Intraspecific Variation in Female Sex Pheromone of the Codling Moth Cydia pomonella


Abstract
:1. Introduction

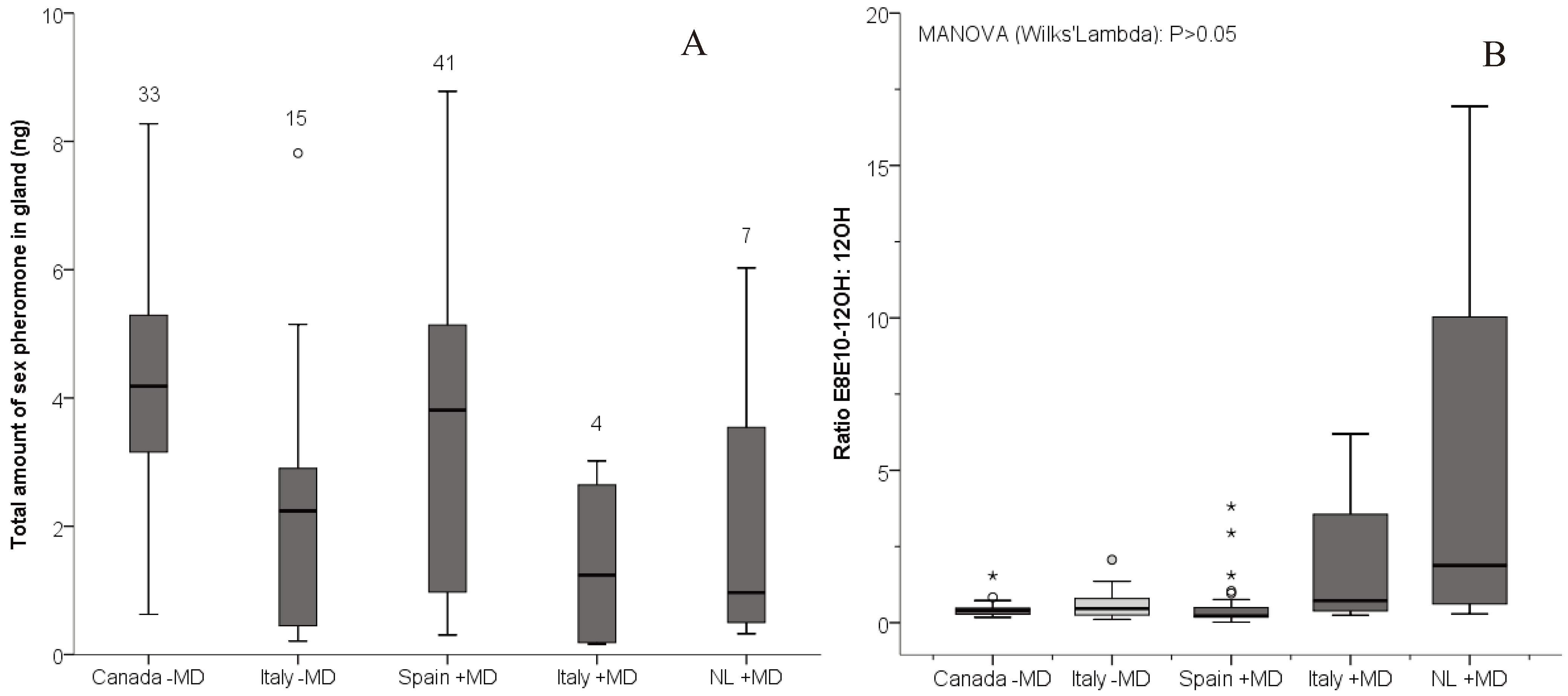{kind=link}
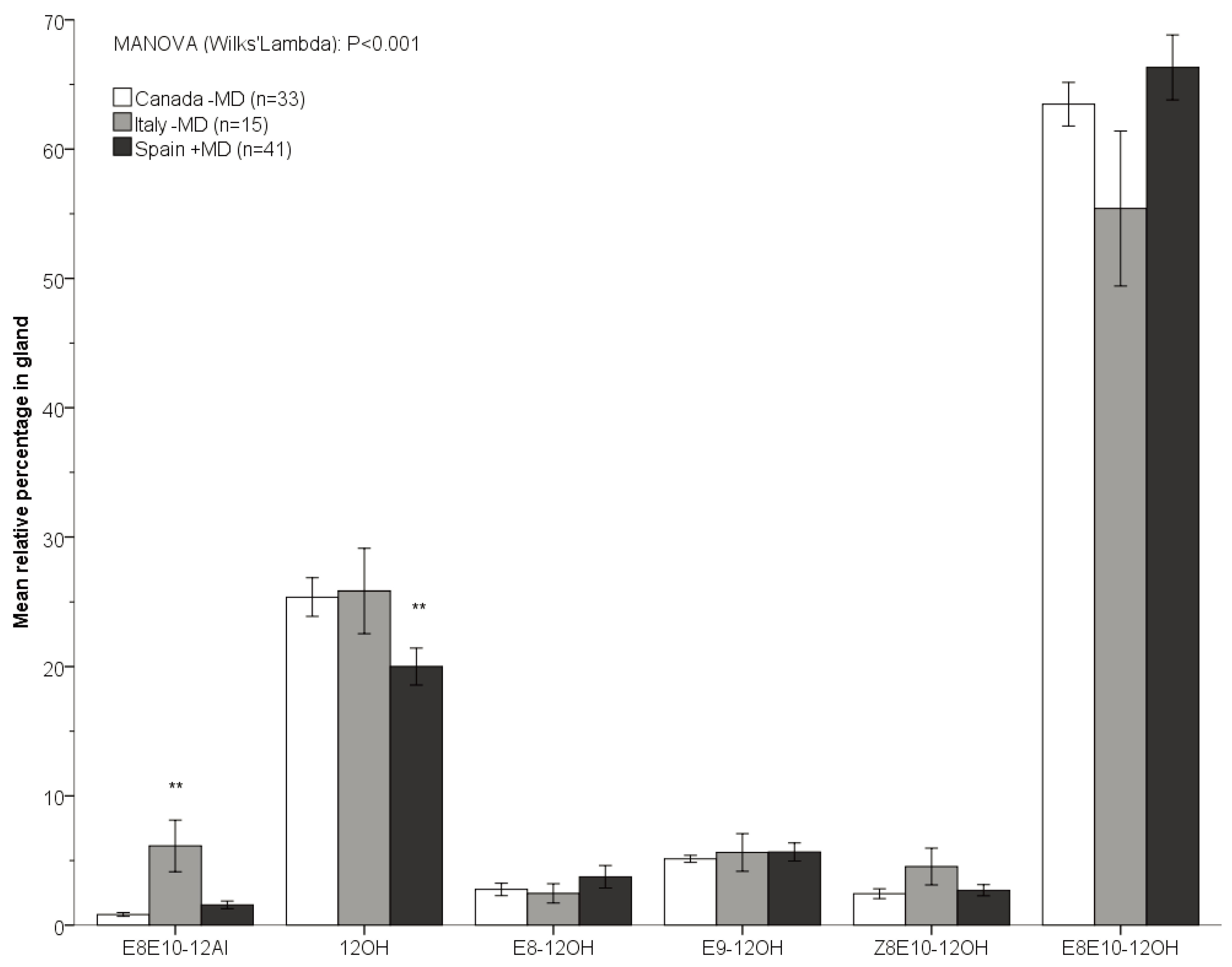{kind=link}
| Compound | Gland Content (ng) | Amount (%) | EAG * | Male Attraction ** WT Field | References | |
|---|---|---|---|---|---|---|
| (1) E8E10-12OH (y) | 2.1 | 100 | +++ | + | + | [29,37,38,40,42] |
| (2) E8E10-12Ac (n) | 0.005 | 0.01 | ++ | − (>1%) + (<1%) | − (>1%) | [40,43,44,54,55,56] |
| (3) 12OH (y) | 1 | 18.4 | + | + | 0 | [29,37,38,39,40,41,50,57] |
| (4) E8-12OH (y) | nd | 0.9 | + | 0 | nd | [40,41] |
| (5) E9-12OH (y) | 0.2 | 5.1 | + | 0 | 0 | [40,41,42,50] |
| (6) E8E10-12Al (y) | 0.02 | 3.9 | + | − | 0 | [40,41] |
| (7) Z8E10-12OH (y) | 0.01 | 0.8 | + | 0 | nd | [40,43,44,54,55] |
| (8) E8Z10-12OH (na) | 0.08 | 1.8 | + | − (>20%) | − | [40,43,44,45,54,55] |
| (9) Z8Z10-12OH (na) | nd | 0.3 | (+) | − (>20%) | nd | [40,43,44,45,54,55] |
| (10) 10OH (y) | 0.005 | 1.4 | 0 | nd | nd | [29,40] |
| (11) 14OH (n) 1 | 0.2 | 3.8 | 0 | nd | 0 | [29,39,40,50] |
| (12) 16OH (na) | 0.04 | 2.6 | 0 | nd | nd | [29,40] |
| (13) 18OH (na) | 0.08 | 3.9 | 0 | nd | nd | [29,40] |
| (14) 18Al (na) | nd | 6.3 | 0 | nd | nd | [29,40] |
| Blend 1 + 3 + 11 | nd | +0 | +0 | [29,38,39,48,58] | ||
| Blend 1 + 3 + 5 + 11 | nd | nd | 0 | [50] | ||
| Blend 1 + 3 + 4 + 5 + 8 + 11 | nd | + | 0 | [40,56] | ||
| Blend 1 + 2 + 8 | nd | nd | − | [40] | ||
| Blend 1 +3 + 5 + 7 + 8 + 10 + 11 + 12 + 13 | nd | 0 | nd | [59] | ||
2. Experimental Section
2.1. Moths
2.2. Gland Extraction
2.3. Gas Chromatography
2.4. Analyses
3. Results

| Source | Sum of Squares | df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Dependent variable: Total amount | |||||
| Mating disruption (+MD, −MD) | 0.102 | 1 | 0.102 | 0.019 | 0.891 |
| Countries | 76.725 | 4 | 19.181 | 3.965 | 0.005 |
| Canada * Spain +MD | 0.632 | ||||
| Canada * Italy −MD | 0.025 | ||||
| Canada * Italy +MD | 0.093 | ||||
| Canada * Neth +MD | 0.135 | ||||
| Spain + MD * Italy −MD | 0.250 | ||||
| Spain + MD * Italy +MD | 0.307 | ||||
| Spain + MD * Neth +MD | 0.499 | ||||
| Neth + MD * Italy −MD | 1.000 | ||||
| Neth + MD * Italy +MD | 0.979 | ||||
| Italy − MD * Italy +MD | |||||
| Dependent variable: Relative amounts | |||||
| Geographic region (Canada −MD, Spain +MD, Italy −MD) | |||||
| E8E10-12Al | 9.787 | 2 | 4.893 | 25.916 | <0.001 |
| Canada * Spain | 0.148 | ||||
| Canada * Italy | <0.001 | ||||
| Spain * Italy | <0.001 | ||||
| 12OH | 0.696 | 2 | 0.348 | 4.170 | 0.018 |
| Canada * Spain | 0.373 | ||||
| Canada * Italy | 0.112 | ||||
| Spain * Italy | 0.007 | ||||
| E8-12OH | 0.053 | 2 | 0.026 | 0.093 | 0.911 |
| Z8E10-12OH | 0.916 | 2 | 0.458 | 2.129 | 0.125 |
| E8E10-12OH | 0.146 | 2 | 0.073 | 0.350 | 0.706 |
| Exposure (+MD (Spain, −MD (Canada and Italy)) | |||||
| E8E10-12Al | 0.274 | 1 | 0.274 | 0.920 | 0.340 |
| 12OH | 0.476 | 1 | 0.476 | 5.603 | 0.020 |
| E8-12OH | 0.161 | 1 | 0.161 | 0.557 | 0.458 |
| Z8E10-12OH | 0.165 | 1 | 0.165 | 0.816 | 0.369 |
| E8E10-12OH | 0.001 | 1 | 0.001 | 0.003 | 0.958 |
| Dependent variable: Ratio to E8E10-12OH | |||||
| Geographic region (Canada −MD, Spain +MD and Italy −MD) | |||||
| 12OH | 0.507 | 2 | 0.254 | 2.176 | 0.120 |
| E8E10-12Al | 8.019 | 2 | 4.009 | 10.585 | <0.001 |
| Canada * Spain | 0.488 | ||||
| Canada * Italy | <0.001 | ||||
| Spain * Italy | 0.001 | ||||
| Exposure (+MD (Spain), −MD (Canada and Italy)) | |||||
| E8E10-12Al | 0.249 | 1 | 0.249 | 0.537 | 0.465 |
| 12OH | 0.443 | 1 | 0.443 | 3.821 | 0.054 |

4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Butlin, R.; Trickett, A.J. Can population genetic simulations help to interpret pheromone evolution? In Insect Pheromone Research: New Direction; Carde, R.T., Minks, A.K., Eds.; Chapman and Hall: New York, NY, USA, 1997; pp. 548–562. [Google Scholar]
- Coyne, J.A.; Orr, H.A. “Patterns of speciation in Drosophila” revisited. Evolution 1997, 51, 295–303. [Google Scholar]
- Löfstedt, C. Moth pheromone genetics and evolution. Phil. Trans. Royal. Soc. Lond. B 1993, 340, 167–177. [Google Scholar]
- Butlin, R.K. Reinforcement—An idea evolving. Trends Ecol. Evol. 1995, 10, 432–434. [Google Scholar]
- Cardé, R.T.; Cardé, A.M.; Hill, A.S.; Roelofs, W.L. Sex pheromone specificity as a reproductive isolating mechanism among sibling species Archips argyrospilus and A. mortuanus and other sympatric tortricine moths (Lepidoptera, Tortricidae). J. Chem. Ecol. 1977, 3, 71–84. [Google Scholar]
- Gries, G.; Schaefer, P.W.; Gries, R.; Liska, J.; Gotoh, T. Reproductive character displacement in Lymantria monacha from northern Japan? J. Chem. Ecol. 2001, 27, 1163–1176. [Google Scholar]
- Groot, A.T.; Horovitz, J.L.; Hamilton, J.; Santangelo, R.G.; Schal, C.; Gould, F. Experimental evidence for interspecific directional selection on moth pheromone communication. Proc. Natl. Acad. Sci. USA 2006, 103, 5858–5863. [Google Scholar]
- Löfstedt, C.; Herrebout, W.M.; Menken, S.B.J. Sex pheromones and their potential role in the evolution of reproductive isolation in small ermine moths (Yponomeutidae). Chemoecol. 1991, 2, 20–28. [Google Scholar]
- McElfresh, J.S.; Millar, J.G. Geographic variation in sex pheromone blend of Hemileuca electra from Southern California. J. Chem. Ecol. 1999, 25, 2505–2525. [Google Scholar]
- McElfresh, J.S.; Millar, J.G. Geographic variation in the pheromone system of the saturniid moth Hemileuca eglanterina. Ecology 2001, 82, 3505–3518. [Google Scholar]
- Eizaguirre, M.; Albajes, R.; Lopez, C.; Sans, A.; Gemeno, C. Inhibition of pheromone response in Sesamia nonagrioides by the pheromone of the sympatric corn borer, Ostrinia nubilalis. Pest Managm. Sci. 2007, 63, 608–614. [Google Scholar]
- Gemeno, C.; Sans, A.; Lopez, C.; Albajes, R.; Eizaguirre, M. Pheromone antagonism in the european corn borer moth Ostrinia nubilalis. J. Chem. Ecol. 2006, 32, 1071–1084. [Google Scholar]
- Haynes, K.F.; Gemeno, C.; Yeargan, K.V.; Millar, J.G.; Johnson, K.M. Aggressive chemical mimicry of moth pheromones by a bolas spider: How does this specialist predator attract more than one species of prey? Chemoecology 2002, 12, 99–105. [Google Scholar]
- Sole, J.; Sans, A.; Riba, M.; Rosa, E.; Bosch, M.P.; Barrot, M.; Palencia, J.; Castella, J.; Guerrero, A. Reduction of damage by the Mediterranean corn borer, Sesamia nonagrioides, and the European corn borer, Ostrinia nubilalis, in maize fields by a trifluoromethyl ketone pheromone analog. Entomol. Exp. Appl. 2008, 126, 28–39. [Google Scholar]
- Cardé, R.T.; Minks, A.K. Control of moth pests by mating disruption—Successes and constraints. Ann. Rev. Entomol 1995, 40, 559–585. [Google Scholar]
- Witzgall, P.; Stelinski, L.; Gutm, L.; Thomson, D. Codling moth management and chemical ecology. Ann. Rev. Entomol. 2008, 53, 503–522. [Google Scholar]
- Cardé, R.T.; Haynes, K.F. Structure of the pheromone communication channel in moths. In Advances in Insect Chemical Ecology; Cardé, R.T., Millar, J.G., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 283–332. [Google Scholar]
- McNeil, J.N.; Delisle, J. The potential of chemical ecology in insect pest management. Phytoprotection 1993, 74, 29–39. [Google Scholar]
- Cardé, R.T. Utilization of pheromones in population management of moth pests. Env. Health Persp. 1976, 14, 133–144. [Google Scholar]
- Evenden, M.L.; Haynes, K.F. Potential for the evolution of resistance to pheromone-based mating disruption tested using two pheromone strains of the cabbage looper, Trichoplusia ni. Entomol. Exp. Appl. 2001, 100, 131–134. [Google Scholar]
- Pearson, G.A.; Dillery, S.; Meyer, J.R. Modeling intra-sexual competition in a sex pheromone system: How much can female movement affect female mating success? J. Theor. Biol. 2004, 231, 549–555. [Google Scholar]
- Mochizuki, F.; Fukumoto, T.; Noguchi, H.; Sugie, H.; Morimoto, T.; Ohtani, K. Resistance to a mating disruptant composed of (Z)-11-tetradecenyl acetate in the smaller tea tortrix, Adoxophyes honmai (Yasuda) (Lepidoptera: Tortricidae). Appl. Entomol. Zool. 2002, 37, 299–304. [Google Scholar]
- Mochizuki, F.; Noguchi, H.; Sugie, H.; Tabata, J.; Kainoh, Y. Sex pheromone communication from a population resistant to mating disruptant of the smaller tea tortrix, Adoxophyes honmai Yasuda (Lepidoptera: Tortricidae). Appl. Entomol. Zool. 2008, 43, 293–298. [Google Scholar]
- Tabata, J.; Noguch, H.; Kainoh, Y.; Mochizuki, F.; Sugie, H. Behavioral response to sex pheromone-component blends in the mating disruption-resistant strain of the smaller tea tortrix, Adoxophyes honmai Yasuda (Lepidoptera: Tortricidae), and its mode of inheritance. Appl. Entomol. Zool. 2007, 42, 675–683. [Google Scholar]
- Tabata, J.; Noguchi, H.; Kainoh, Y.; Mochizuki, F.; Sugie, H. Sex pheromone production and perception in the mating disruption-resistant strain of the smaller tea leafroller moth, Adoxophyes honmai. Entomol. Exp. Appl. 2007, 122, 145–153. [Google Scholar]
- Phelan, P.L. Evolution of sex pheromones and the role of asymmetric tracking. In Insect Chemical Ecology: An Evolutionary Approach; Roitber, B.D., Isman, M.B., Eds.; Chapman and Hall: New York, NY, USA, 1992; pp. 265–314. [Google Scholar]
- Evenden, M.L.; Spohn, B.G.; Moore, A.J.; Preziosi, R.F.; Haynes, K.F. Inheritance and evolution of male response to sex pheromone in Trichoplusia ni (Lepidoptera: Noctuidae). Chemoecology 2002, 12, 53–59. [Google Scholar]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar]
- Arn, H.; Guerin, P.M.; Buser, H.R.; Rauscher, S.; Mani, E. Sex pheromone blend of the codling moth, Cydia pomonella—Evidence for a behavioral role of dodecan-1-ol. Experientia 1985, 41, 1482–1484. [Google Scholar]
- Knight, A.L. Evaluating pheromone emission rate and blend in disrupting sexual communication of codling moth (Lepidoptera: Tortricidae). Environm. Entomol. 1995, 24, 1396–1403. [Google Scholar]
- Howell, J.F.; Knight, A.L.; Unruh, T.R.; Brown, D.F.; Krysan, J.L.; Sell, C.R.; Kirsch, P.A. Control of codling moth in apple and pear with sex pheromone mediate mating disruption. J. Econ. Entomol. 1992, 85, 918–925. [Google Scholar]
- Brown, D.F.; Knight, A.L.; Howell, J.F.; Sell, C.R.; Krysan, J.L.; Weiss, M. Emission characteristics of a polyethylene pheromone dispenser for mating disruption of codling moth (Lepidoptera, Tortricidae). J. Econ. Entomol. 1992, 85, 910–917. [Google Scholar]
- Barnes, M.M.; Millar, J.G.; Kirsch, P.A.; Hawks, D.C. Codling moth (Lepidoptera, Tortricidae) control by dissemination of synthetic female sex pheromone. J. Econ. Entomol. 1992, 85, 1274–1277. [Google Scholar]
- Witzgall, P.; Backman, A.C.; Svensson, M.; Koch, U.; Rama, F.; El-Sayed, A.; Brauchli, J.; Arn, H.; Bengtsson, M.; Lofqvist, J. Behavioral observations of codling moth, Cydia pomonella, in orchards permeated with synthetic pheromone. BioControl 1999, 44, 211–237. [Google Scholar]
- Huang, J.; Gut, L.J.; Miller, J.R. Codling moth, Cydia pomonella, captures in monitoring traps as influenced by proximately to competing female-like- vs. high-releasing pheromone point sources. J. Insect Behav. 2013, 26, 660–666. [Google Scholar]
- Roelofs, W.L.; Carde, R.T. Hydrocarbon sex pheromone in tiger moths (Arctiidae). Science 1971, 171, 684–686. [Google Scholar]
- Einhorn, J.; Beauvais, F.; Gallois, M.; Descoins, C.; Causse, R. Secondary components of the codling moth (Cydia pomonella L) (Lepidoptera, Tortricidae) sex pheromone. C. R. Acad. Sci. III Sci. Vie 1984, 299, 773–792. [Google Scholar]
- Einhorn, J.; Witzgall, P.; Audemard, H.; Boniface, B.; Causse, R. Secondary components of the codling moth (Cydia pomonella) (Lepidoptera, Tortricidae) sex pheromone. 2. 1st results on the behavioral effects. C. R. Acad. Sci. III Sci. Vie 1986, 302, 263–266. [Google Scholar]
- Bartell, R.J.; Bellas, T.E.; Whittle, C.P. Evidence for biological activity of 2 further alcohols in the sex pheromone of female Cydia pomonella (L) (Lepidoptera, Tortricidae). J. Austr. Entomol. Soc. 1988, 27, 11–12. [Google Scholar]
- Witzgall, P.; Bengtsson, M.; Rauscher, S.; Liblikas, I.; Backman, A.C.; Coracini, M.; Anderson, P.; Lofqvist, J. Identification of further sex pheromone synergists in the codling moth, Cydia pomonella. Entomol. Exp. Appl. 2001, 101, 131–141. [Google Scholar]
- Ebbinghaus, D.; Losel, P.M.; Lindemann, M.; Scherkenbeck, J.; Zebitz, C.P.W. Detection of major and minor sex pheromone components by the male codling moth Cydia pomonella (Lepidoptera: Tortricidae). J. Insect Physiol. 1998, 44, 49–58. [Google Scholar]
- Roelofs, W.; Comeau, A.; Hill, A.; Milicevic, G. Sex attractant of the codling moth: Characterization with electroantennogram technique. Science 1971, 174, 297–299. [Google Scholar]
- Hathaway, D.O.; McGovern, T.P.; Beroza, M.; Moffitt, H.R.; McDonough, L.M.; Buit, B.A. An inhibitor of sexual attraction of male codling moths to a synthetic sex pheromone and virgin females in traps. Environm. Entomol. 1974, 3, 522–524. [Google Scholar]
- George, D.A.; McDonough, L.M.; Hathaway, D.O.; Moffitt, H.R. Inhibitors of sexual attraction of male codling moths. Environm. Entomol. 1975, 4, 606–608. [Google Scholar]
- Backman, A.C.; Anderson, P.; Bengtsson, M.; Lofqvist, J.; Unelius, C.R.; Witzgall, P. Antennal response of codling moth males, Cydia pomonella L. (Lepidoptera: Tortricidae), to the geometric isomers of codlemone and codlemone acetate. J. Comp. Physiol. A 2000, 186, 513–519. [Google Scholar]
- El-Sayed, A.M. Behavioural effect of (E)-8,(Z)-10-dodecadien-1-OL and (E)-8,(E)-10-dodecadienyl acetate on the upwind orientation of male codling moth, Cydia pomonella to pheromone source. Behaviour 2004, 141, 313–325. [Google Scholar]
- Rothschild, G.H.L. Mating disruption of lepidopterous pests: Current status and future prospects. In Management of Insect Pests with Semiochemicals: Concepts and Practice; Mitchell, E.R., Ed.; Plenum Press: New York, NY, USA, 1981; pp. 207–228. [Google Scholar]
- McDonough, L.M.; Davis, H.G.; Chapman, P.S.; Smithhisler, C.L. Response of male codling moths (Cydia pomonella) to componens of conspecific female sex pheromone glands in flight tunnel tests. J. Chem. Ecol. 1993, 19, 1737–1748. [Google Scholar]
- El-Sayed, A.; Liblikas, I.; Unelius, R. Flight and molecular modeling study on the response of codling moth, Cydia pomonella (Lepidoptera: Tortricidae) to (E,E)-8,10-dodecadien-1-ol and its geometrical isomers. Z. Naturforsch. C 2000, 55, 1011–1017. [Google Scholar]
- Bäckman, A.C.; Bengtsson, M.; Witzgall, P. Pheromone release by individual females of codling moth, Cydia pomonella. J. Chem. Ecol. 1997, 23, 807–815. [Google Scholar]
- Svensson, M.G.E.; Bengtsson, M.; Lofqvist, J. Individual variation and repeatability of sex pheromone emission of female turnip moths Agrotis segetum. J. Chem. Ecol. 1997, 23, 1833–1850. [Google Scholar]
- Du, J.W.; Lofstedt, C.; Lofqvist, J. Repeatability of pheromone emissions from individual female ermine moths Yponomeuta padellus and Yponomeuta rorellus. J. Chem. Ecol. 1987, 13, 1431–1441. [Google Scholar]
- Miklas, N.; Renou, M.; Malosse, I.; Malosse, C. Repeatability of pheromone blend composition in individual males of the southern green stink bug, Nezara viridula. J. Chem. Ecol. 2000, 26, 2473–2485. [Google Scholar]
- Witzgall, P.; Backman, A.C.; Svensson, M.; Bengtsson, M.; Unelius, C.R.; Vrkoc, J.; Kirsch, P.A.; Ioriatti, C.; Lofqvist, J. Potential of a blend of E8,E10-120H and E8,E10-12Ac for mating disruption of codling moth, Cydia pomonella L. (Lep, Tortricidae). J. Appl. Entomol. 1996, 120, 611–614. [Google Scholar]
- El-Sayed, A.; Unelius, R.C.; Liblikas, I.; Lofqvist, J.; Bengtsson, M.; Witzgall, P. Effect of codlemone isomers on codling moth (Lepidoptera: Tortricidae) male attraction. Env. Entomol. 1998, 27, 1250–1254. [Google Scholar]
- El-Sayed, A.; Bengtsson, M.; Rauscher, S.; Lofqvist, J.; Witzgall, P. Multicomponent sex pheromone in codling moth (Lepidoptera: Tortricidae). Environm. Entomol. 1999, 28, 775–779. [Google Scholar]
- Causse, R.; Barthes, J.; Witzgall, P.; Einhorn, J. Secondary components of the codling moth Cydia pomonella L. (Lepidoptera, Tortricidae) sex pheromone. 3. Synergistic properties of dodecanol in field trapping. C. R. Acad. Sci. III Sci. Vie 1988, 306, 125–128. [Google Scholar]
- McDonough, L.M.; Davis, H.G.; Chapman, P.S.; Smithhisler, C.L. Codling moth (Cydia pomonella)—Disruptants of sex pheromonal communication. J. Chem. Ecol. 1994, 20, 171–181. [Google Scholar]
- McDonough, L.M.; Davis, H.G.; Chapman, P.S.; Smithhisler, C.L. Codling moth, Cydia pomonella, (Lepidoptera, Torticidae)—Is its sex pheromone multicomponent? J. Chem. Ecol. 1995, 21, 1065–1071. [Google Scholar]
- The Pherobase: Database of insect pheromones and semiochemicals. Available online: http://www.pherobase.com (accessed on 3 February 2014).
- ChemStation Software, Agilent Technologies: Santa Clara, CA, USA, 2010.
- Groot, A.T.; Classen, A.; Staudacher, H.; Schal, C.; Heckel, D.G. Phenotypic plasticity in sexual communication signal of a noctuid moth. J. Evol. Biol. 2010, 23, 2731–2738. [Google Scholar]
- IBM Corporation. IBM SPSS Statistics for Windows, 20.0 ed. IBM Corporation: Armonk, NY, USA, 2011.
- Harari, A.R.; Zahavi, T.; Thiery, D. Fitness cost of pheromone production in signaling female moths. Evolution 2011, 65, 1572–1582. [Google Scholar]
- Symonds, M.R.E.; Johnson, T.L.; Elgar, M.A. Pheromone production, male abundance, body size, and the evolution of elaborate antennae in moths. Ecol. Evol. 2012, 2, 227–246. [Google Scholar]
- Ansebo, L.; Ignell, R.; Lofqvist, J.; Hansson, B.S. Responses to sex pheromone and plant odours by olfactory receptor neurons housed in sensilla auricillica of the codling moth, Cydia pomonella (Lepidoptera: Tortricidae). J. Insect Physiol. 2005, 51, 1066–1074. [Google Scholar]
- Yang, Z.H.; Bengtsson, M.; Witzgall, P. Host plant volatiles synergize response to sex pheromone in codling moth, Cydia pomonella. J. Chem. Ecol. 2004, 30, 619–629. [Google Scholar]
- Weissling, T.J.; Knight, A.L. Oviposition and calling behavior of codling moth (Lepidoptera: Tortricidae) in the presence of codlemone. Ann. Entomol. Soc. Am. 1996, 89, 142–147. [Google Scholar]
- Noguchi, H.; Tamaki, Y. Conspecific female sex pheromone delays calling behavior of Adoxophyes sp and Homon magnanima (Lepidoptera, Tortricidae). Jpn. J. Appl. Entomol. Zool. 1985, 29, 113–118. [Google Scholar]
- Groot, A.T.; Schofl, G.; Inglis, O.; Donnerhacke, S.; Classen, A.; Schmalz, A.; Santangelo, R.; Emerson, J.; Gould, F.; Schal, C.; et al. Within-population variability in a moth sex pheromone blend: genetic basis and behavioral consequences. Proc. Biol. Sci. 2014, 281, 20133054. [Google Scholar]
- Crispo, E. Modifying effects of phenotypic plasticity on interactions among natural selection, adaptation and gene flow. J. Evol. Biol. 2008, 21, 1460–1469. [Google Scholar]
- Price, T.D.; Qvarnstrom, A.; Irwin, D.E. The role of phenotypic plasticity in driving genetic evolution. Proc. Royal. Soc. Lond. B 2003, 270, 1433–1440. [Google Scholar]
- Timm, A.E.; Geertsema, H.; Warnich, L. Gene flow among Cydia pomonella (Lepidoptera: Tortricidae) geographic and host populations in South Africa. J. Econ. Entomol. 2006, 99, 341–348. [Google Scholar]
- Franck, P.; Timm, A.E. Population genetic structure of Cydia pomonella: A review and case study comparing spatiotemporal variation. J. Appl. Entomol. 2010, 134, 191–200. [Google Scholar]
- Chen, M.H.; Dorn, S. Microsatellites reveal genetic differentiation among populations in an insect species with high genetic variability in dispersal, the codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae). Bull. Entomol. Res. 2010, 100, 75–85. [Google Scholar]
- Men, Q.-L.; Chen, M.-H.; Zhang, Y.-L.; Feng, J.-N. Genetic structure and diversity of a newly invasive species, the codling moth, Cydia pomonella (L.) (Lepidoptera: Tortricidae) in China. Biol. Invasions 2013, 15, 447–458. [Google Scholar]
- Fuentes-Contreras, E.; Espinoza, J.L.; Lavandero, B.; Ramirez, C.C. Population genetic structure of codling moth (Lepidoptera: Tortricidae) from apple orchards in central Chile. J. Econ. Entomol. 2008, 101, 190–198. [Google Scholar]
- Timm, A.E.; Geertsema, H.; Warnich, L. Population genetic structure of economically important Tortricidae (Lepidoptera) in South Africa: A comparative analysis. Bull. Entomol. Res. 2010, 100, 421–431. [Google Scholar]
- Margaritopoulos, J.T.; Voudouris, C.C.; Olivares, J.; Sauphanor, B.; Mamuris, Z.; Tsitsipis, J.A.; Franck, P. Dispersal ability in codling moth: Mark-release-recapture experiments and kinship analysis. Agricult. Forest Entomol. 2012, 14, 399–407. [Google Scholar]
- Voudouris, C.C.; Franck, P.; Olivares, J.; Sauphanor, B.; Mamuris, Z.; Tsitsipis, J.A.; Margaritopoulos, J.T. Comparing the genetic structure of codling moth Cydia pomonella (L.) from Greece and France: Long distance gene-flow in a sedentary pest species. Bull. Entomol. Res. 2012, 102, 185–198. [Google Scholar]
- Basoalto, E.; Miranda, M.; Knight, A.L.; Fuentes-Contreras, E. Landscape analysis of adult codling moth (Lepidoptera: Tortricidae) distribution and dispersal within typical agroecosystems dominated by apple production in central Chile. Environm. Entomol. 2010, 39, 1399–1408. [Google Scholar]
- Light, D.M.; Knight, A.L.; Henrick, C.A.; Rajapaska, D.; Lingren, B.; Dickens, J.C.; Reynolds, K.M.; Buttery, R.G.; Merrill, G.; Roitman, J.; et al. A pear-derived kairomone with pheromonal potency that attracts male and female codling moth, Cydia pomonella (L.). Naturwissenschaften 2001, 88, 333–338. [Google Scholar]
- El-Sayed, A.M.; Cole, L.; Revell, J.; Manning, L.-A.; Twidle, A.; Knight, A.L.; Bus, V.G.M.; Suckling, D.M. Apple volatiles synergize the response of codling moth to pear ester. J. Chem. Ecol. 2013, 39, 643–652. [Google Scholar]
- Knight, A.L.; Light, D.M. Combined approaches using sex pheromone and pear ester for behavioural disruption of codling moth (Lepidoptera: Tortricidae). J. Appl. Entomol. 2014, 138, 96–108. [Google Scholar]
- Trimble, R.M.; El-Sayed, A.M. Potential of ethyl (2E,4Z)-2,4-decadienoate for monitoring activity of codling moth (Lepidoptera: Tortricidae) in eastern North American apple orchards. Can. Entomol. 2005, 137, 110–116. [Google Scholar]
- Mitchell, V.J.; Manning, L.-A.; Cole, L.; Suckling, D.M.; El-Sayed, A.M. Efficacy of the pear ester as a monitoring tool for coming moth Cydia pomonella (Lepidoptera: Tortricidae) in New Zealand apple orchards. Pest Manag. Sci. 2008, 64, 209–214. [Google Scholar]
- Groot, A.T.; Inglis, O.; Bowdrigde, S.; Santangelo, R.G.; Blanco, C.; Lopez, J.D., Jr.; Vargas, A.T.; Gould, F.; Schal, C. Geographic and temporal variation in moth chemical communication. Evolution 2009, 63, 1987–2003. [Google Scholar]
- Svensson, G.P.; Ryne, C.; Lofstedt, C. Heritable variation of sex pheromone composition and the potential for evolution of resistance to pheromone-based control of the Indian meal moth, Plodia interpunctella. J. Chem. Ecol. 2002, 28, 1447–1461. [Google Scholar]
- Xu, J.; Huigens, M.E.; Orr, D.; Groot, A.T. Differential response of Trichogramma wasps to extreme sex pheromone types of the noctuid moth Heliothis virescens. Ecol. Entomol. 2014, 39, 627–636. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duménil, C.; Judd, G.J.R.; Bosch, D.; Baldessari, M.; Gemeno, C.; Groot, A.T. Intraspecific Variation in Female Sex Pheromone of the Codling Moth Cydia pomonella. Insects 2014, 5, 705-721. https://0-doi-org.brum.beds.ac.uk/10.3390/insects5040705
Duménil C, Judd GJR, Bosch D, Baldessari M, Gemeno C, Groot AT. Intraspecific Variation in Female Sex Pheromone of the Codling Moth Cydia pomonella. Insects. 2014; 5(4):705-721. https://0-doi-org.brum.beds.ac.uk/10.3390/insects5040705
Chicago/Turabian StyleDuménil, Claire, Gary J. R. Judd, Dolors Bosch, Mario Baldessari, César Gemeno, and Astrid T. Groot. 2014. "Intraspecific Variation in Female Sex Pheromone of the Codling Moth Cydia pomonella" Insects 5, no. 4: 705-721. https://0-doi-org.brum.beds.ac.uk/10.3390/insects5040705