
Occurrence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter spp. in Intensive Pig Production in South Africa


 , and
, and
Abstract
:1. Introduction
2. Results
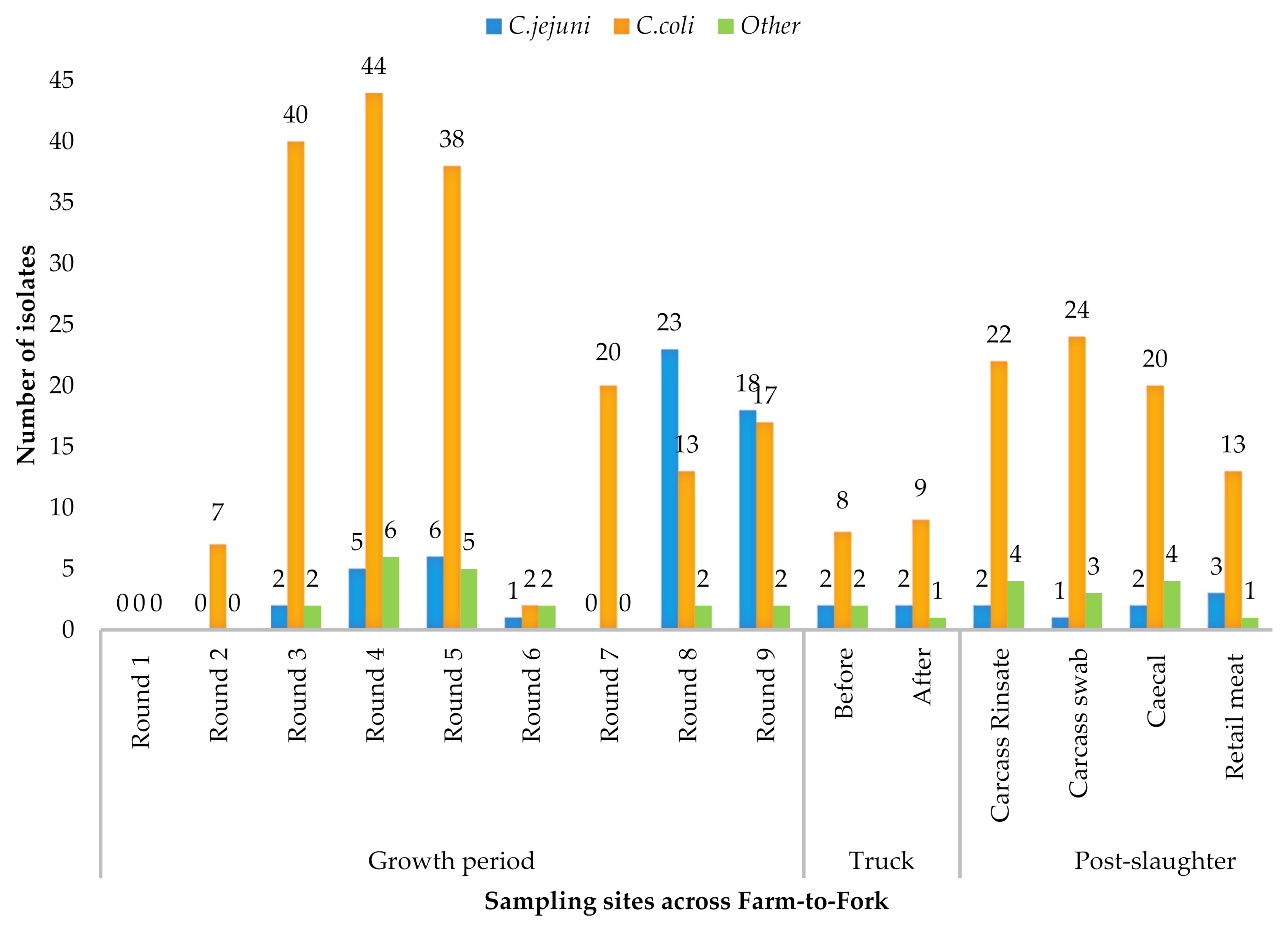
2.1. Prevalence of Campylobacter spp.
2.2. Antibiotic Susceptibility Profiles
2.3. Detection of Antibiotic Resistance Genes
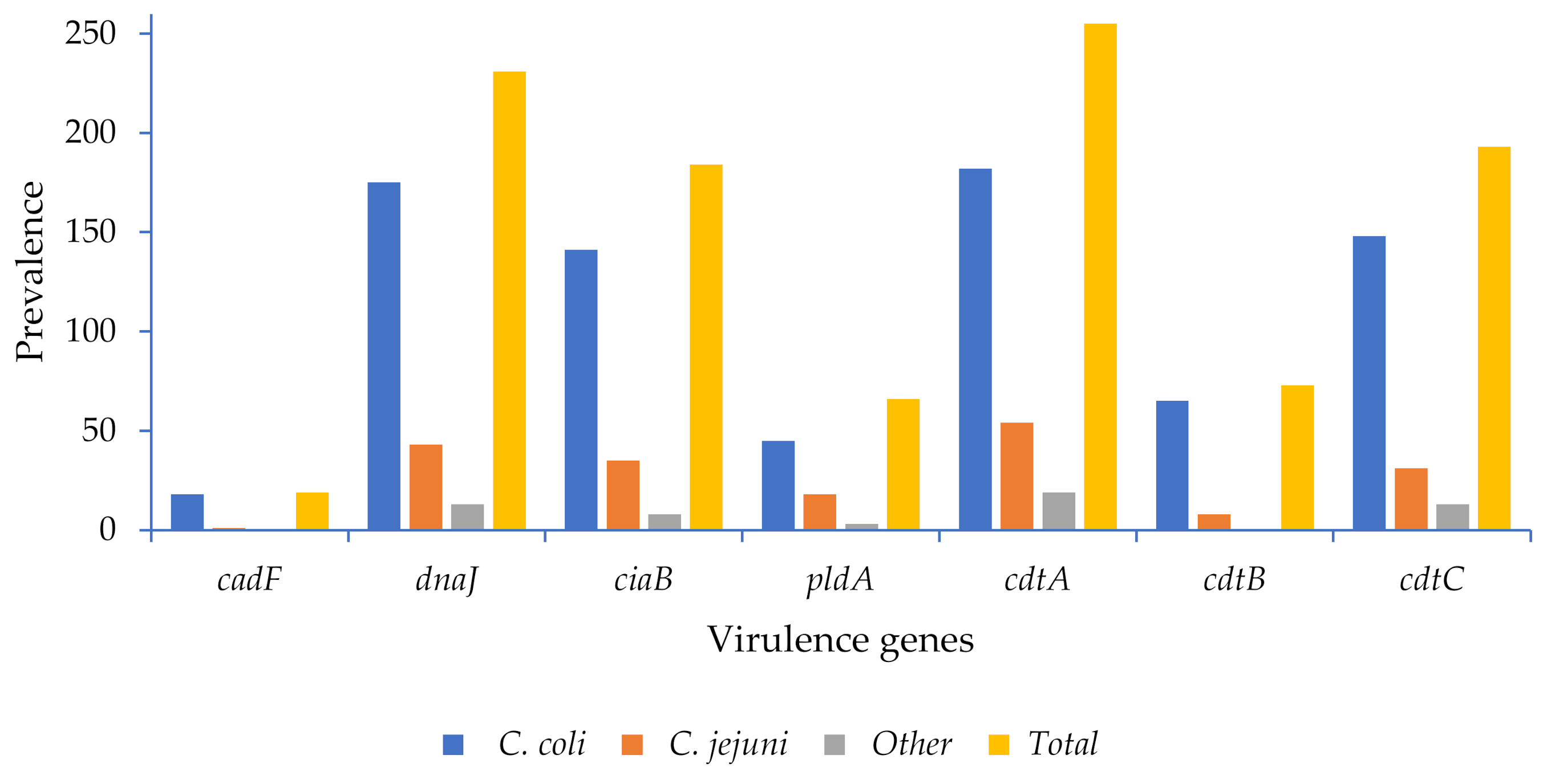
2.4. Virulence Genes
2.5. Clonality
3. Discussion
3.1. Prevalence of Campylobacter spp.
3.2. Antibiotic Resistance Profiles and Resistance Determinants
3.3. Virulence Determinants
3.4. Clonality
4. Materials and Methods
4.1. Ethical Considerations
4.2. Study Site
4.3. Participants
4.4. Sampling
- Production (animals on the farm): samples collected included feces, litter (the bedding), and slurry. These were collected every two weeks (nine rounds of sampling) from birth to slaughter over four months (September 2018–January 2019). The block sampling method was used to ensure an even representation of the entire herd within the pig pen house.
- Hand and nasal samples were collected from the farmworkers on Weeks 3, 5–7, and 9 over four months based on individual voluntary consent.
- Holding and transport samples consisted of swab samples from the holding pens and truck floor. These were collected before and after the transportation of the pigs to the abattoir.
- Post-slaughter (abattoir) samples included carcass swabs, caecal samples, and carcass rinsate.
- Swab samples of meat products, including whole cuts (head, body, and thigh) sold to consumers, were collected from the meat processing plant at the retail point.
4.5. Isolation of Campylobacter spp.
4.6. DNA Extraction
4.7. Molecular Confirmation of Isolates
4.8. Antibiotic Susceptibility Testing
4.9. Detection of Antibiotic Resistance and Virulence Genes
4.10. Clonality (ERIC-PCR)
4.11. Data Analysis and Interpretation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fèvre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic resistance is the quintessential One Health issue. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 377–380. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Karikari, A.B.; Obiri-Danso, K.; Frimpong, E.H.; Krogfelt, K.A. Antibiotic Resistance of Campylobacter recovered from faeces and carcasses of healthy livestock. Biomed Res. Int. 2017, 2017, 4091856. [Google Scholar] [CrossRef] [Green Version]
- Pillay, S.; Amoako, D.G.; Abia, A.L.K.; Somboro, A.M.; Shobo, C.O.; Perrett, K.; Bester, L.A.; Essack, S.Y. Characterisation of Campylobacter spp. Isolated from Poultry in KwaZulu-Natal, South Africa. Antibiotics 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igwaran, A.; Okoh, A.I. Campylobacteriosis agents in meat carcasses collected from two district municipalities in the Eastern Cape Province, South Africa. Foods 2020, 9, 203. [Google Scholar] [CrossRef] [Green Version]
- Heredia, N.; García, S. Animals as sources of foodborne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Frasao, B.D.S.; Marin, V.A.; Conte-Junior, C.A. Molecular detection, typing, and quantification of Campylobacter spp. in foods of animal origin. Compr. Rev. Food Sci. Food Saf. 2017, 16, 721–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Organisation for Animal Health. OIE List of Antimicrobial Agents of Veterinary Importance. 2019. Available online: https://www.oie.int/scientific-expertise/veterinary-products/antimicrobials/ (accessed on 3 July 2020).
- WHO. Integrated Surveillance of Antimicrobial Resistance in Foodborne Bacteria: Application of a One Health Approach; WHO: Geneva, Switzerland, 2017; ISBN 978-92-4-151241-1. [Google Scholar]
- Food and Agriculture Organization of the United Nations. The FAO Action Plan on Antimicrobial Resistance. 2016. Available online: http://www.fao.org/publications (accessed on 21 January 2021).
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, D.; Petridou, E.; Filioussis, G.; Papadopoulos, T.; Papageorgiou, K.; Chatzistilianou, M.; Kritas, S. Prevalence and antibiotic resistance of Campylobacter coli and Campylobacter jejuni in Greek swine farms. Am. J. Microbiol. Immunol. 2020, 5, 6. [Google Scholar]
- Gwimi, P.B.; Faleke, O.O.; Salihu, M.D.; Magaji, A.A.; Abubakar, M.B.; Nwankwo, I.O.; Ibitoye, E.B. Prevalence of Campylobacter species in fecal samples of pigs and humans from Zuru Kebbi State, Nigeria. Int. J. One Health 2015, 1, 1–5. [Google Scholar] [CrossRef]
- Van, T.T.H.; Yidana, Z.; Smooker, P.M.; Coloe, P.J. Antibiotic use in food animals worldwide, with a focus on Africa: Pluses and minuses. J. Glob. Antimicrob. Resist. 2020, 20, 170–177. [Google Scholar] [CrossRef] [PubMed]
- South African Veterinary Association (SAVA). Guidelines for the Use of Antimicrobials in the South African Pig Industry. Available online: http://www.sava.co.za/2017/05/26/antibiotic-guidelines-pigindustry/ (accessed on 23 January 2021).
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Future Microbiol. 2009, 4, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Jesse, T.W.; Englen, M.D.; Pittenger-Alley, L.G.; Fedorka-Cray, P.J. Two distinct mutations in gyrA lead to ciprofloxacin and nalidixic acid resistance in Campylobacter coli and Campylobacter jejuni isolated from chickens and beef cattle. J. Appl. Microbiol. 2006, 100, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, K.; Rao, T.; Kiranmayi, C.; Kavitha, L. Antibiogram of Campylobacter jejuni and Campylobacter coli isolated from animals, foods of animal origin and humans in areas surrounding Gannavaram, Andhra Pradesh. Int. J. Sci. Environ. Technol. 2019, 8, 233–246. [Google Scholar]
- Sproston, E.L.; Wimalarathna, H.M.L.; Sheppard, S.K. Trends in fluoroquinolone resistance in Campylobacter. Microb. Genom. 2018, 4, e000198. [Google Scholar] [CrossRef]
- Di Donato, G.; Marotta, F.; Nuvoloni, R.; Zilli, K.; Neri, D.; Di Sabatino, D.; Calistri, P.; Di Giannatale, E. Prevalence, population diversity and antimicrobial resistance of Campylobacter coli isolated in Italian swine at slaughterhouse. Microorganisms 2020, 8, 222. [Google Scholar] [CrossRef] [Green Version]
- Iovine, N.M. Resistance mechanisms in Campylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef] [Green Version]
- Sheykhsaran, E.; Baghi, H.B.; Soroush, M.H.; Ghotaslou, R. An overview of tetracyclines and related resistance mechanisms. Rev. Med. Microbiol. 2019, 30, 69–75. [Google Scholar] [CrossRef]
- Aroori, S.V.; Cogan, T.A.; Humphrey, T.J. The effect of growth temperature on the pathogenicity of Campylobacter. Curr. Microbiol. 2013, 67, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Wafaa, M.M.H.; Mekky, A.A.A.; Enany, M.E. Review on some virulence factors associated with Campylobacter colonization and infection in poultry and human. Am. J. Biomed. Sci. Res. 2019, 3, 460–463. [Google Scholar] [CrossRef]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a foodborne pathogen: A review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef] [Green Version]
- Biswas, D.; Hannon, S.J.; Townsend, H.G.G.; Potter, A.; Allan, B.J. Genes coding for virulence determinants of Campylobacter jejuni in human clinical and cattle isolates from Alberta, Canada, and their potential role in colonization of poultry. Int. Microbiol. 2011, 14, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Ngobese, B.; Zishiri, O.T.; El Zowalaty, M.E. Molecular detection of virulence genes in Campylobacter species isolated from livestock production systems in South Africa. J. Integr. Agric. 2020, 19, 1656–1670. [Google Scholar] [CrossRef]
- Lee, S.-K.; Park, H.-J.; Lee, J.-H.; Lim, J.-S.; Seo, K.-H.; Heo, E.-J.; Kim, Y.-J.; Wee, S.-H.; Moon, J.-S. Distribution and molecular characterization of Campylobacter species at different processing stages in two poultry processing plants. Foodborne Pathog. Dis. 2017, 14, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Shobo, C.O.; Bester, L.A.; Baijnath, S.; Somboro, A.M.; Peer, A.K.C.; Essack, S.Y. Antibiotic resistance profiles of Campylobacter species in the South Africa private health care sector. J. Infect. Dev. Ctries. 2016, 10, 1214–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.; Zishiri, O.T. Genetic characterisation of virulence genes associated with adherence, invasion and cytotoxicity in Campylobacter spp. Isolated from commercial chickens and human clinical cases. Onderstepoort J. Vet. Res. 2018, 85, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chukwu, M.O.; Abia, A.L.K.; Ubomba-Jaswa, E.; Obi, L.; Dewar, J.B. Dewar Characterization and Phylogenetic Analysis of Campylobacter Species Isolated from Paediatric Stool and Water Samples in the Northwest Province, South Africa. Int. J. Environ. Res. Public Health 2019, 16, 2205. [Google Scholar] [CrossRef] [Green Version]
- EUCAST European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 8. 2017. Available online: www.eucast.org/clinical_breakpoints/. (accessed on 5 January 2018).
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: 27th Edition Informational Supplement M100-S27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Cha, W.; Mosci, R.E.; Wengert, S.L.; Vargas, C.V.; Rust, S.R.; Bartlett, P.C.; Grooms, D.L.; Manning, S.D. Comparing the genetic diversity and antimicrobial resistance profiles of Campylobacter jejuni recovered from cattle and humans. Front. Microbiol. 2017, 9, 818. [Google Scholar] [CrossRef] [Green Version]
- McIver, K.S.; Amoako, D.G.; Abia, A.L.K.; Bester, L.A.; Chenia, H.Y.; Essack, S.Y. Molecular Epidemiology of Antibiotic-Resistant Escherichia coli from Farm-to-Fork in Intensive Poultry Production in KwaZulu-Natal, South Africa. Antibiotics 2020, 9, 850. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Total Samples | No. of Positive Samples | Sample Proportion Among the Positives | Total Isolates | C. coli (% Per Site) | C. jejuni (% Per Site) | Other (% Per Site) |
|---|---|---|---|---|---|---|---|
| Feces | 232 | 195 | 51.50% | 255 | 181 (65.3) | 55 (82.1) | 19 (59.9) |
| Litter | 16 | 16 | 4.20% | ||||
| Slurry | 97 | 44 | 11.60% | ||||
| Truck and crate swabs | 40 | 24 | 6.30% | 24 | 17 (6.1) | 4 (5.9) | 3 (8.8) |
| Carcass swabs | 39 | 31 | 8.20% | 82 | 66 (23.8) | 5 (7.5) | 11 (32.4) |
| Carcass rinsates | 32 | 25 | 6.60% | ||||
| Caeca | 44 | 26 | 6.80% | ||||
| head, body and Thigh | 20 | 17 | 4.40% | 17 | 13 (4.7) | 3 (4.5) | 1 (2.9) |
| TOTAL | 520 | 378 | 378 | 277 | 67 | 34 |
| Species and Number (%) of Resistant Isolates | |||||
|---|---|---|---|---|---|
| Antimicrobial Class | Antimicrobials | C. coli (n = 277) | C. jejuni (n = 67) | Others (n = 34) | Total (n = 378) |
| Aminoglycosides | Gentamycin | 34 (12.2) | 8 (11.9) | 2 (5.8) | 44 (11.6) |
| Streptomycin | 240 (86.6) | 62 (92.5) | 31 (91.1) | 333 (88.0) | |
| Macrolides | Erythromycin | 247 (89.1) | 66 (98.5) | 30 (88.2) | 343 (90.7) |
| Penicillins | Ampicillin | 191 (68.9) | 57 (85.0) | 28 (82.3) | 276 (73.0) |
| Quinolones | Ciprofloxacin | 148 (53.4) | 45 (67.1) | 23 (67.6) | 216 (57.1) |
| Nalidixic acid | 77 (27.7) | 18 (26.8) | 8 (23.5) | 103 (27.2) | |
| Tetracyclines | Tetracycline | 227 (81.9) | 64 (95.5) | 28 (82.3) | 319 (84.3) |
| No. of Resistance | Resistance Patterns | Number of Isolates Per Sample Type | C. coli (n = 277) | C. jejuni (n = 67) | Other (n = 34) | Total (n = 378) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | S | L | TC | CR | CS | C | RM | ||||||
| 7 | AMP+NAL+CIP+ERY+TET+GEN+STR | 14 | 2 | 10 | 5 | 1 | 16 | ||||||
| TOTAL | 10 | 5 | 1 | 16 | |||||||||
| 6 | AMP+NAL+CIP+ERY+TET+STR | 16 | 3 | 1 | 3 | 1 | 1 | 17 | 7 | 1 | 25 | ||
| NAL+CIP+ERY+TET+GEN+STR | 2 | 2 | 1 | 5 | 0 | 0 | 5 | ||||||
| AMP+CIP+ERY+TET+GEN+STR | 13 | 1 | 11 | 2 | 1 | 14 | |||||||
| AMP+NAL+ERY+TET+GEN+STR | 1 | 1 | 0 | 0 | 1 | ||||||||
| TOTAL | 34 | 9 | 2 | 45 | |||||||||
| 5 | AMP+CIP+ERY+TET+STR | 55 | 9 | 4 | 4 | 1 | 2 | 15 | 1 | 56 | 25 | 13 | 94 |
| AMP+NAL+ERY+TET+STR | 5 | 1 | 4 | 1 | 1 | 6 | |||||||
| AMP+NAL+CIP+ERY+STR | 2 | 2 | 4 | 0 | 0 | 4 | |||||||
| NAL+CIP+ERY+TET+STR | 2 | 1 | 1 | 2 | 0 | 3 | |||||||
| NAL+CIP+TET+GEN+STR | 1 | 1 | 0 | 0 | 1 | ||||||||
| TOTAL | 66 | 28 | 14 | 108 | |||||||||
| 4 | ERY+TET+GEN+STR | 2 | 2 | 0 | 0 | 2 | |||||||
| AMP+ERY+TET+STR | 37 | 7 | 17 | 4 | 1 | 8 | 1 | 51 | 15 | 6 | 72 | ||
| CIP+ERY+TET+STR | 2 | 2 | 1 | 5 | 9 | 0 | 1 | 10 | |||||
| AMP+CIP+ERY+STR | 5 | 7 | 9 | 1 | 2 | 12 | |||||||
| NAL+ERY+TET+STR | 1 | 1 | 0 | 0 | 1 | ||||||||
| NAL+CIP+ERY+TET | 2 | 1 | 1 | 3 | 1 | 0 | 4 | ||||||
| AMP+NAL+ERY+TET | 2 | 1 | 1 | 0 | 2 | ||||||||
| NAL+CIP+ERY+STR | 1 | 0 | 0 | 1 | 1 | ||||||||
| AMP+NAL+ERY+STR | 5 | 6 | 10 | 0 | 1 | 11 | |||||||
| NAL+CIP+TET+STR | 1 | 11 | 1 | 11 | 0 | 2 | 13 | ||||||
| TOTAL | 97 | 18 | 13 | 128 | |||||||||
| 3 | AMP+ERY+TET | 2 | 1 | 2 | 0 | 1 | 3 | ||||||
| NAL+ERY+TET | 1 | 1 | 0 | 0 | 1 | ||||||||
| ERY+TET+STR | 9 | 2 | 9 | 2 | 0 | 11 | |||||||
| AMP+ERY+STR | 1 | 3 | 7 | 1 | 12 | 0 | 0 | 12 | |||||
| CIP+ERY+STR | 2 | 1 | 1 | 0 | 2 | ||||||||
| AMP+CIP+ERY | 1 | 1 | 0 | 0 | 1 | ||||||||
| ERY+TET+GEN | 1 | 1 | 0 | 0 | 1 | ||||||||
| CIP+ERY+TET | 1 | 1 | 2 | 0 | 0 | 2 | |||||||
| TOTAL | 29 | 3 | 1 | 33 | |||||||||
| Total MDR isolates | 236 | 63 | 31 | 330 | |||||||||
| Percentage MDR | 85.1% | 94.0% | 91.1% | ||||||||||
| None MDR | 41 | 4 | 3 | 48 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sithole, V.; Amoako, D.G.; Abia, A.L.K.; Perrett, K.; Bester, L.A.; Essack, S.Y. Occurrence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter spp. in Intensive Pig Production in South Africa. Pathogens 2021, 10, 439. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10040439
Sithole V, Amoako DG, Abia ALK, Perrett K, Bester LA, Essack SY. Occurrence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter spp. in Intensive Pig Production in South Africa. Pathogens. 2021; 10(4):439. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10040439
Chicago/Turabian StyleSithole, Viwe, Daniel Gyamfi Amoako, Akebe Luther King Abia, Keith Perrett, Linda A. Bester, and Sabiha Y. Essack. 2021. "Occurrence, Antimicrobial Resistance, and Molecular Characterization of Campylobacter spp. in Intensive Pig Production in South Africa" Pathogens 10, no. 4: 439. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10040439