Evaluation of a Push-Pull System for the Management of Frankliniella Species (Thysanoptera: Thripidae) in Tomato

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plot Establishment and Maintenance
2.2. Insect Sampling
2.3. Statistical Analyses
3. Results
3.1. Abundance of Thrips and O. insidiosus in B. alba
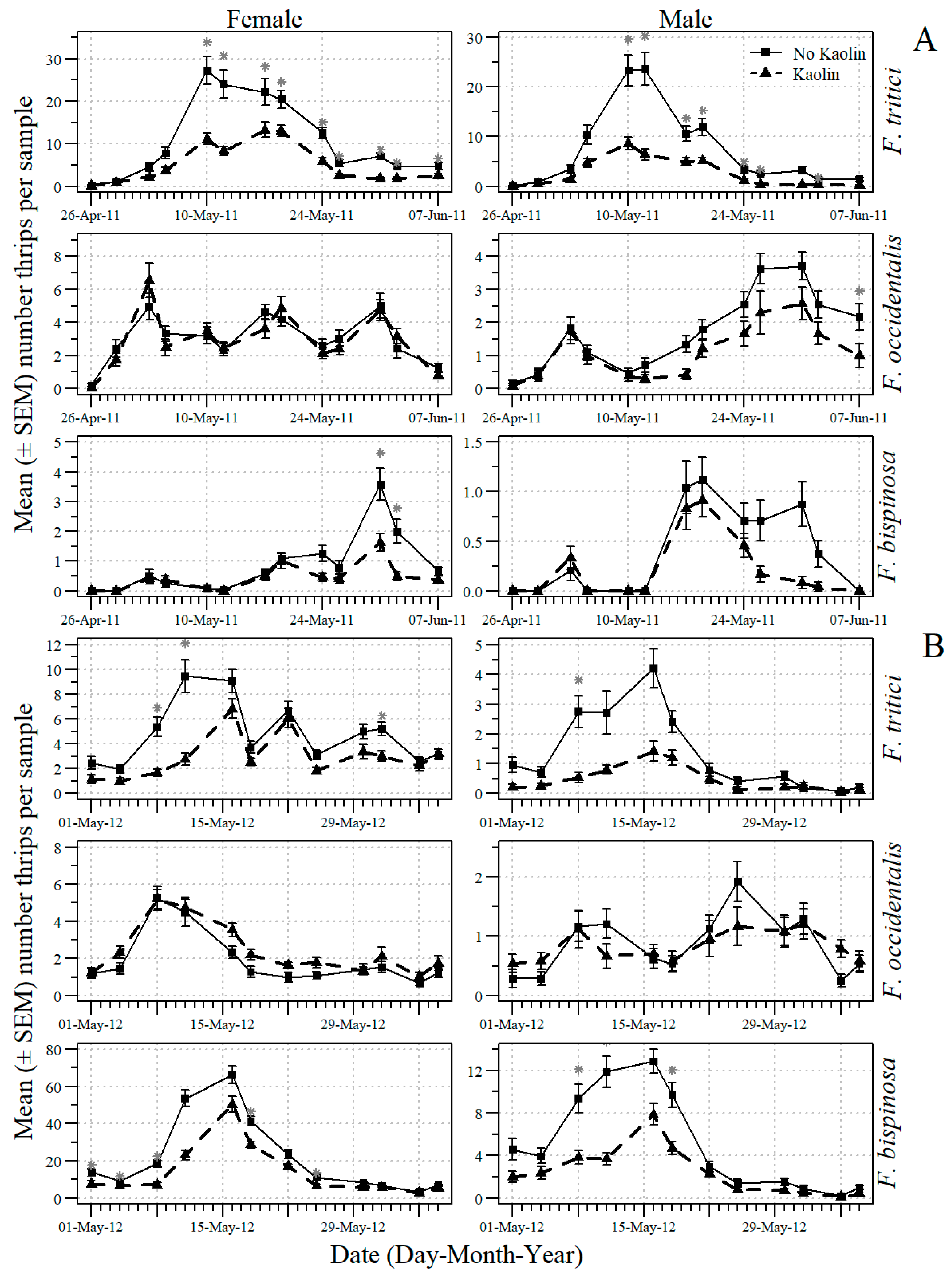
3.2. Effects of Mulch and Kaolin on Thrips and O. insidiosus in B. alba
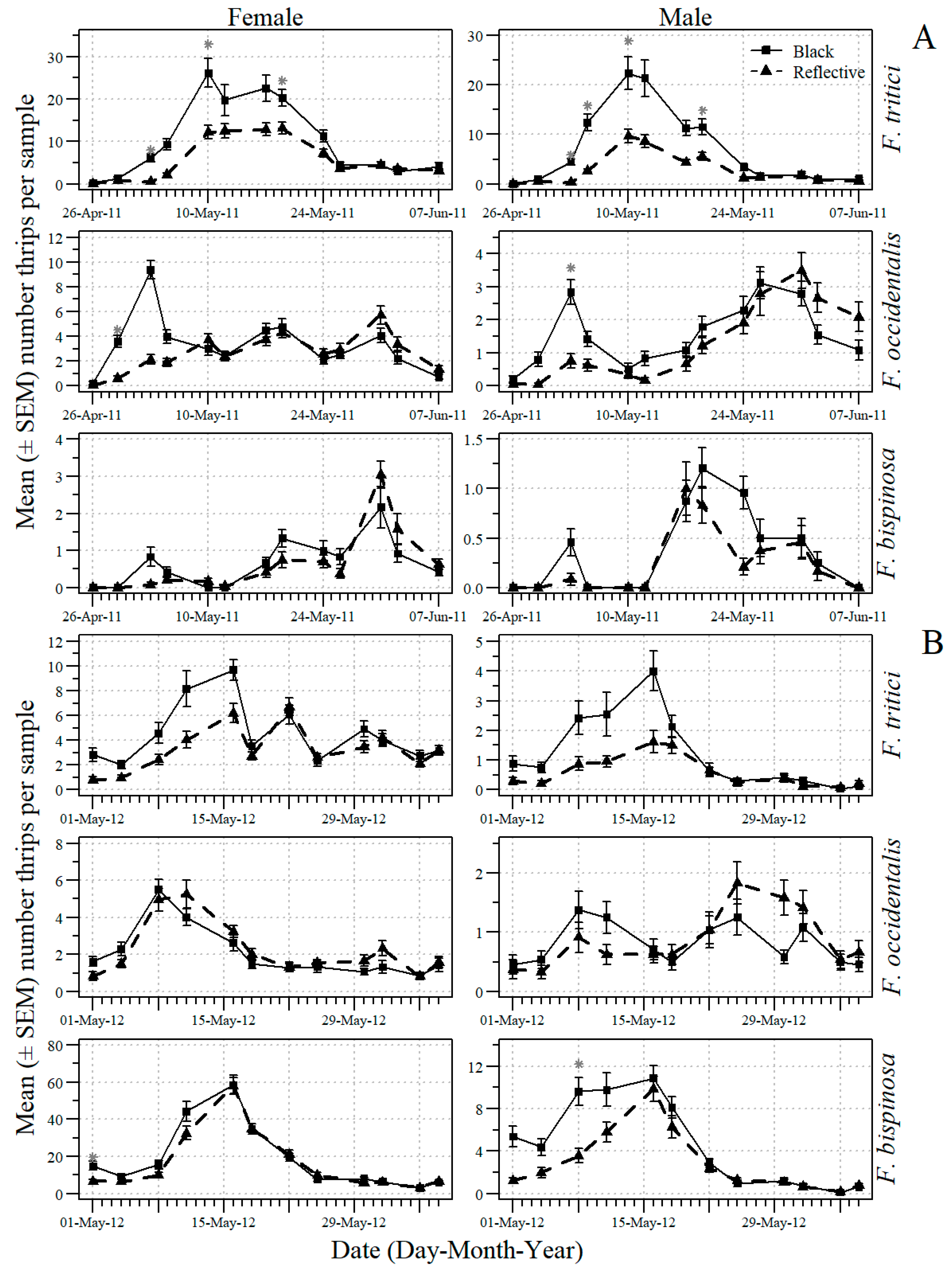
3.3. Effects of Mulch and Kaolin on Thrips and O. insidiosus in Tomato
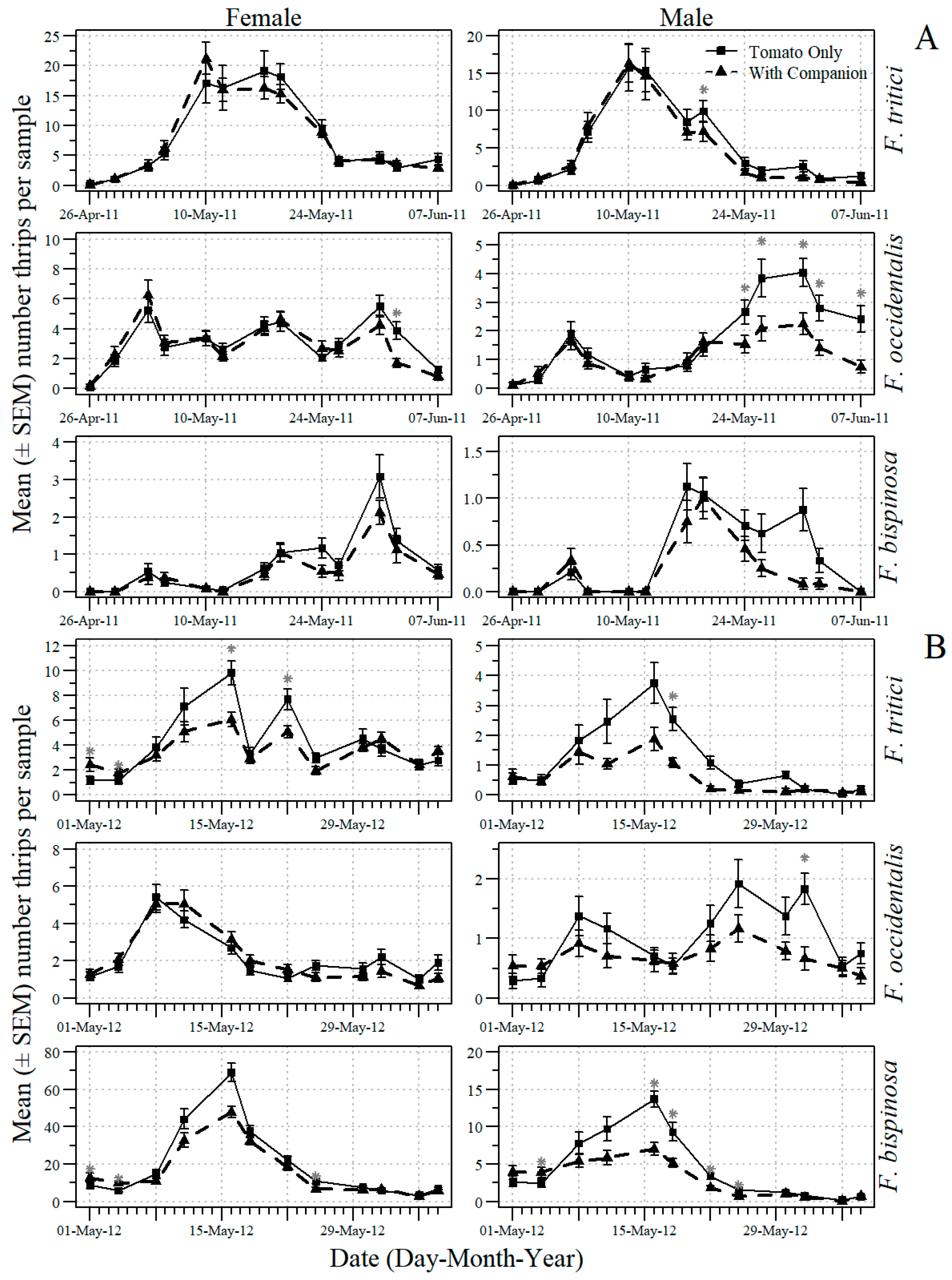
3.4. Effects of Companion Plants on Thrips and O. insidiosus in Tomato
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Salguero-Navas, V.E.; Funderburk, J.E.; Olson, S.M.; Beshear, R.J. Damage to tomato fruit caused by the western flower thrips (Thysanoptera: Thripidae). J. Entomol. Sci. 1991, 26, 436–442. [Google Scholar] [CrossRef]
- Ghidiu, G.M.; Hitchner, E.M.; Funderburk, J.E. Goldfleck damage to tomato fruit caused by feeding of Frankliniella occidentalis (Thysanoptera: Thripidae). Fla. Entomol. 2006, 89, 279–282. [Google Scholar] [CrossRef]
- Webster, C.G.; Frantz, G.; Reitz, S.R.; Funderburk, J.E.; Mellinger, H.C.; McAvoy, E.; Turechek, W.W.; Marshall, S.H.; Tantiwanich, Y.; McGrath, M.T.; et al. Emergence of Groundnut ringspot virus and Tomato chlorotic spot virus in vegetables in Florida and the southeastern United States. Phytopathology 2015, 105, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Demirozer, O.; Tyler-Julian, K.; Funderburk, J.; Leppla, N.; Reitz, S. Frankliniella occidentalis (Pergande) integrated pest management programs for fruiting vegetables in Florida. Pest Manag. Sci. 2012, 68, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paini, D.; Funderburk, J.E.; Reitz, S.R. Competitive exclusion of a worldwide invasive pest by a native: Quantifying competition between two phytophagous insects on two host plant species. J. Anim. Ecol. 2008, 77, 184–190. [Google Scholar] [CrossRef]
- Funderburk, J.; Stavisky, J.; Olson, S. Predation of Franklinella occidentalis (Thysanoptera: Thripidae) in field peppers by Orius insidiosus (Hemiptera: Anthocoridae). Environ. Entomol. 2000, 29, 376–382. [Google Scholar] [CrossRef]
- Baez, I.; Reitz, S.R.; Funderburk, J.E. Predation by Orius insidiosus (Hemiptera: Anthocoridae) on life stages and species of Frankliniella flower thrips (Thysanoptera: Thripidae) in pepper flowers. Environ. Entomol. 2004, 33, 662–670. [Google Scholar] [CrossRef]
- Reitz, S.R.; Funderburk, J.; Waring, S. Differential predation by the generalist predator Orius insidiosus on congeneric species of thrips that vary in size and behavior. Entomol. Exp. Appl. 2006, 119, 179–188. [Google Scholar] [CrossRef]
- Tyler-Julian, K.; Funderburk, J.; Frantz, G.; Mellinger, C. Evaluation of a push-pull strategy for the management of Frankliniella bispinosa (Thysanoptera: Thripidae) in bell pepper. Environ. Entomol. 2014, 43, 1364–1378. [Google Scholar] [CrossRef]
- Baez, I.; Reitz, S.R.; Funderburk, J.E.; Olson, S.M. Variation within and between Frankliniella thrips species in host plant utilization. J. Insect Sci. 2012, 11, 41. Available online: https://0-academic-oup-com.brum.beds.ac.uk/jinsectscience/article/11/1/41/2492490 (accessed on 25 September 2018).
- Coll, C.; Ridgway, R.L. Functional and numerical response of Orius insidiosus (Heteroptera: Anthocoridae) to its prey in different vegetable crops. Ann. Entomol. Soc. Am. 1995, 88, 732–738. [Google Scholar] [CrossRef]
- Funderburk, J. Management of the western flower thrips (Thysanoptera: Thripidae) in fruiting vegetables. Florida Entomol. 2009, 92, 1–6. [Google Scholar] [CrossRef]
- Reitz, S.R.; Funderburk, J. Management Strategies for Western Flower Thrips and the Role of Insecticides, Insecticides—Pest Engineering; Perveen, F., Ed.; InTech: Rijeka, Croatia, 2012; Available online: http://www.intechopen.com/books/insecticides-pest-engineering/management-strategies-for-western-flower-thrips-and-the-role-of-insecticides (accessed on 26 September 2018).
- Momol, M.T.; Olson, S.M.; Funderburk, J.E.; Stavisky, J.; Marois, J.J. Integrated management of tomato spotted wilt on field-grown tomatoes. Plant Dis. 2004, 88, 882–890. [Google Scholar] [CrossRef]
- Reitz, S.R.; Yearby, E.L.; Funderburk, J.E.; Stavisky, J.; Momol, M.T.; Olson, S.M. Integrated management tactics for Frankliniella thrips (Thysanoptera: Thripidae) in field-grown pepper. J. Econ. Entomol. 2003, 96, 1201–1214. [Google Scholar] [CrossRef]
- Gao, Y.; Lei, Z.; Reitz, S.R. Western flower thrips resistance: Detection, mechanisms, and management strategies. Pest Manag. Sci. 2012, 68, 1111–1121. [Google Scholar] [CrossRef]
- Terry, L.I. Host selection, communication, and reproductive behavior. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Wallingford, UK, 1997; pp. 65–118. ISBN 0-85199-178-5. [Google Scholar]
- Cantore, V.; Pace, B.; Albrizio, R. Kaolin-based particle film technology affects tomato physiology, yield and quality. Environ. Exp. Bot. 2009, 66, 279–288. [Google Scholar] [CrossRef]
- Bar-Joseph, M.; Frenkel, H. Spraying citrus plants with kaolin suspensions reduces the spirea aphid (Aphis citricola van der Goot). Crop Prot. 1983, 2, 371–374. [Google Scholar] [CrossRef]
- Lapointe, S.L. Particle film deters oviposition by Diaprepes abbreviatus (Coleoptera: Curculionidae). J. Econ. Entomol. 2000, 93, 1459–1463. [Google Scholar] [CrossRef]
- Barker, J.E.; Fulton, A.; Evans, K.A.; Powell, G. The effects of kaolin particle film Plutella xylostella behaviour and development. Pest Manag. Sci. 2006, 62, 498–504. [Google Scholar] [CrossRef]
- Larentzaki, E.; Shelton, A.M.; Plate, J. Effect of kaolin particle film on Thrips tabaci (Thysanoptera: Thripidae) oviposition, feeding, and development on onions: A lab and field case study. Crop Prot. 2008, 25, 727–734. [Google Scholar] [CrossRef]
- Peng, L.; Trumble, J.T.; Munyaneza, J.E.; Liu, T.-X. Repellancy of a kaolin particle film to potato psyllid, Bactericera cockerelli (Hemiptera: Psyllidae), on tomato under laboratory and field conditions. Pest Manag. Sci. 2011, 67, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Spiers, J.D.; Matta, F.B.; Marshall, D.A.; Sampson, B.J. Effects of kaolin clay application on flower bud development, fruit quality, and yield, and flower thrips [Frankliniella spp. (Thysanoptera: Thripidae)] populations on blueberry plants. Small Fruits Rev. 2004, 3, 361–373. [Google Scholar] [CrossRef]
- Reitz, S.R.; Maiorino, G.; Olson, S.; Sprenkel, R.; Crescenzi, A.; Momol, M.T. Integrating plant essential oils and kaolin for the sustainable management of thrips and tomato spotted wilt on tomato. Plant Dis. 2008, 92, 878–886. [Google Scholar] [CrossRef]
- Cook, S.M.; Khan, Z.R.; Picket, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Tyler-Julian, K.A.; Funderburk, J.E.; Olson, S.M.; Paret, M.L.; Webster, C.G.; Adkins, S. A stimulo-deterrent method of thrips and Tomato spotted wilt virus management in tomatoes. Acta Hortic. 2015, 1069, 251–258. [Google Scholar] [CrossRef]
- OMRI (Organic Materials Review Institute). Surround WP Crop Protectant; OMRI: Eugene, OR, USA, 2018; Available online: http://www.omri.org (accessed on 25 September 2018).
- Hansen, E.A.; Funderburk, J.E.; Reitz, S.R.; Ramachandran, S.; Eger, J.E.; McAuslane, H. Within-plant distribution of Frankliniella species (Thysanoptera: Thripidae) and Orius insidiosus (Heteroptera: Anthocoridae) in field pepper. Environ. Entomol. 2003, 32, 1035–1044. [Google Scholar] [CrossRef]
- Northfield, T.D.; Paini, D.R.; Funderburk, J.E.; Reitz, S.R. Annual cycles of Frankliniella spp. (Thysanoptera: Thripidae) thrips abundance on north Florida uncultivated reproductive hosts: Predicting possible sources of pest outbreaks. Ann. Entomol. Soc. Am. 2008, 101, 769–778. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT 9.2 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]
- Funderburk, C.; Funderburk, J.; Tyler-Julian, K.; Srivastava, M.; Knox, G.; Andersen, P.; Adkins, S. Population dynamics of Frankliniella bispinosa (Thysanoptera: Thripidae) and the predator Orius insidiosus (Hemiptera: Anthocoridae) as influenced by flower color of Lagerstroemia (Lythraceae). Environ. Entomol. 2015, 44, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Mound, L.A. Homologies and host-plant specificity: Recurrent problems in the study of thrips. Fla. Entomol. 2013, 96, 318–322. [Google Scholar] [CrossRef]
- Bottenberg, H.; Frantz, G.; Mellinger, H.C. Refuge and cover crop plantings for beneficial insect habitats. Proc. Fla. State Hort. Soc. 1999, 112, 339–341. [Google Scholar]
- Kasina, J.; Nderitu, J.; Nyamasyo, G.; Olubayo, E.; Waturu, C.; Obudho, E.; Yobera, D. Evaluation of companion plant for thrips (Thysanoptera: Thripidae) management on French green bean Phaseolus vulgaris (Fabaceae). Int. J. Trop. Insect Sci. 2006, 26, 121–125. [Google Scholar] [CrossRef]
- Lopez, R.; Shepard, B.M. Feverfew as a companion crop reduces spider mites, whiteflies, and thrips in other medicinal plants. Acta Hortic. 2007, 756, 33–38. [Google Scholar] [CrossRef]
- Frantz, G.; Mellinger, H.C. Shifts in western flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae), population abundance and crop damage. Fla. Entomol. 2009, 92, 29–34. [Google Scholar] [CrossRef]
- Nyasani, J.O.; Meyhofer, R.; Subramanian, S.; Poehling, H.M. Effect of intercrops on thrips species composition and population abundance on French beans in Kenya. Entomol. Exp. Appl. 2012, 142, 236–246. [Google Scholar] [CrossRef]
- Matteson, N.; Terry, L.I. Response to color by male and female Frankliniella occidentalis during swarming and non-swarming behavior. Entomol. Exp. Appl. 1992, 63, 187–201. [Google Scholar] [CrossRef]
- Potting, R.P.J.; Perry, J.N.; Powell, W. Insect behavioral ecology and other factors affecting the ecology control efficacy of agro-ecosystem diversification strategies. Ecol. Model. 2005, 182, 199–216. [Google Scholar] [CrossRef]
- Needham, J.G. Ecological notes on the insect population in the flower heads of Bidens pilosa. Ecol. Monogr. 1948, 18, 431–446. [Google Scholar] [CrossRef]
- Ramachandran, S.; Funderburk, J.; Stavisky, J.; Olson, S. Population abundance and movement of Frankliniella species and Orius insidiosus in field pepper. Agric. For. Entomol. 2001, 3, 129–137. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Mean no. per 10 Tomato Flowers (SEM) | |||||||
|---|---|---|---|---|---|---|---|---|
| F. occidentalis | F. tritici | F. bispinosa | Thrips Larvae | O. insidiosus | ||||
| Males | Females | Males | Females | Males | Females | |||
| 2011 | ||||||||
| Black mulch | 2.1 (0.2) | 3.1 (0.3) | 12.0 (1.6) | 16.7 (2.0) | 0.6 (0.1) | 1.1 (0.2) | 6.8 (0.9) | 0.10 (0.05) |
| Black mulch and companion plants | 1.5 (0.2) | 3.1 (0.4) | 10.1 (1.5) | 12.2 (1.3) | 0.7 (0.1) | 0.3 (0.1) | 5.7 (0.7) | 0.19 (0.06) |
| Black mulch and kaolin | 1.5 (0.2) | 3.7 (0.4) | 3.0 (0.5) | 5.1 (0.7) | 0.2 (0.1) | 0.5 (0.1) | 4.1 (0.6) | 0.06 (0.03) |
| Black mulch and companion plants and kaolin | 1.1 (0.2) | 3.4 (0.4) | 3.7 (0.6) | 6.9 (0.9) | 0.3 (0.1) | 0.3 (0.1) | 4.5 (0.7) | 0.14 (0.05) |
| UV mulch | 2.0 (0.3) | 3.0 (0.3) | 3.9 (0.5) | 6.5 (0.8) | 0.5 (0.1) | 0.8 (0.2) | 5.3 (0.9) | 0.12 (0.04) |
| UV mulch and companion plants | 1.2 (0.2) | 3.0 (0.3) | 3.7 (0.6) | 8.3 (1.0) | 0.2 (0.1) | 0.7 (0.2) | 4.9 (0.8) | 0.19 (0.05) |
| UV mulch and kaolin | 1.3 (0.3) | 2.5 (0.3) | 2.4 (0.5) | 4.4 (0.6) | 0.2 (0.1) | 0.5 (0.1) | 3.9 (0.6) | 0.13 (0.05) |
| UV mulch and companion plants and kaolin | 0.6 (0.1) | 2.2 (0.2) | 1.6 (0.3) | 4.3 (0.5) | 0.1 (0.0) | 0.5 (0.1) | 3.7 (0.5) | 0.09 (0.03) |
| Analysis of variance F-value | ||||||||
| Date (12, 26 df) | 20.0 *** | 22.8 *** | 102.6 *** | 70.3 *** | 20.1 *** | 24.6 *** | 81.4 *** | 3.4 ** |
| Rep (2, 26 df) | 4.6 * | 5.0 * | 1.3 | 0.5 | 0.1 | 1.3 | 0.7 | 1.6 |
| Mulch (1, 26 df) | 11.2 ** | 7.8 ** | 111.9 *** | 48.1 *** | 8.2 ** | 0.5 | 13.2 ** | 0.2 |
| Date X mulch (12, 26 df) | 3.7 ** | 8.1 *** | 7.2 *** | 4.6 *** | 2.2 * | 2.9 ** | 2.0 | 0.5 |
| Kaolin (1, 52 df) | 39.3 *** | 1.1 | 278.1 *** | 169.0 *** | 10.1 ** | 24.0 ** | 24.9 *** | 1.7 |
| Date X kaolin (12, 52 df) | 1.7 | 1.7 | 6.1 *** | 4.6 *** | 2.3* | 3.9 *** | 1.7 | 0.5 |
| Mulch X kaolin (1, 52 df) | 0.8 | 5.4* | 35.0 *** | 21.3 *** | 0.1 | 3.7 | 8.0 ** | 0.0 |
| Date X mulch X kaolin (12, 52 df) | 0.5 | 1.7 | 1.6 | 1.9* | 0.9 | 1.1 | 1.3 | 0.9 |
| Companion plant (1, 104 df) | 21.3 *** | 2.0 | 5.8* | 2.0 | 12.1 *** | 7.3 ** | 0.6 | 2.2 |
| Date X companion plant (12, 104 df) | 3.0 ** | 1.8 | 2.4 ** | 0.9 | 2.5 ** | 0.9 | 0.3 | 0.4 |
| Mulch X companion plant (1, 104 df) | 0.3 | 0.5 | 0.0 | 0.2 | 0.1 | 0.7 | 0.2 | 0.8 |
| Date X mulch X companion plant (12, 104 df) | 0.5 | 1.4 | 1.7 | 0.3 | 0.9 | 1.0 | 2.0* | 0.4 |
| Kaolin X companion plant (1, 104 df) | 0.5 | 1.4 | 7.2 ** | 2.8 | 12.4 *** | 0.2 | 1.4 | 0.9 |
| Date X kaolin X companion plant (1, 104 df) | 0.5 | 0.9 | 1.6 | 1.1 | 2.6 ** | 1.2 | 0.5 | 1.0 |
| Mulch X kaolin X companion plant (1, 104 df) | 0.1 | 0.1 | 9.3 ** | 12.8 *** | 3.6 | 0.3 | 0.6 | 0.3 |
| Date X mulch X kaolin X companion plant (12, 104 df) | 0.4 | 0.8 | 1.7 | 1.3 | 1.5 | 1.2 | 0.7 | 1.7 |
| 2012 | ||||||||
| Black mulch | 0.9 (0.1) | 1.6 (0.2) | 2.6 (0.4) | 6.9 (0.7) | 7.6 (0.9) | 26.4 (3.3) | 8.1 (0.9) | 0.14 (0.05) |
| Black mulch and companion plants | 0.6 (0.1) | 1.6 (0.2) | 1.1 (0.2) | 4.4 (0.3) | 4.7 (0.6) | 20.9 (2.2) | 6.1 (0.7) | 0.18 (0.05) |
| Black mulch and kaolin | 1.1 (0.2) | 2.7 (0.3) | 0.7 (0.1) | 3.3 (0.4) | 3.5 (0.5) | 15.7 (2.2) | 7.3 (0.9) | 0.14 (0.05) |
| Black mulch and companion plants and kaolin | 0.8 (0.1) | 2.5 (0.3) | 0.4 (0.1) | 3.4 (0.3) | 2.4 (0.3) | 13.2 (1.6) | 7.7 (0.8) | 0.14 (0.04) |
| UV mulch | 1.2 (0.2) | 2.2 (0.3) | 1.0 (0.2) | 4.1 (0.4) | 4.6 (0.7) | 22.2 (2.9) | 7.4 (1.1) | 0.19 (0.06) |
| UV mulch and companion plants | 0.8 (0.1) | 2.4 (0.3) | 0.6 (0.1) | 3.8 (0.3) | 3.2 (0.4) | 18.2 (2.1) | 10.3 (1.3) | 0.19 (0.06) |
| UV mulch and kaolin | 0.9 (0.2) | 2.4 (0.2) | 0.4 (0.1) | 2.5 (0.3) | 2.2 (0.4) | 14.8 (2.1) | 6.8 (0.8) | 0.13 (0.04) |
| UV mulch and companion plants and kaolin | 0.6 (0.1) | 2.1 (0.2) | 0.4 (0.1) | 2.7 (0.3) | 1.7 (0.2) | 12.0 (1.5) | 7.9 (0.9) | 0.15 (0.06) |
| Analysis of variance F-value | ||||||||
| Date (11, 24 df) | 3.6 *** | 26.3 *** | 19.9 *** | 25.4 *** | 83.7 *** | 95.0 *** | 24.0 *** | 6.2 *** |
| Rep (2, 24 df) | 0.7 | 0.9 | 1.9 | 5.9 ** | 0.3 | 10.0 *** | 4.8* | 1.8 |
| Mulch (1, 24 df) | 0.2 | 2.2 | 21.8 *** | 24.2 *** | 37.5 *** | 7.7 ** | 0.0 | 0.0 |
| Date X mulch (11, 24 df) | 1.2 | 2.1 | 2.1 | 3.0 ** | 5.5 *** | 2.4 * | 0.9 | 0.4 |
| Kaolin (1, 48 df) | 0.0 | 19.5 *** | 74.6 *** | 72.7 *** | 91.6 *** | 111.1 *** | 0.2 | 1.0 |
| Date X kaolin (11, 48 df) | 1.9 | 0.9 | 4.7 *** | 3.3 ** | 3.5* | 3.6 ** | 1.9 | 0.7 |
| Mulch X Kaolin (1, 48 df) | 8.2 ** | 7.7 ** | 13.0 *** | 2.7 | 0.3 | 0.8 | 0.6 | 0.1 |
| Date X mulch X kaolin (11, 48 df) | 1.0 | 0.9 | 2.3 * | 1.8 | 0.8 | 0.7 | 0.2 | 1.1 |
| Companion plant (1, 96 df) | 10.3 ** | 0.7 | 26.2 *** | 0.0 | 11.7 *** | 4.1* | 4.0* | 0.3 |
| Date X companion plant (11, 96 df) | 2.0 * | 1.7 | 2.7 ** | 2.9 ** | 4.3 *** | 3.7 *** | 1.9 * | 0.8 |
| Mulch X companion plant (1, 96 df) | 0.3 | 0.0 | 6.6 ** | 3.4 | 4.1 * | 0.0 | 6.6 ** | 0.1 |
| Date X mulch X companion plant (11, 96 df) | 0.8 | 0.9 | 1.5 | 0.9 | 1.0 | 1.1 | 1.2 | 0.5 |
| Kaolin X companion plant (1, 96 df) | 0.1 | 2.5 | 6.2 ** | 6.1 ** | 2.1 | 1.1 | 0.0 | 0.1 |
| Date X kaolin X companion plant (11, 96 df) | 1.0 | 0.5 | 1.7 | 0.7 | 1.2 | 0.9 | 0.9 | 0.5 |
| Mulch X kaolin X companion plant (1, 96 df) | 0.3 | 0.7 | 0.2 | 2.4 | 0.0 | 0.0 | 6.6 ** | 0.1 |
| Date X mulch X kaolin X companion plant (11, 96 df) | 0.7 | 0.7 | 0.9 | 0.8 | 0.7 | 0.7 | 1.3 | 0.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyler-Julian, K.; Funderburk, J.; Srivastava, M.; Olson, S.; Adkins, S. Evaluation of a Push-Pull System for the Management of Frankliniella Species (Thysanoptera: Thripidae) in Tomato. Insects 2018, 9, 187. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040187
Tyler-Julian K, Funderburk J, Srivastava M, Olson S, Adkins S. Evaluation of a Push-Pull System for the Management of Frankliniella Species (Thysanoptera: Thripidae) in Tomato. Insects. 2018; 9(4):187. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040187
Chicago/Turabian StyleTyler-Julian, Kara, Joe Funderburk, Mrittunjai Srivastava, Steve Olson, and Scott Adkins. 2018. "Evaluation of a Push-Pull System for the Management of Frankliniella Species (Thysanoptera: Thripidae) in Tomato" Insects 9, no. 4: 187. https://0-doi-org.brum.beds.ac.uk/10.3390/insects9040187