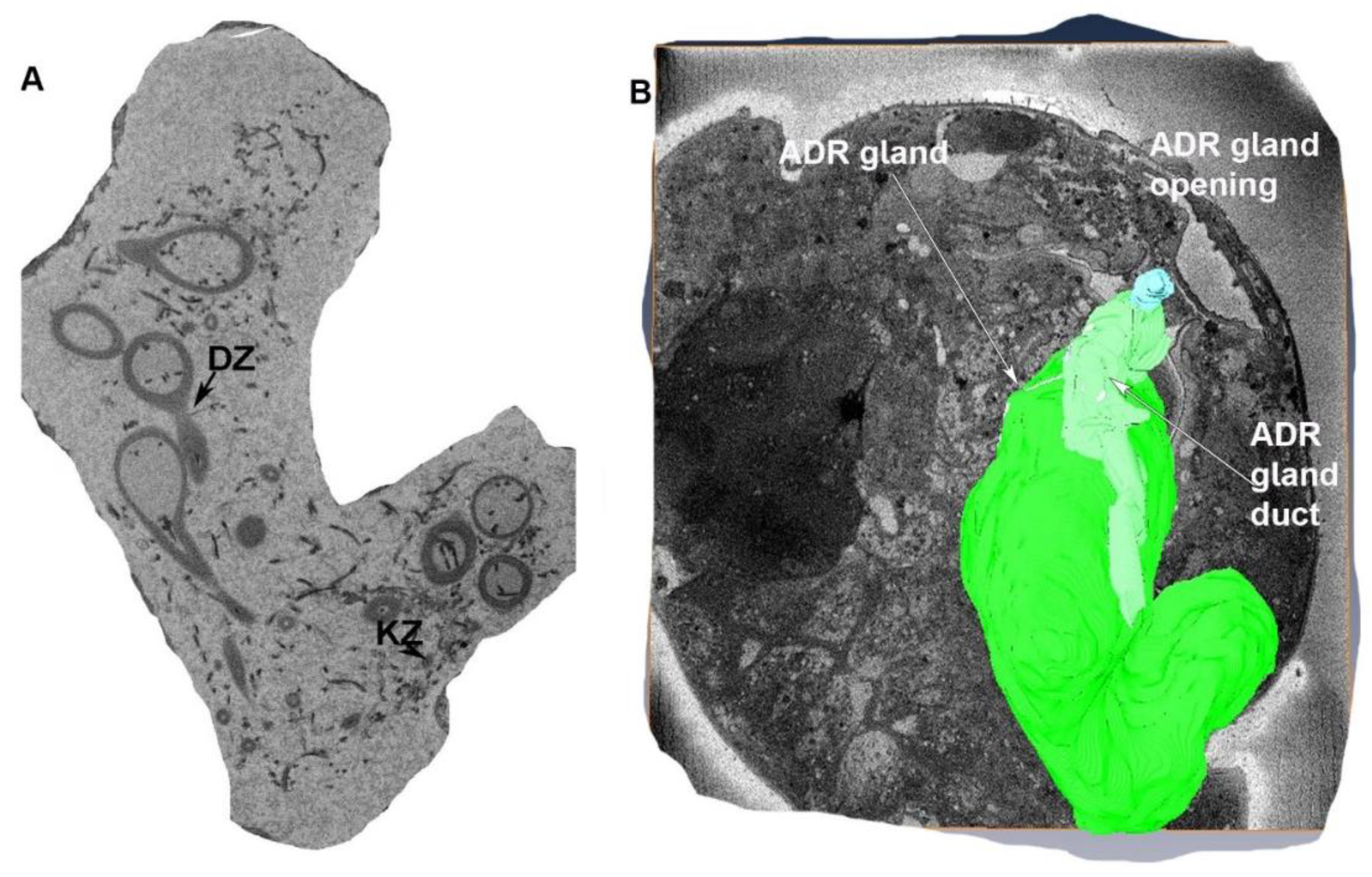
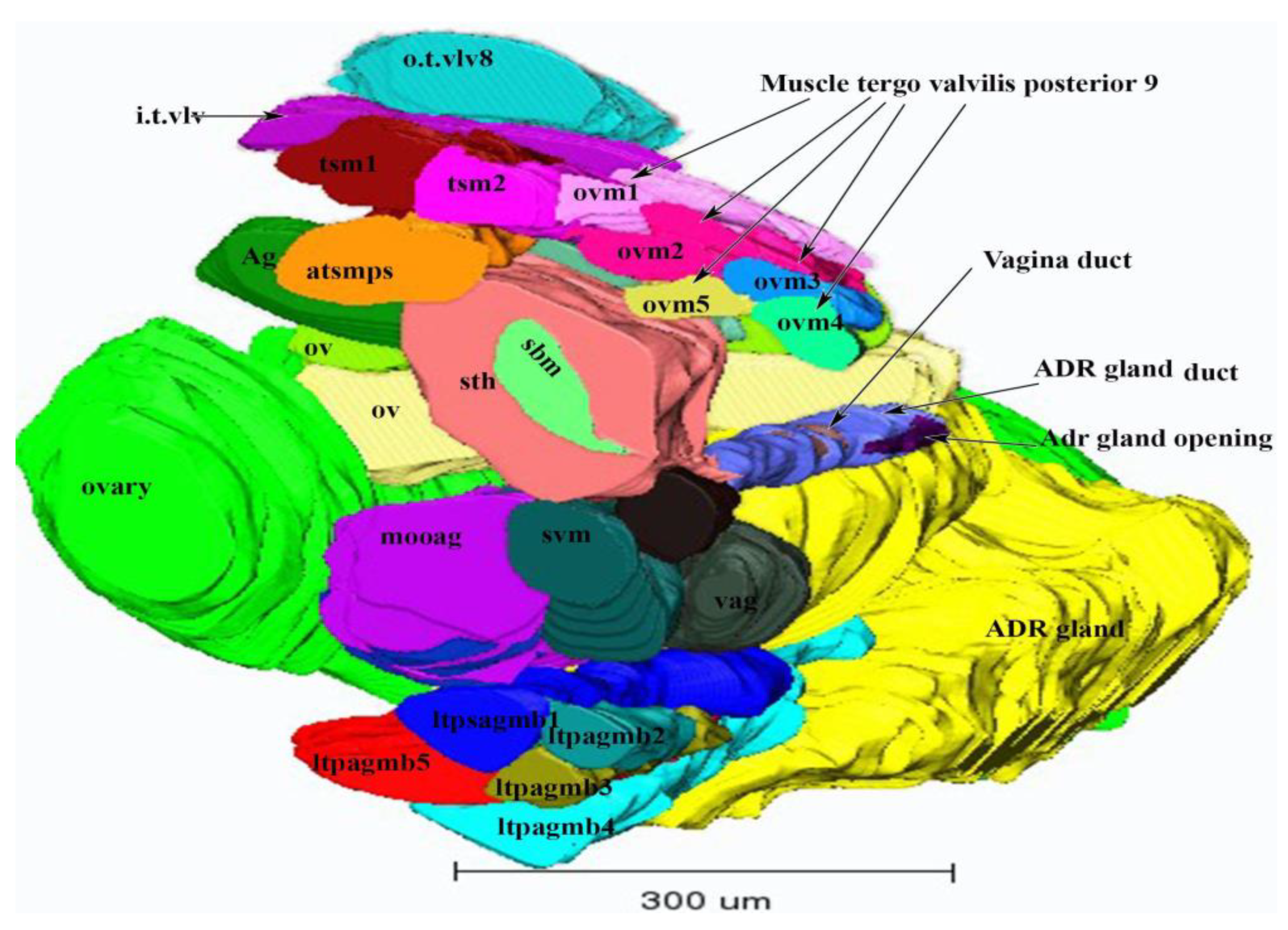
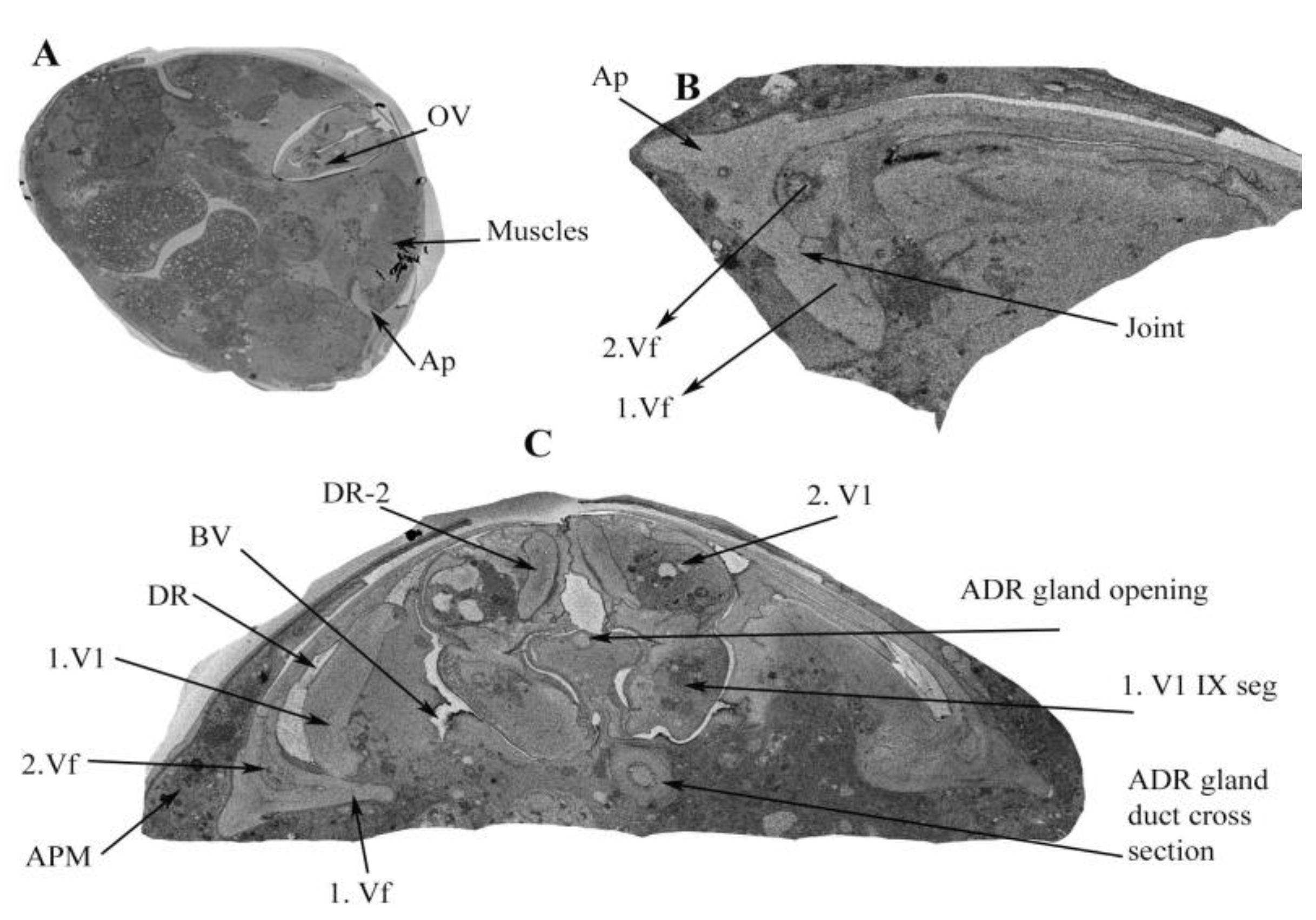
3.1.4. Accessory Glands
3.1.5. The Appendage Gland
3.1.6. Common Oviduct and Vagina
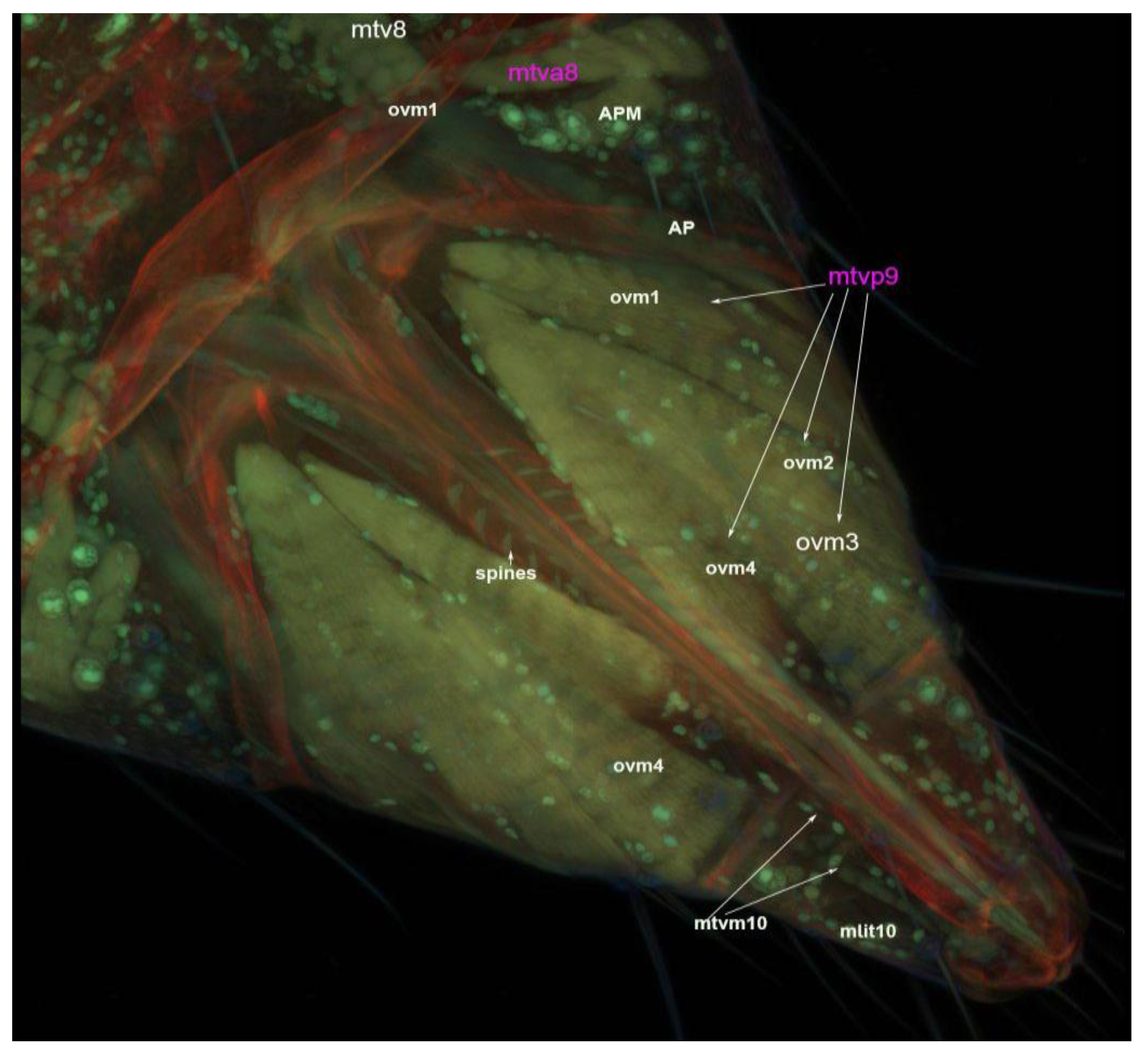
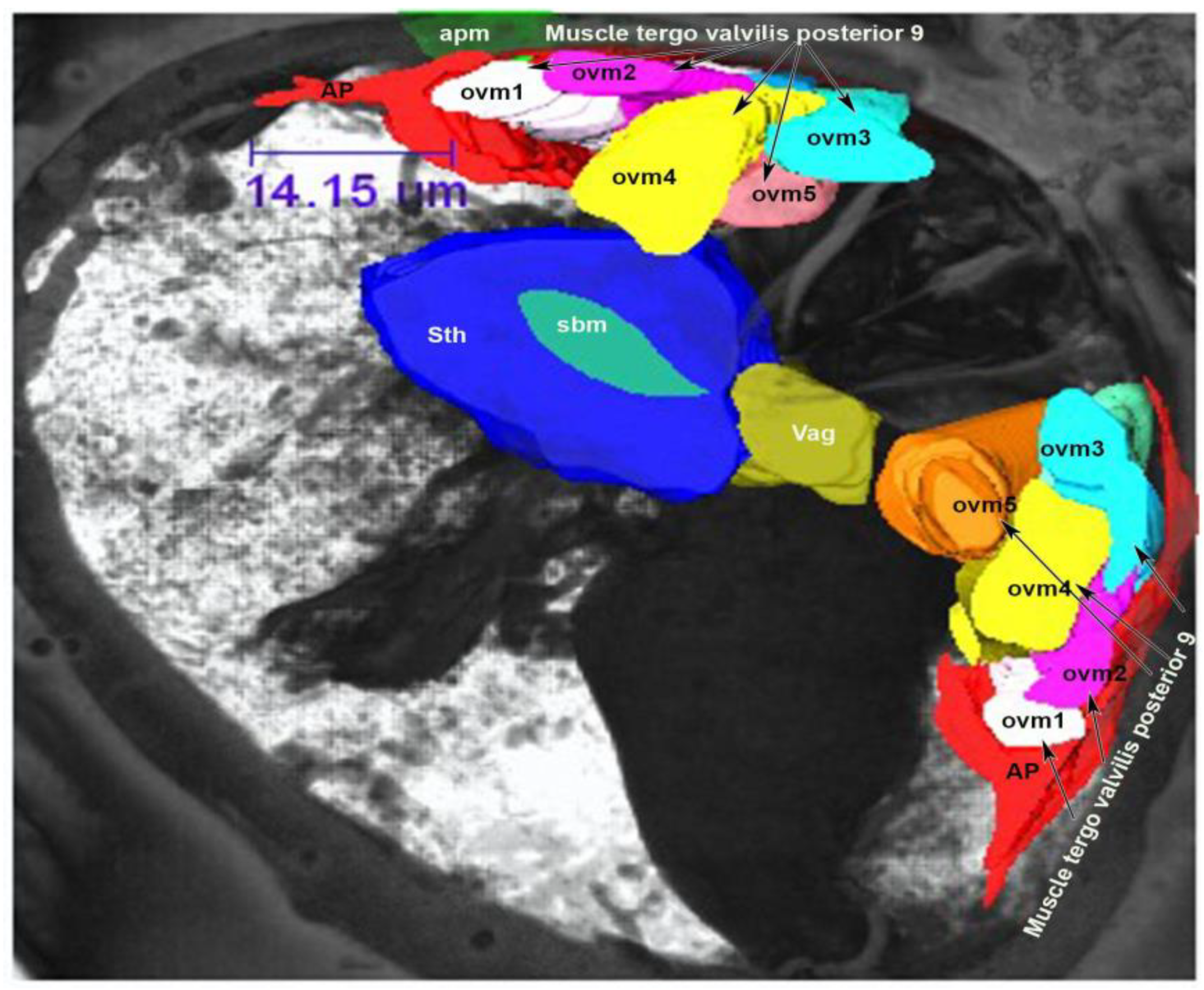
3.2. Muscles of the Internal Genitalia
3.3. External Genitalia
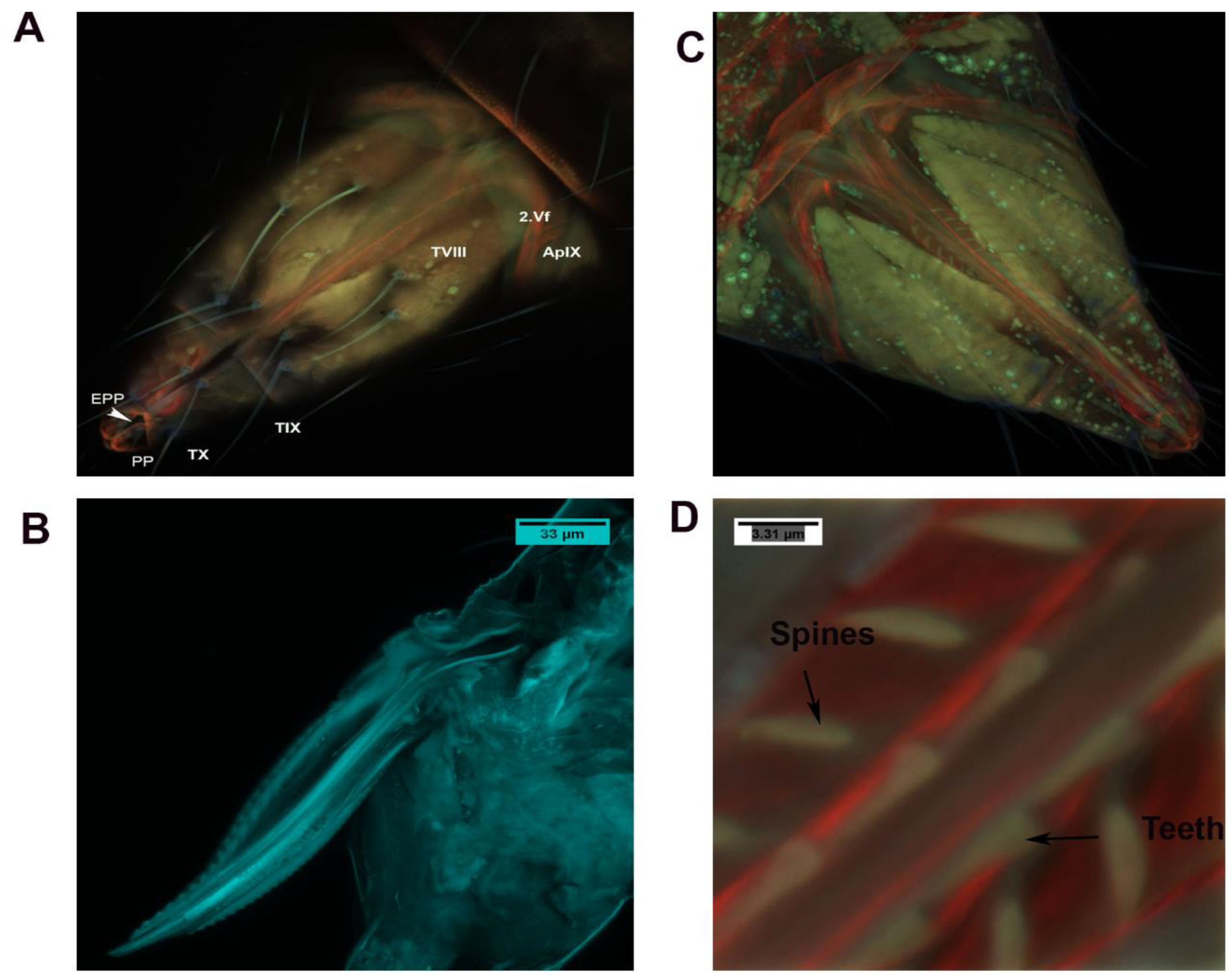
3.3.1. Genital Chamber
3.3.2. Basivalvulae (BV 1 and 2)
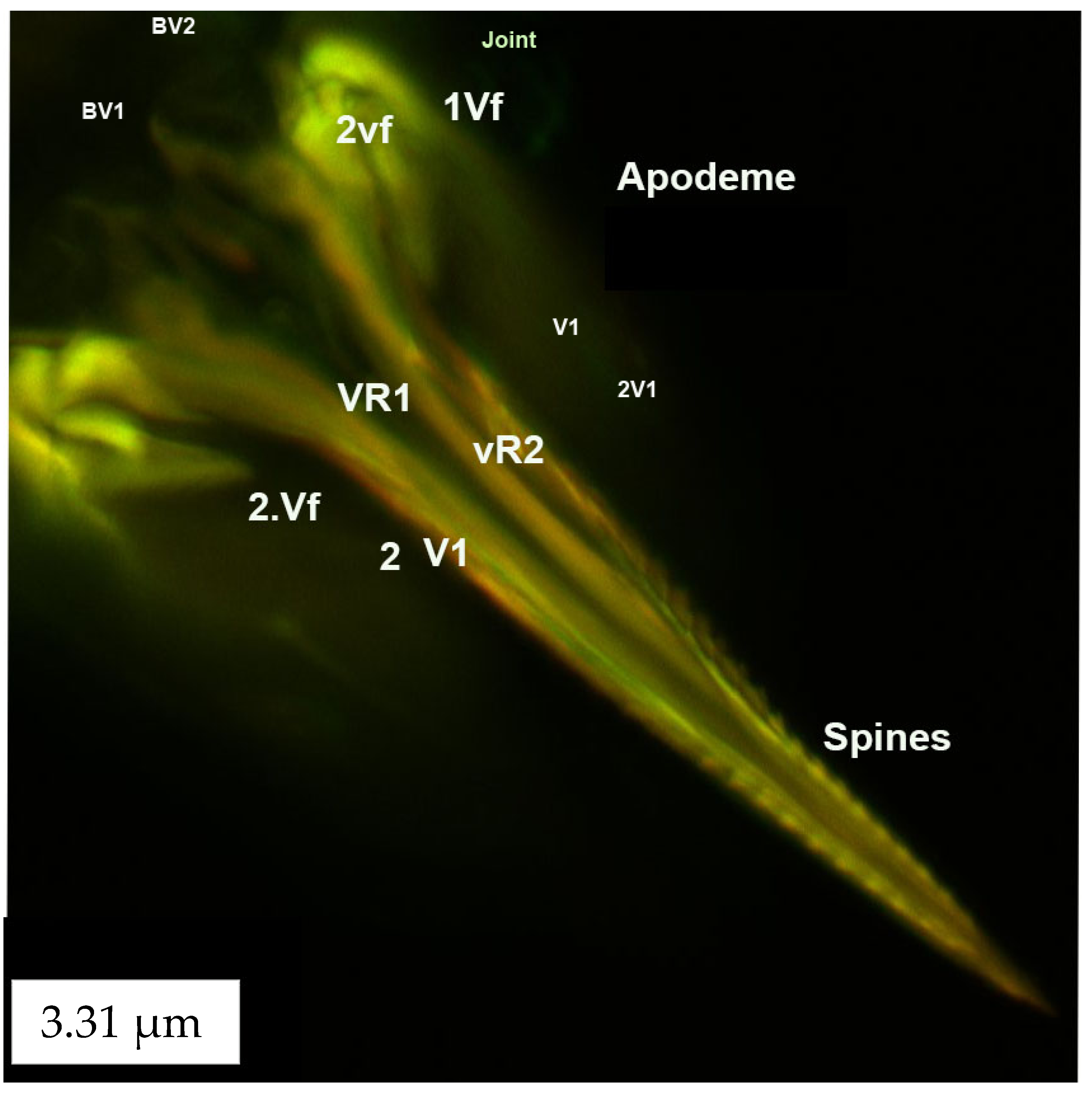
3.3.3. Ventral and Dorsal Ramus
3.4. Muscles of the External Genitalia (Segments VIII, IX and X)
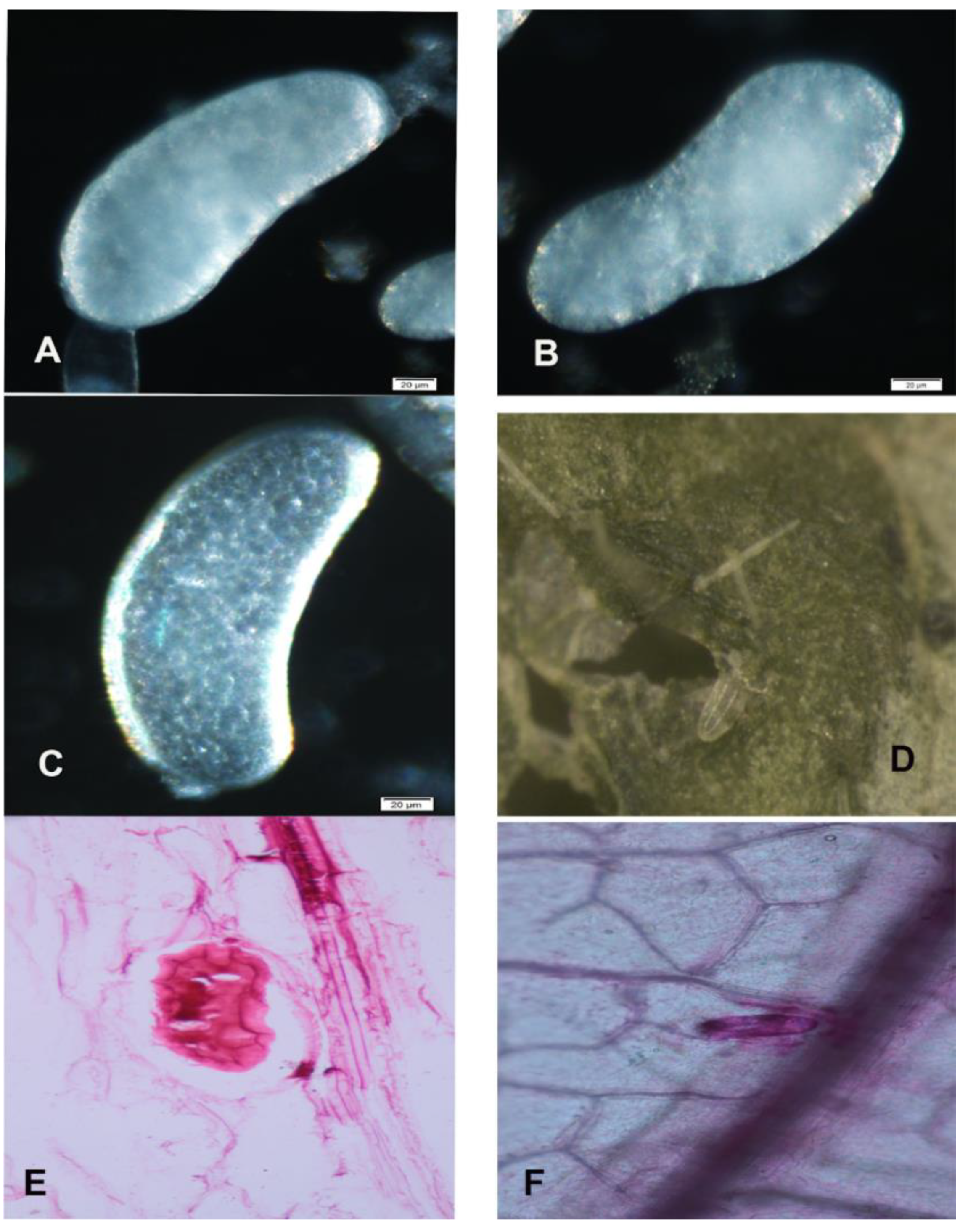
3.5. Morphology of Mature and Immature Eggs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, S.; Tang, L.; Fang, F.; Li, D.; Yuan, X.; Lei, Z.; Gao, Y. Screening, efficacy and mechanisms of microbial control agents against sucking pest insects as thrips. Adv. Insect Physiol. 2018, 55, 199–217. [Google Scholar]
- Kashkouli, M.; Khajehali, J.; Poorjavad, N. Impact of entomopathogenic nematodes on Thrips tabaci Lindeman (Thysanoptera: Thripidae) life stages in the laboratory and under semi-field conditions. J. Biopestic. 2014, 7, 77. [Google Scholar]
- Stuart, R.R.; Gao, Y.-l.; Lei, Z.-R. Thrips: Pests of concern to China and the United States. Agric. Sci. China 2011, 10, 867–892. [Google Scholar] [CrossRef]
- Mound, L.A.; Morris, D.C. The insect order Thysanoptera: Classification versus systematics. Zootaxa 2007, 1668, 395–411. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Kantartzi, S.; Wen, R.-H.; Newman, M.; Hajimorad, M.; Rupe, J.; Tzanetakis, I. Molecular characterization of a new tospovirus infecting soybean. Virus Genes 2011, 43, 289. [Google Scholar] [CrossRef] [PubMed]
- Bloomingdale, C.; Irizarry, M.D.; Groves, R.L.; Mueller, D.S.; Smith, D.L. Seasonal population dynamics of Thrips (Thysanoptera) in Wisconsin and Iowa soybean fields. J. Econ. Entomol. 2016, 110, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, M. Soybean Vein Necrosis Virus: Impacts of Infection on Yield Loss and Seed Quality and Expansion of Plant Host Range. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2016. [Google Scholar]
- Keough, S.A. Surveying Indiana Soybean for Soybean vein necrosis virus (SVNV) and Analysis of SVNV on Life History Traits and Host Preference of Thrips Vectors. Ph.D. Thesis, Purdue University, West Lafayette, IN, USA, 2015. [Google Scholar]
- Bloomingdale, C.; Bradley, C.; Chilvers, M.; Giesler, L.; Groves, R.; Mueller, D.; Smith, D.; Tenuta, A.; Wise, K. An Overview of Soybean Vein Necrosis Virus; Crop Protection Network: Ames, IA, USA, 2015. [Google Scholar] [CrossRef]
- Abd El-Wahab, A.S.; El-Shazly, M.A. Identification and characterization of soybean vein necrosis virus (SVNV): A newly isolated thrips-borne tospovirus in Egypt. J. Virol. Sci. 2017, 1, 76–90. [Google Scholar]
- Abd El-Wahab, A. Molecular characterization and incidence of new tospovirus: Soybean Vein Necrosis Virus (SVNV) in Egypt. Braz. J. Biol. 2021, 84. [Google Scholar] [CrossRef]
- Zhou, J.; Tzanetakis, I.E. Epidemiology of Soybean vein necrosis-associated virus. Phytopathology 2013, 103, 966–971. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Tzanetakis, I.E. Soybean vein necrosis virus: An emerging virus in North America. Virus Genes 2019, 55, 12–21. [Google Scholar] [CrossRef]
- Anderson, N.R.; Irizarry, M.D.; Bloomingdale, C.A.; Smith, D.L.; Bradley, C.A.; Delaney, D.P.; Kleczewski, N.M.; Sikora, E.J.; Mueller, D.S.; Wise, K.A. Effect of soybean vein necrosis on yield and seed quality of soybean. Can. J. Plant Pathol. 2017, 39, 334–341. [Google Scholar] [CrossRef]
- Schoonhoven, A.V.; Pena, J. Estimation of yield losses in cassava following attack from thrips. J. Econ. Entomol. 1976, 69, 514–516. [Google Scholar] [CrossRef]
- Chapman, T.W.; Crespi, B. High relatedness and inbreeding in two species of haplodiploid eusocial thrips (Insecta: Thysanoptera) revealed by microsatellite analysis. Behav. Ecol. Sociobiol. 1998, 43, 301–306. [Google Scholar] [CrossRef]
- Brødsgaard, H.F. Frankliniella occidentalis (Thysanoptera: Thripidae)—A new pest in Danish glasshouses. Tidsskr. Planteavl. 1989, 93, 83–91. [Google Scholar]
- Nault, B.A.; Shelton, A.M.; Gangloff-Kaufmann, J.L.; Clark, M.E.; Werren, J.L.; Cabrera-la Rosa, J.C.; Kennedy, G.G. Reproductive modes in onion thrips (Thysanoptera: Thripidae) populations from New York onion fields. Environ. Entomol. 2006, 35, 1264–1271. [Google Scholar] [CrossRef]
- Leskey, T.C.; Teulon, D.A.; Cameron, E.A. Position and Abundance of Pear Thrips Eggs in Sugar Maple Flowering and Vegetative Buds. In Thrips Biology and Management; Springer: Cham, Switzerland, 1995; pp. 93–95. [Google Scholar]
- Heming, B.S. History of the Germ Line in Male and Female Thrips. In Thrips Biology and Management; Springer: Cham, Switzerland, 1995; pp. 505–535. [Google Scholar]
- Bailey, S.F.; Bailey, S.F. The Thrips of California; University of California Press: Oakland, CA, USA, 1957. [Google Scholar]
- Stannard, L.J. The thrips, or Thysanoptera, of Illinois. Illinois Natural History Survey Bulletin; Illinois Natural History Survey: Champaign, IL, USA, 1968; Volume 29, No. 04. [Google Scholar]
- Tipping, C. Thrips (Thysanoptera). In Encyclopedia of Entomology; Springer: Cham, Switzerland, 2008; Volume 4, pp. 3769–3770. [Google Scholar]
- Mehle, N.; Trdan, S. Traditional and modern methods for the identification of thrips (Thysanoptera) species. J. Pest Sci. 2012, 85, 179–190. [Google Scholar] [CrossRef]
- Johnson, K.P.; Dietrich, C.H.; Friedrich, F.; Beutel, R.G.; Wipfler, B.; Peters, R.S.; Allen, J.M.; Petersen, M.; Donath, A.; Walden, K.K. Phylogenomics and the evolution of hemipteroid insects. Proc. Natl. Acad. Sci. USA 2018, 115, 12775–12780. [Google Scholar] [CrossRef] [Green Version]
- Parrella, M.P. IPM—Approaches and prospects. In Thrips Biology and Management; Springer: Cham, Switzerland, 1995; pp. 357–363. [Google Scholar]
- Jones, T. The external morphology of Chirothrips hamatus (Tryborn) (Thysanoptera). Trans. R. Ent Society London 1954, 105, 163–187. [Google Scholar] [CrossRef]
- Jordan, K. Anatomie und Biologie der Physapoda. Z. Wiss. Zool 1888, 47, 541–620. [Google Scholar]
- Bode, W. Der Ovipositor und die weiblichen geschlechtswege der Thripiden (Thysanoptera, Terebrantia). Z. Morphol. Tiere 1975, 81, 1–53. [Google Scholar] [CrossRef]
- Buckman, R.S.; Mound, L.A.; Whiting, M.F. Phylogeny of thrips (Insecta: Thysanoptera) based on five molecular loci. Syst. Entomol. 2013, 38, 123–133. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Mound, M.L.; Paris, D.L. Thrips of California—Lucid Key Server; CBIT Publishing: Queensland, Australia, 2012; Available online: https://keys.lucidcentral.org/keys/v3/thrips_of_california/Thrips_of_California.html (accessed on 1 June 2022).
- Backus, E.A.; Hunter, W.B.; Arne, C.N. Technique for staining leafhopper (Homoptera: Cicadellidae) salivary sheaths and eggs within unsectioned plant tissue. J. Econ. Entomol. 1988, 81, 1819–1823. [Google Scholar] [CrossRef]
- McBryde, M.C. A method of demonstrating rust hyphae and haustoria in unsectioned leaf tissue. Am. J. Bot. 1936, 23, 686–688. [Google Scholar] [CrossRef]
- Davies, R. The skeletal musculature and its metamorphosis in Limothrips cerealium Haliday (Thysanoptera: Thripidae). Trans. R. Entomol. Soc. Lond. 1969, 121, 167–233. [Google Scholar] [CrossRef]
- Snodgrass, R. Principles of Insect Morphology; McGraw-Hill Book Company: New York, NY, USA, 1935; 667p. [Google Scholar]
- Snodgrass, R.E. Principles of Insect Morphology; Cornell University Press: Ithaca, NY, USA, 2018. [Google Scholar]
- Zhou, Y.; Rédei, D. From lanceolate to plate-like: Gross morphology, terminology, and evolutionary trends of the trichophoran ovipositor. Arthropod Struct. Dev. 2020, 54, 100914. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Deerinck, T.J.; Bushong, E.A.; Thor, A.; Ellisman, M.H. NCMIR methods for 3D EM: A new protocol for preparation of biological specimens for serial block face scanning electron microscopy. Microscopy 2010, 1, 6–8. [Google Scholar]
- Boring, C.A.; Sharkey, M.J.; Nychka, J.A. Structure and functional morphology of the ovipositor of Homolobus truncator (Hymenoptera: Ichneumonoidea: Braconidae). J. Hymenopt. Res. 2009, 18, 1–24. [Google Scholar]
- Melis, A. Tisantteri italiani Genus Melanthrips. Redia 1993, 20, 1–143. [Google Scholar]
- Scudder, G. The comparative morphology of the insect ovipositor. Trans. R. Entomol. Soc. Lond. 1961, 113, 25–40. [Google Scholar] [CrossRef]
- Scudder, G. The female genitalia of the heteroptera: Morphology and bearing on classification 1. Trans. R. Entomol. Soc. Lond. 1959, 111, 405–467. [Google Scholar] [CrossRef]
- Ernst, A.; Miko, I.; Deans, A. Morphology and function of the ovipositor mechanism in Ceraphronoidea (Hymenoptera, Apocrita). J. Hymenopt. Res. 2013, 33, 25. [Google Scholar]
- Matushkina, N.A.; Lambret, P.H. Ovipositor morphology and egg laying behaviour in the dragonfly Lestes macrostigma (Zygoptera: Lestidae). Int. J. Odonatol. 2011, 14, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Quicke, D.; Ficken, L.; Fitton, M. New diagnostic ovipositor characters for doryctine wasps (Hymenoptera, Braconidae). J. Nat. Hist. 1992, 26, 1035–1046. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hameed, A.; Ulmer, J.M.; Miko, I.; Rosa, C.; Rajotte, E.G. Morphology of the Female Reproductive System of the Soybean Thrips, Neohydatothrips variabilis (Beach, 1896) (Thysanoptera: Thripidae). Insects 2022, 13, 566. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070566
Hameed A, Ulmer JM, Miko I, Rosa C, Rajotte EG. Morphology of the Female Reproductive System of the Soybean Thrips, Neohydatothrips variabilis (Beach, 1896) (Thysanoptera: Thripidae). Insects. 2022; 13(7):566. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070566
Chicago/Turabian StyleHameed, Asifa, Jonah M. Ulmer, Istvan Miko, Cristina Rosa, and Edwin G. Rajotte. 2022. "Morphology of the Female Reproductive System of the Soybean Thrips, Neohydatothrips variabilis (Beach, 1896) (Thysanoptera: Thripidae)" Insects 13, no. 7: 566. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13070566