
Pan-Piscine Orthoreovirus (PRV) Detection Using Reverse Transcription Quantitative PCR

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Pan-PRV RT-qPCR Assay Development and Optimization
2.1.1. Primer and Probe Design
2.1.2. Primer and Probe Set Elimination and qPCR Optimization
2.2. Pan-PRV RT-qPCR Assay Universality, Exclusivity, and Sensitivity
2.2.1. Universal Detection of PRV Genogroups
2.2.2. Exclusivity of Pan-PRV RT-qPCR
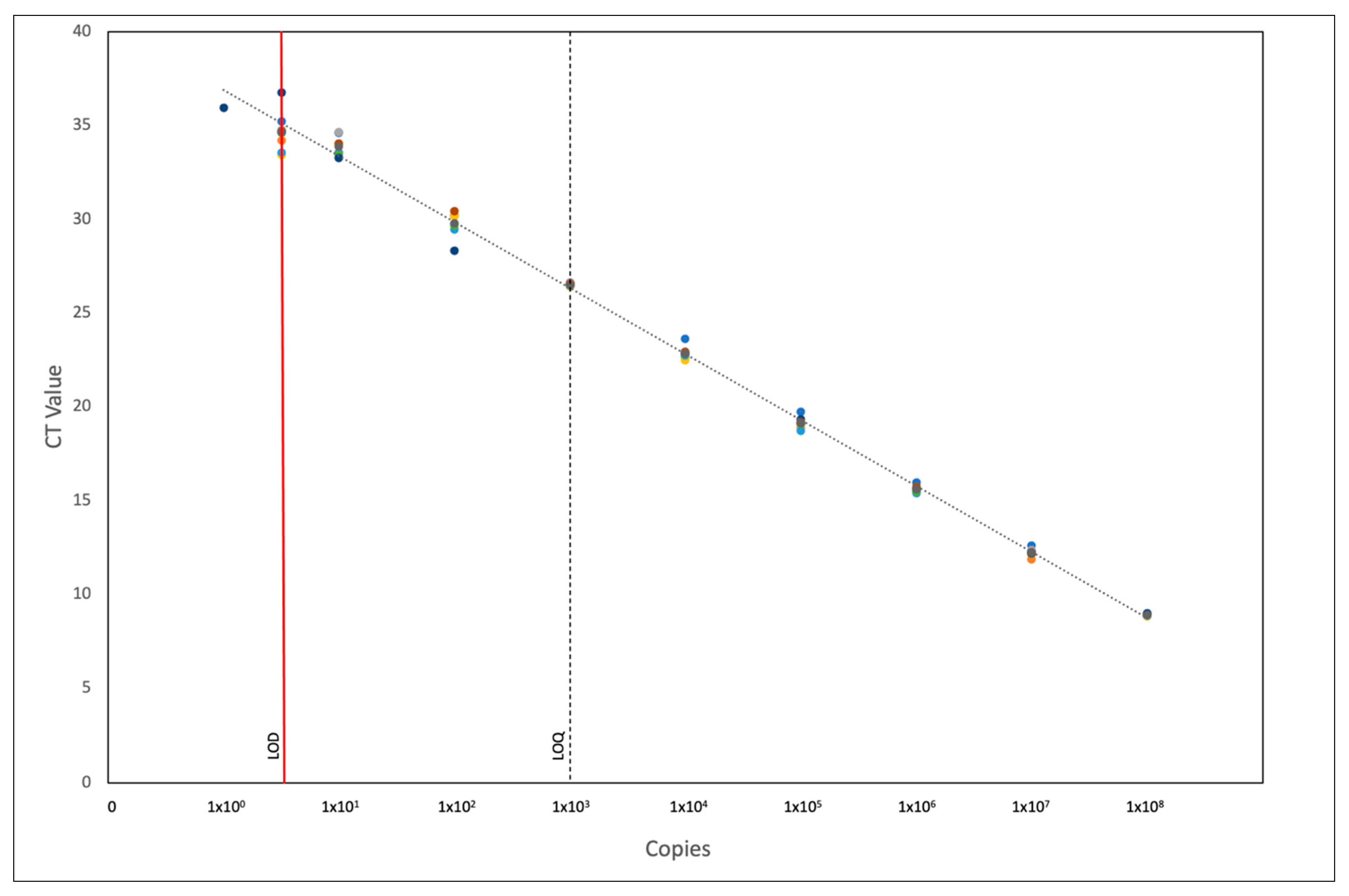
2.2.3. Pan-PRV qPCR Sensitivity
2.3. One-Step Pan-PRV RT-qPCR
2.4. Reproducibility and Robustness—Inter-Laboratory Comparison
Inter-Laboratory Proficiency Test
3. Discussion
4. Materials and Methods
4.1. Pan-PRV RT-qPCR Assay Development and Optimization
4.1.1. PRV Source Material, RNA Extraction, and cDNA Synthesis
4.1.2. Primer and Probe Design
4.1.3. Primer and Probe Set Elimination and qPCR Optimization
4.2. Pan-PRV RT-qPCR Assay Universality, Exclusivity, and Sensitivity
4.2.1. Universal Detection of PRV Genotypes
4.2.2. Exclusivity of Pan-PRV RT-qPCR
4.2.3. Pan-PRV qPCR Sensitivity
4.3. One-Step Pan-PRV RT-qPCR
4.4. Reproducibility and Robustness—Inter-Laboratory Comparison
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polinski, M.P.; Vendramin, N.; Cuenca, A.; Garver, K.A. Piscine orthoreovirus: Biology and distribution in farmed and wild fish. J. Fish Dis. 2020, 43, 1331–1352. [Google Scholar] [CrossRef]
- Kibenge, M.J.; Iwamoto, T.; Wang, Y.; Morton, A.; Godoy, M.G.; Kibenge, F.S. Whole-genome analysis of piscine reovirus (PRV) shows PRV represents a new genus in family Reoviridae and its genome segment S1 sequences group it into two separate sub-genotypes. Virol. J. 2013, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Palacios, G.; Lovoll, M.; Tengs, T.; Hornig, M.; Hutchison, S.; Hui, J.; Kongtorp, R.T.; Savji, N.; Bussetti, A.V.; Solovyov, A.; et al. Heart and skeletal muscle inflammation of farmed salmon is associated with infection with a novel reovirus. PLoS ONE 2010, 5, e11487. [Google Scholar] [CrossRef]
- Dhamotharan, K.; Vendramin, N.; Markussen, T.; Wessel, Ø.; Cuenca, A.; Nyman, I.B.; Olsen, A.B.; Tengs, T.; Krudtaa Dahle, M.; Rimstad, E. Molecular and Antigenic Characterization of Piscine orthoreovirus (PRV) from Rainbow Trout (Oncorhynchus mykiss). Viruses 2018, 10, 170. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Nawata, A.; Sakai, T.; Matsuyama, T.; Ito, T.; Kurita, J.; Terashima, S.; Yasuike, M.; Nakamura, Y.; Fujiwara, A.; et al. Full-Genome Sequencing and Confirmation of the Causative Agent of Erythrocytic Inclusion Body Syndrome in Coho Salmon Identifies a New Type of Piscine Orthoreovirus. PLoS ONE 2016, 11, e0165424. [Google Scholar] [CrossRef] [Green Version]
- Wessel, Ø.; Braaen, S.; Alarcon, M.; Haatveit, H.; Roos, N.; Markussen, T.; Tengs, T.; Dahle, M.K.; Rimstad, E. Infection with purified Piscine orthoreovirus demonstrates a causal relationship with heart and skeletal muscle inflammation in Atlantic salmon. PLoS ONE 2017, 12, e0183781. [Google Scholar] [CrossRef] [Green Version]
- Wessel, Ø.; Hansen, E.F.; Dahle, M.K.; Alarcon, M.; Vatne, N.A.; Nyman, I.B.; Soleim, K.B.; Dhamotharan, K.; Timmerhaus, G.; Markussen, T.; et al. Piscine Orthoreovirus-1 Isolates Differ in Their Ability to Induce Heart and Skeletal Muscle Inflammation in Atlantic Salmon (Salmo salar). Pathogens 2020, 9, 1050. [Google Scholar] [CrossRef]
- Vendramin, N.; Kannimuthu, D.; Olsen, A.B.; Cuenca, A.; Teige, L.H.; Wessel, Ø.; Iburg, T.M.; Dahle, M.K.; Rimstad, E.; Olesen, N.J. Piscine orthoreovirus subtype 3 (PRV-3) causes heart inflammation in rainbow trout (Oncorhynchus mykiss). Vet. Res. 2019, 50, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartagena, J.; Tambley, C.; Sandino, A.M.; Spencer, E.; Tello, M. Detection of piscine orthoreovirus in farmed rainbow trout from Chile. Aquaculture 2018, 528, 735480. [Google Scholar] [CrossRef]
- Godoy, M.G.; Kibenge, M.J.; Wang, Y.; Suarez, R.; Leiva, C.; Vallejos, F.; Kibenge, F.S. First description of clinical presentation of piscine orthoreovirus (PRV) infections in salmonid aquaculture in Chile and identification of a second genotype (Genotype II) of PRV. Virol. J. 2016, 13, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Løvoll, M.; Alarcón, M.; Bang Jensen, B.; Taksdal, T.; Kristoffersen, A.B.; Tengs, T. Quantification of piscine reovirus (PRV) at different stages of Atlantic salmon Salmo salar production. Dis. Aquat. Organ. 2012, 99, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marty, G.D.; Morrison, D.B.; Bidulka, J.; Joseph, T.; Siah, A. Piscine reovirus in wild and farmed salmonids in British Columbia, Canada: 1974–2013. J. Fish Dis. 2015, 38, 713–728. [Google Scholar] [CrossRef]
- Purcell, M.K.; Powers, R.L.; Evered, J.; Kerwin, J.; Meyers, T.R.; Stewart, B.; Winton, J.R. Molecular testing of adult Pacific salmon and trout (Oncorhynchus spp.) for several RNA viruses demonstrates widespread distribution of piscine orthoreovirus in Alaska and Washington. J. Fish Dis. 2018, 41, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.; Vendramin, N.; Priess, C.; Kannimuthu, D.; Henriksen, N.H.; Iburg, T.M.; Olesen, N.J.; Cuenca, A. Emergence and Spread of Piscine orthoreovirus Genotype 3. Pathogens 2020, 9, 823. [Google Scholar] [CrossRef] [PubMed]
- Vendramin, N.; Cuenca, A.; Sørensen, J.; Alencar, A.L.F.; Christiansen, D.H.; Jacobsen, J.A.; Axen, C.; Lieffrig, F.; Ruane, N.M.; Martin, P.; et al. Presence and genetic variability of Piscine orthoreovirus genotype 1 (PRV-1) in wild salmonids in Northern Europe and North Atlantic Ocean. J. Fish Dis. 2019, 42, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Dhamotharan, K.; Tengs, T.; Wessel, Ø.; Braaen, S.; Nyman, I.B.; Hansen, E.F.; Christiansen, D.H.; Dahle, M.K.; Rimstad, E.; Markussen, T. Evolution of the Piscine orthoreovirus Genome Linked to Emergence of Heart and Skeletal Muscle Inflammation in Farmed Atlantic Salmon (Salmo salar). Viruses 2019, 11, 465. [Google Scholar] [CrossRef] [Green Version]
- Polinski, M.P.; Marty, G.D.; Snyman, H.N.; Garver, K.A. Piscine orthoreovirus demonstrates high infectivity but low virulence in Atlantic salmon of Pacific Canada. Sci. Rep. 2019, 9, 3297. [Google Scholar] [CrossRef] [Green Version]
- Pham, P.H.; Misk, E.; Papazotos, F.; Jones, G.; Polinski, M.P.; Contador, E.; Russell, S.; Garver, K.A.; Lumsden, J.S.; Bols, N.C. Screening of Fish Cell Lines for Piscine Orthoreovirus-1 (PRV-1) Amplification: Identification of the Non-Supportive PRV-1 Invitrome. Pathogens 2020, 9, 833. [Google Scholar] [CrossRef]
- Olsen, A.B.; Hjortaas, M.; Tengs, T.; Hellberg, H.; Johansen, R. First Description of a New Disease in Rainbow Trout (Oncorhynchus mykiss (Walbaum)) Similar to Heart and Skeletal Muscle Inflammation (HSMI) and Detection of a Gene Sequence Related to Piscine Orthoreovirus (PRV). PLoS ONE 2015, 10, e0131638. [Google Scholar] [CrossRef]
- Adamek, M.; Hellmann, J.; Flamm, A.; Teitge, F.; Vendramin, N.; Fey, D.; Riße, K.; Blakey, F.; Rimstad, E.; Steinhagen, D. Detection of piscine orthoreoviruses (PRV-1 and PRV-3) in Atlantic salmon and rainbow trout farmed in Germany. Transbound. Emerg. Dis. 2019, 66, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Bigarre, L. First detection of Piscine reovirus in France. In Proceedings of the Oral Presentation at PD-Trination Meeting, Aberdeen, UK, 12–13 October 2016. [Google Scholar]
- Hauge, H.; Vendramin, N.; Taksdal, T.; Olsen, A.B.; Wessel, Ø.; Mikkelsen, S.S.; Alencar, A.L.F.; Olesen, N.J.; Dahle, M.K. Infection experiments with novel Piscine orthoreovirus from rainbow trout (Oncorhynchus mykiss) in salmonids. PLoS ONE 2017, 12, e0180293. [Google Scholar] [CrossRef] [Green Version]
- Labrut, S.; Bigarré, L.; Boitard, P.; Jamin, M. Emergence of the heart and skeletal muscle inflammation syndrome in France. In Proceedings of the World Aquaculture Society Meeting, Montpellier, France, 25–29 August 2018. [Google Scholar]
- Rodger, H.D.; McCleary, S.J.; Ruane, N.M. Clinical cardiomyopathy syndrome in Atlantic salmon, Salmo salar L. J. Fish Dis. 2014, 37, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Kralik, P.; Ricchi, M. A Basic Guide to Real Time PCR in Microbial Diagnostics: Definitions, Parameters, and Everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haugland, O.; Mikalsen, A.B.; Nilsen, P.; Lindmo, K.; Thu, B.J.; Eliassen, T.M.; Roos, N.; Rode, M.; Evensen, O. Cardiomyopathy syndrome of atlantic salmon (Salmo salar L.) is caused by a double-stranded RNA virus of the Totiviridae family. J. Virol. 2011, 85, 5275–5286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy, M.; Medina, D.A.; Suarez, R.; Valenzuela, S.; Romero, J.; Kibenge, M.; Wang, Y.; Kibenge, F. Extensive Phylogenetic Analysis of Piscine Orthoreovirus Genomic Sequences Shows the Robustness of Subgenotype Classification. Pathogens 2021, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Markussen, T.; Dahle, M.K.; Tengs, T.; Løvoll, M.; Finstad, Ø.; Wiik-Nielsen, C.R.; Grove, S.; Lauksund, S.; Robertsen, B.; Rimstad, E. Sequence analysis of the genome of piscine orthoreovirus (PRV) associated with heart and skeletal muscle inflammation (HSMI) in Atlantic salmon (Salmo salar). PLoS ONE 2013, 8, e70075. [Google Scholar] [CrossRef]
- Jayasuriya, A.K.; Nibert, M.L.; Fields, B.N. Complete nucleotide sequence of the M2 gene segment of reovirus type 3 dearing and analysis of its protein product mu 1. Virology 1988, 163, 591–602. [Google Scholar] [CrossRef]
- Nibert, M.L.; Odegard, A.L.; Agosto, M.A.; Chandran, K.; Schiff, L.A. Putative autocleavage of reovirus mu1 protein in concert with outer-capsid disassembly and activation for membrane permeabilization. J. Mol. Biol. 2005, 345, 461–474. [Google Scholar] [CrossRef]
- Varela, R.; Benavente, J. Protein coding assignment of avian reovirus strain S1133. J. Virol. 1994, 68, 6775–6777. [Google Scholar] [CrossRef] [Green Version]
- Haatveit, H.M.; Wessel, Ø.; Markussen, T.; Lund, M.; Thiede, B.; Nyman, I.B.; Braaen, S.; Dahle, M.K.; Rimstad, E. Viral Protein Kinetics of Piscine Orthoreovirus Infection in Atlantic Salmon Blood Cells. Viruses 2017, 9, 49. [Google Scholar] [CrossRef]
- Siah, A.; Breyta, R.B.; Warheit, K.I.; Gagne, N.; Purcell, M.K.; Morrison, D.; Powell, J.F.F.; Johnson, S.C. Genomes reveal genetic diversity of Piscine orthoreovirus in farmed and free-ranging salmonids from Canada and USA. Virus Evol. 2020, 6, veaa054. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Target | Name | Nucleotide Sequence (5′→3′) | Tm (°C) | GC (%) | Amplicon (bp) |
|---|---|---|---|---|---|
| PRV M2 | 28F | TGGGTAACTATCAGACAAGTAACAAC | 58.8 | 39 | |
| 112R | GTAGARTCGAGTCCGCCTTCAG | 60.5–62.1 | 57 | 85 | |
| 54P | FAM-CAATTTTGGGTAACTGGCGACGGCAATGA-MGBNFQ | 68.2 | 48 |
| Isolate. | Genotype | GenBank Accession No. | Source | Reference | |
|---|---|---|---|---|---|
| Host Species | Tissue | ||||
| 16-005 | PRV-1a | MH347359-MH347368 | Atlantic salmon | Blood | [17] |
| r17_1227 | PRV-1a | R17_1227: (MW354796, MW354808, MW354820, MW354832, MW354844, MW354856, MW354868, MW354880, MW354892, MW354904) | Atlantic salmon | Blood | [33] |
| NOR2012_V3621 | PRV-1b | KY429943-KY429952 | Atlantic salmon | Blood | [6] |
| --- | PRV-2 | LC145608-LC145617 | Coho salmon | Heart | [5] |
| NOR/060214 | PRV-3a | MG253807-MG253816 | Rainbow trout | Spleen and heart (pooled) | [4] |
| DK/PRV315 | PRV-3b | MW012855-MW012864 | Rainbow trout | Spleen and heart (pooled) | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Vendramin, N.; Cuenca, A.; Polinski, M.; Hawley, L.M.; Garver, K.A. Pan-Piscine Orthoreovirus (PRV) Detection Using Reverse Transcription Quantitative PCR. Pathogens 2021, 10, 1548. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121548
Zhao J, Vendramin N, Cuenca A, Polinski M, Hawley LM, Garver KA. Pan-Piscine Orthoreovirus (PRV) Detection Using Reverse Transcription Quantitative PCR. Pathogens. 2021; 10(12):1548. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121548
Chicago/Turabian StyleZhao, Julie, Niccolò Vendramin, Argelia Cuenca, Mark Polinski, Laura M. Hawley, and Kyle A. Garver. 2021. "Pan-Piscine Orthoreovirus (PRV) Detection Using Reverse Transcription Quantitative PCR" Pathogens 10, no. 12: 1548. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10121548