
Transcriptional Regulation of the Multiple Resistance Mechanisms in Salmonella—A Review

 , , , and
, , , and
Abstract
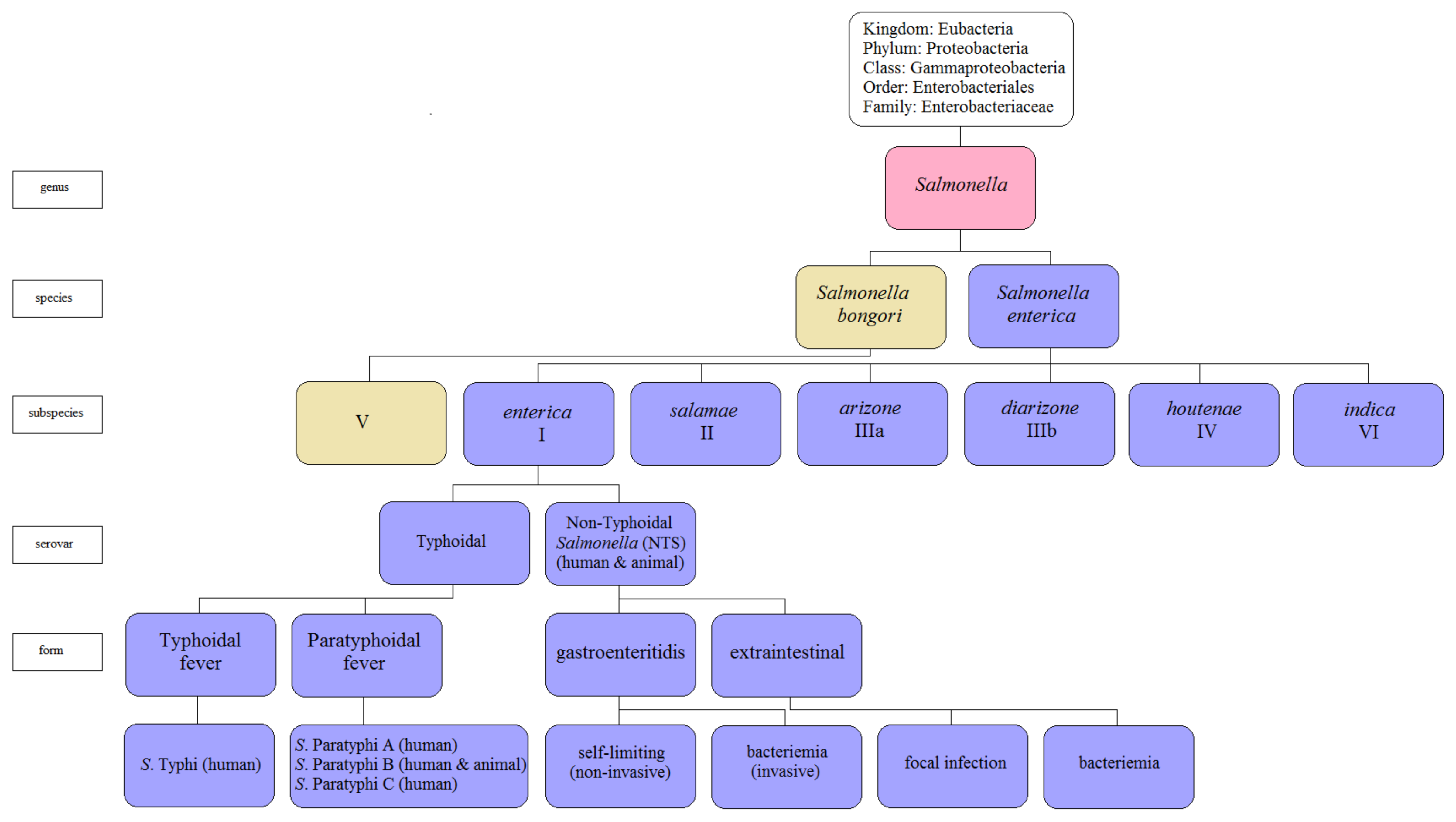
:1. Introduction
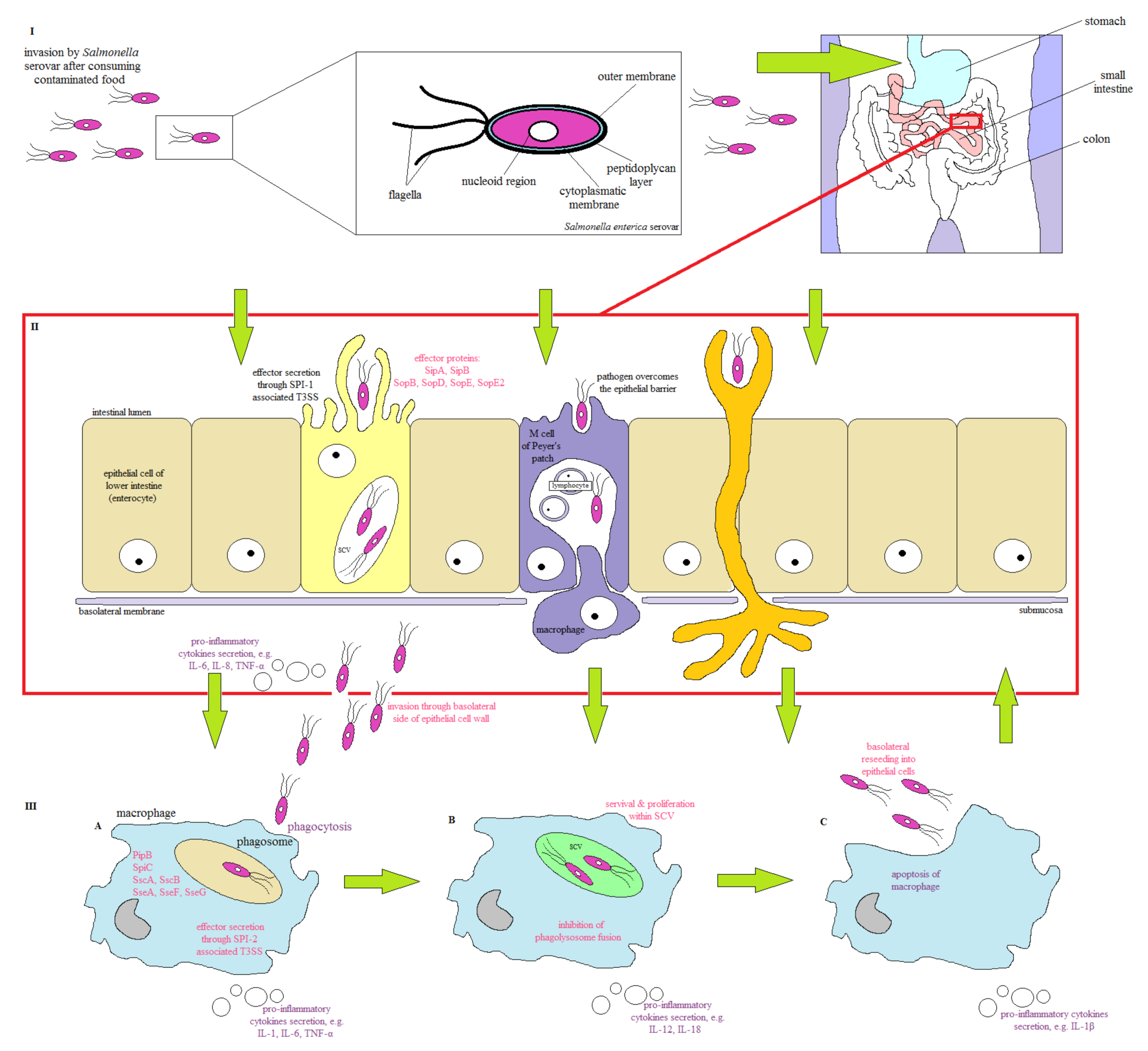
2. Salmonella Pathogenesis
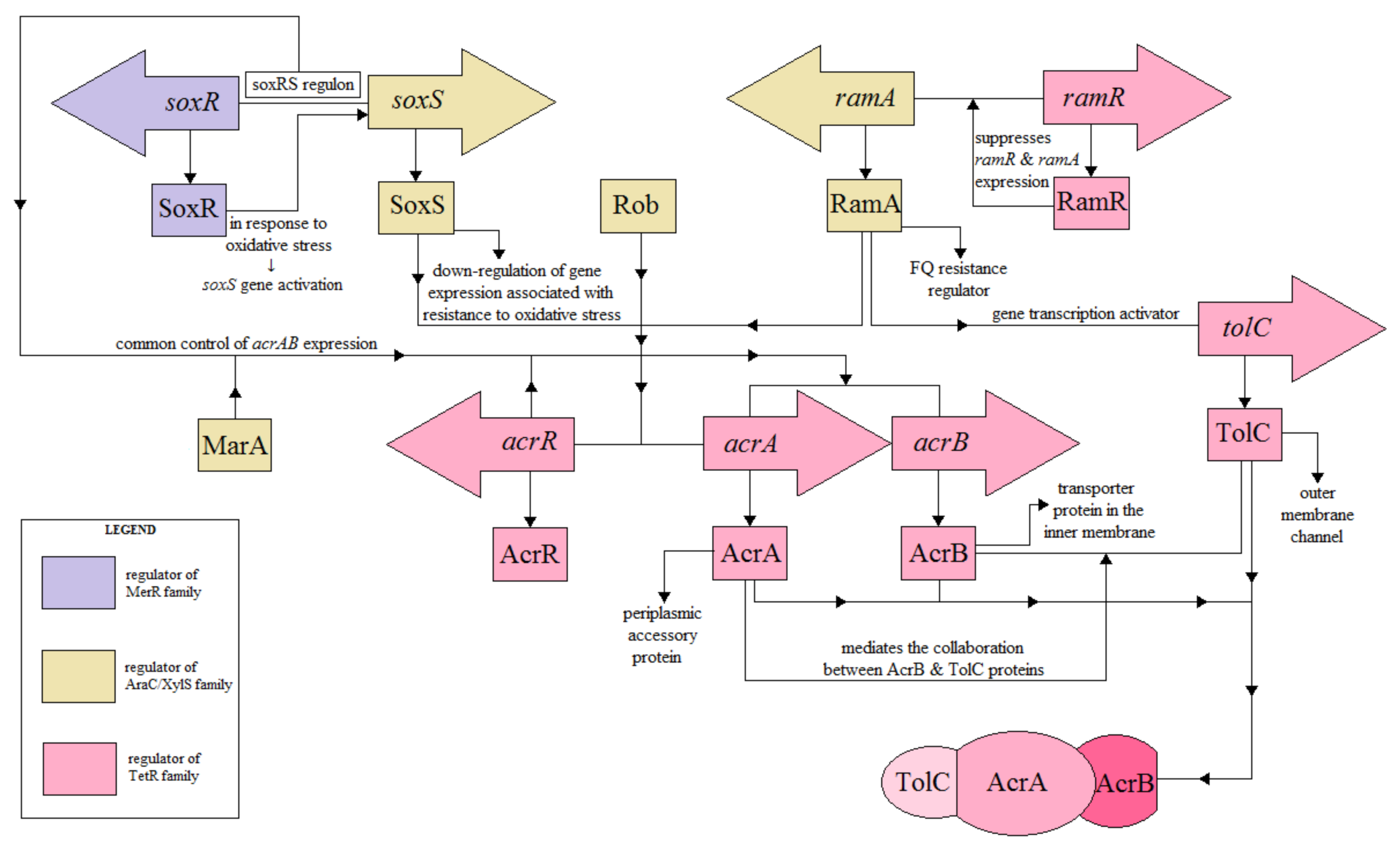
3. Molecular Systems of Multiple Resistance in Salmonella
3.1. The TetR Family

3.2. The AraC/XylS Family
3.3. Two-Component Signal Transduction Systems
3.3.1. The PhoP-PhoQ System
3.3.2. The CpxAR System
3.3.3. The BaeSR System
3.3.4. The EnvZ/OmpR System
3.3.5. The PmrAB System
3.4. The MerR Family
3.5. The MarR Family
3.6. The Histone-Like Protein Family (H-NS)
3.7. The LysR Family
3.7.1. SpvR
3.7.2. LeuO
3.7.3. LoiA
3.7.4. STM0030
3.7.5. CysB
3.7.6. LtrR
3.7.7. Hrg
4. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eng, S.K.; Pusparajah, P.; Mutalib, N.S.A.; Ser, H.L.; Chan, K.G.; Lee, L.H. Salmonella: A review on pathogenesis epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Heredia, N.; García, S. Animals as sources of food-borne pathogens: A review. Anim. Nutr. 2018, 4, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-analysis. Appl. Environ. Microb. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Biswas, S.; Paudyal, N.; Pan, H.; Li, X.; Fang, W.; Yue, M. Antibiotic Resistance in Salmonella Typhimurium Isolates Recovered From the Food Chain Through National Antimicrobial Resistance Monitoring System Between 1996 and 2016. Front. Microbiol. 2019, 10, 985. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, A.M.P.; Ferrari, R.G.; Conte-Junior, C.A. Virulence Factors in Salmonella Typhimurium: The Sagacity of a Bacterium. Curr. Microbiol. 2018, 76, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yi, W.; Yao, Y.; Zhu, N.; Qin, P. Characteristic diversity and antimicrobial resistance of Salmonella from gastroenteritis. J. Infect. Chemother. 2018, 24, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.R.; Goggins, J.A.; McLachlan, J.B. Salmonella infection: Interplay between the bacteria and host immune system. Immunol. Lett. 2017, 190, 42–50. [Google Scholar] [CrossRef]
- Shaheen, A.; Tariq, A.; Shehzad, A.; Iqbal, M.; Mirza, O.; Maslov, D.A.; Rahman, M. Transcriptional regulation of drug resistance mechanisms in Salmonella: Where we stand and what we need to know. World J. Microbiol. Biotechnol. 2020, 36, 85. [Google Scholar] [CrossRef]
- Wemyss, M.A.; Pearson, J.S. Host Cell Death Responses to Non-typhoidal Salmonella Infection. Front. Immunol. 2019, 10, 1758. [Google Scholar] [CrossRef]
- Kingsley, R.A.; Bäumler, A.J. Host adaptation and the emergence of infectious disease: The Salmonella paradigm. Mol. Microbiol. 2000, 36, 1006–1014. [Google Scholar] [CrossRef]
- Huang, H.; Sun, Y.; Yuan, L.; Pan, Y.; Gao, Y.; Ma, C.; Hu, G. Regulation of the Two-Component Regulator CpxR on Aminoglycosides and β-lactams Resistance in Salmonella enterica serovar Typhimurium. Front. Microbiol. 2016, 7, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, D.; McCusker, M.P.; Fanning, S.; Martins, M. Salmonella—Host interactions—Modulation of the host innate immune system. Front. Immunol. 2014, 5, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Połaska, M.; Sokołowska, B. Bacteriophages—A new hope or huge problem in the food industry. AIMS Microbiol. 2019, 5, 324–346. [Google Scholar] [CrossRef] [PubMed]
- Wójcicki, M.; Błażejak, S.; Gientka, I.; Brzezicka, K. The concept of using bacteriophages to improve the microbiological quality of minimally-processed foods. Acta Sci. Pol. Technol. Aliment. 2019, 18, 373–383. [Google Scholar] [CrossRef]
- Vargiu, A.V.; Pos, K.M.; Poole, K.; Nikaido, H. Editorial: Bad Bugs in the XXIst Century: Resistance Mediated by Multi-Drug Efflux Pumps in Gram-Negative Bacteria. Front. Microbiol. 2016, 7, 833. [Google Scholar] [CrossRef] [Green Version]
- Debroy, R.; Miryala, S.K.; Naha, A.; Anbarasu, A.; Ramaiah, S. Gene interaction network studies to decipher the multi-drug resistance mechanism in Salmonella enterica serovar Typhi CT18 reveal potential drug targets. Microb. Pathog. 2020, 142, 104096. [Google Scholar] [CrossRef]
- Weston, N.; Sharma, P.; Ricci, V.; Piddock, L.J.V. Regulation of the AcrAB-TolC efflux pump in Enterobacteriaceae. Res. Microbiol. 2018, 169, 425–431. [Google Scholar] [CrossRef]
- Blair, J.M.; Richmond, G.E.; Piddock, L.J. Multidrug efflux pumps in Gram-negative bacteria and their role in antibiotic resistance. Future Microbiol. 2014, 9, 1165–1177. [Google Scholar] [CrossRef]
- Alav, I.; Sutton, M.J.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, S.; Nikaido, E.; Nakashima, R.; Sakurai, K.; Fujiwara, D.; Fujii, I.; Nishino, K. The crystal structure of multidrug-resistance regulator RamR with multiple drugs. Nat. Commun. 2013, 4, 2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, A.E.; Yan, M.B.; Tran, M.; Wright, N.; Luzader, D.H.; Kendall, M.M.; Ruiz-Perez, F.; Nataro, J.P. A large family of anti-activators accompanying XylS/AraC family regulatory proteins. Mol. Microbiol. 2016, 101, 314–332. [Google Scholar] [CrossRef]
- Zschiedrich, C.P.; Keidel, V.; Szurmant, H. Molecular mechanisms of two-component signal transduction. J. Mol. Biol. 2016, 428, 3752–3775. [Google Scholar] [CrossRef] [Green Version]
- Mattos-Graner, R.O.; Duncan, M.J. Two-component signal transduction systems in oral bacteria. J. Oral Microbiol. 2017, 9, 1400858. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Sun, D.; Zhu, J.; Liu, W. Two-Component Signal Transduction Systems: A Major Strategy for Connecting Input Stimuli to Biofilm Formation. Front. Microbiol. 2019, 9, 3279. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, H.; Liu, Y.; Lv, R.; Liu, X.; Song, X.; Wang, J.; Jiang, L. SoxS is a positive regulator of key pathogenesis genes and promotes intracellular replication and virulence of Salmonella Typhimurium. Microb. Pathog. 2020, 139, 103925. [Google Scholar] [CrossRef]
- Banda, M.M.; Zavala-Alvarado, C.; Pérez-Morales, D.; Bustamante, V.H. SlyA and HilD Counteract H-NS-Mediated Repression on the ssrAB Virulence Operon of Salmonella enterica Serovar Typhimurium and Thus Promote Its Activation by OmpR. J. Bacteriol. 2019, 201, e00530-18. [Google Scholar] [CrossRef] [Green Version]
- Zha, L.; Garrett, S.; Sun, J. Salmonella Infection in Chronic Inflammation and Gastrointestinal Cancer. Diseases 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, A.D.; Slauch, J.M. SPECIAL COLLECTION Advancing Knowledge of Biological Mechanisms of the Superorganism for Next Generation (NextGen) Microbial Dose-Response Modeling: Salmonellosis 3. Mechanisms of Salmonella pathogenesis in animal models. Hum. Ecol. Risk Assess. 2017, 1877–1892. [Google Scholar] [CrossRef] [PubMed]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Salmonella infection—Prevention and treatment by antibiotics and probiotic yeasts: A review. Microbiology 2018, 164, 1327–1344. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Schwerk, P.; Tedin, K. Rapid Isolation of intact Salmonella-containing vacuoles using paramagnetic nanoparticles. Gut Pathog. 2018, 10, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero-Fresno, A.; Olsen, J.E. Salmonella Typhimurium metabolism affects virulence in the host—A mini-review. Food Microbiol. 2018, 71, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella Pathogenicity and Host Adaptation in Chicken-Associated Serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Zhang, H.; Ma, S.; Song, Y.; Lv, R.; Liu, X.; Yang, B.; Huang, D.; Jiang, L. Transcriptome analysis of virulence gene regulation by the ATP-dependent Lon protease in Salmonella Typhimurium. Future Microbiol. 2019, 14, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Jepson, M.A.; Clark, M.A. The role of M cells in Salmonella infection. Microbes Infect. 2001, 3, 1183–1190. [Google Scholar] [CrossRef]
- Tahoun, A.; Mahajan, S.; Paxton, E.; Malterer, G.; Donaldson, D.S.; Wang, D.; Tan, A.; Gillespie, T.L.; O’Shea, M.; Roe, A.J.; et al. Salmonella Transforms Follicle-Associated Epithelial Cells into M Cells to Promote Intestinal Invasion. Cell Host Microbe 2012, 12, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Andesfha, E.; Indrawati, A.; Mayasari, N.L.P.I.; Rahayuningtyas, I.; Jusa, I. Detection of Salmonella pathogenicity island and Salmonella plasmid virulence genes in Salmonella Enteritidis originated from layer and broiler farms in Java Island. J. Adv. Vet. Anim. Res. 2019, 6, 384–393. [Google Scholar] [CrossRef]
- Song, X.; Zhang, H.; Liu, X.; Yuan, J.; Wang, P.; Lv, R.; Yang, B.; Huang, D.; Jiang, L. The putative transcriptional regulator STM14_3563 facilitates Salmonella Typhimurium pathogenicity by activating virulence-related genes. Int. Microbiol. 2019, 23, 381–390. [Google Scholar] [CrossRef]
- Ilyas, B.; Tsai, C.N.; Coombes, B.K. Evolution of Salmonella-Host Cell Interactions through a Dynamic Bacterial Genome. Front. Cell Infect. Microbiol. 2017, 7, 428. [Google Scholar] [CrossRef]
- Jiang, X.; Li, X.; Sun, S.; Jiang, L. The transcriptional regulator VarN contributes to Salmonella Typhimurium growth in macrophages and virulence in mice. Res. Microbiol. 2018, 169, 214–221. [Google Scholar] [CrossRef]
- Zhang, H.; Song, X.; Lv, R.; Liu, X.; Wang, P.; Jiang, L. The LysR-type transcriptional regulator STM0030 contributes to Salmonella Typhimurium growth in macrophages and virulence in mice. J. Basic Microbiol. 2019, 59, 1143–1153. [Google Scholar] [CrossRef]
- Raffatellu, M.; Wilson, R.P.; Chessa, D.; Andrews-Polymenis, H.; Tran, Q.T.; Lawhon, S.; Khare, S.; Adams, L.G.; Bäumler, A.J. SipA, SopA, SopB, SopD, and SopE2 contribute to Salmonella enterica serotype Typhimurium invasion of epithelial cells. Infect. Immun. 2005, 73, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Ibarra, J.A.; Knodler, L.A.; Sturdevant, D.E.; Virtaneva, K.; Carmody, A.B.; Fischer, E.R.; Porcella, S.F.; Steele-Mortimer, O. Induction of Salmonella pathogenicity island 1 under different growth conditions can affect Salmonella-host cell interactions in vitro. Microbiology 2010, 156, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- Figueira, R.; Watson, K.G.; Holden, D.W.; Helaine, S. Identification of Salmonella pathogenicity island-2 type III secretion system effectors involved in intramacrophage replication of S. enterica serovar Typhimurium: Implications for rational vaccine design. mBio 2013, 4, e00065-13. [Google Scholar] [CrossRef] [Green Version]
- Rychlik, I.; Karasova, D.; Sebkova, A.; Volf, J.; Sisak, F.; Havlickova, H.; Kummer, V.; Imre, A.; Szmolka, A.; Nagy, B. Virulence potential of five major pathogenicity islands (SPI-1 to SPI-5) of Salmonella enterica serovar Enteritidis for chickens. BMC Microbiol. 2009, 9, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sana, T.G.; Flaugnatti, N.; Lugo, K.A.; Lam, L.H.; Jacobson, A.; Baylot, V.; Durand, E.; Journet, L.; Cascales, E.; Monack, D.M. Salmonella Typhimurium utilizes a T6SS-mediated antibacterial weapon to establish in the host gut. Proc. Natl. Acad. Sci. USA 2016, 113, E5044–E5051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liaquat, S.; Sarwar, Y.; Ali, A.; Haque, A.; Farooq, M.; Martinez-Ballesteros, I.; Laorden, L.; Garaizar, J.; Bikandi, J. Virulotyping of Salmonella enterica serovar Typhi isolates from Pakistan: Absence of complete SPI-10 in Vi negative isolates. PLoS Neglect. Trop. Dis. 2018, 12, e0006839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xian, H.; Yuan, Y.; Yin, C.; Wang, Z.; Ji, R.; Chu, C.; Jiao, X.; Li, Q. The SPI-19 encoded T6SS is required for Salmonella Pullorum survival within avian macrophages and initial colonization in chicken dependent on inhibition of host immune response. Vet. Microbiol. 2020, 250, 108867. [Google Scholar] [CrossRef]
- Jiang, L.; Feng, L.; Yang, B.; Zhang, W.; Wang, P.; Jiang, X.; Wang, L. Signal transduction pathway mediated by the novel regulator LoiA for low oxygen tension induced Salmonella Typhimurium invasion. PLoS Pathog. 2017, 13, e1006429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, A.D.S.; Dorman, C.J. A Fundamental Regulatory Mechanism Operating through OmpR and DNA Topology Controls Expression of Salmonella Pathogenicity Islands SPI-1 and SPI-2. PLoS Genet. 2012, 8, e1002615. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504–521. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.N.; MacNair, C.R.; Cao, M.P.T.; Perry, J.N.; Magolan, J.; Brown, E.D.; Coombes, B.K. Targeting Two-Component Systems Uncovers a Small-Molecule Inhibitor of Salmonella Virulence. Cell Chem. Biol. 2020, 27, 793–805. [Google Scholar] [CrossRef]
- Foley, S.L.; Lynne, A.M. Food animal-associated Salmonella challenges: Pathogenicity and antimicrobial resistance. J. Anim. Sci. 2008, 86, 173–187. [Google Scholar] [CrossRef]
- Fricke, W.F.; McDermott, P.F.; Mammel, M.K.; Zhao, S.; Johnson, T.J.; Rasko, D.A.; Fedorka-Cray, P.J.; Pedroso, A.; Whichard, J.M.; LeClerc, J.E.; et al. Antimicrobial Resistance-Conferring Plasmids with Similarity to Virulence Plasmids from Avian Pathogenic Escherichia coli Strains in Salmonella enterica Serovar Kentucky Isolates from Poultry. Appl. Environ. Microb. 2009, 75, 5963–5971. [Google Scholar] [CrossRef] [Green Version]
- Chopra, A.K.; Huang, J.H.; Xu, X.J.; Burden, K.; Niesel, D.W.; Rosenbaum, M.W.; Popov, V.L.; Peterson, J.W. Role of Salmonella enterotoxin in overall virulence of the organism. Microb. Pathog. 1999, 27, 155–171. [Google Scholar] [CrossRef]
- Hung, C.C.; Garner, C.D.; Slauch, J.M.; Dwyer, Z.W.; Lawhon, S.D.; Frye, J.G.; McClelland, M.; Ahmer, B.M.M.; Altier, C. The intestinal fatty acid propionate inhibits Salmonella invasion through the post-translational control of HilD. Mol. Microbiol. 2013, 87, 1045–1060. [Google Scholar] [CrossRef] [Green Version]
- Golubeva, Y.A.; Ellermeier, J.R.; Cott Chubiz, J.E.; Slauch, J.M. Intestinal Long-Chain Fatty Acids Act as a Direct Signal To Modulate Expression of the Salmonella Pathogenicity Island 1 Type III Secretion System. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosire, E.M.; Eade, C.R.; Schiltz, C.; Reid, A.J.; Troutman, J.; Chappie, J.S.; Altier, C. Diffusible signal factors act through AraC-type transcriptional regulators as chemical cues to repress virulence of enteric pathogens. Infect. Immun. 2020, 88, e00226-20. [Google Scholar] [CrossRef] [PubMed]
- Thota, S.S.; Chubiz, L.M. Multidrug Resistance Regulators MarA, SoxS, Rob, and RamA Repress Flagellar Gene Expression and Motility in Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2019, 201, e00385-19. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Cazares, N.; García-Contreras, R.; Pérez-López, M.; Castillo-Juárez, I. Phenolic compounds with anti-virulence properties. In Phenolic Compounds—Biological Activity; Marcos Soto-Hernandez, M., Palma-Tenango, M., Del Rosario Garcia-Mateos, M., Eds.; IntechOpen: London, UK, 2017; pp. 139–167. [Google Scholar]
- Rütschlin, S.; Böttcher, T. Inhibitors of Bacterial Swarming Behavior. Chem. Eur. J. 2020, 26, 964–979. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Sheng, X.; Zhang, H.; Zou, X.; Ni, B.; Xu, S.; Zhu, X.; Xu, H.; Huang, X. RpoE may Promote Flagellar Gene Expression in Salmonella enterica Serovar Typhi Under Hyperosmotic Stress. Curr. Microbiol. 2011, 62, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Osterman, I.A.; Dikhtyar, Y.Y.; Bogdanov, A.A.; Dontsova, O.A.; Sergiev, P.V. Regulation of flagellar gene expression in Bacteria. Biochemistry 2015, 80, 1447–1456. [Google Scholar] [CrossRef]
- Singer, H.M.; Kuhne, C.; Deditius, J.A.; Hughes, K.T.; Erhardt, M. The Salmonella Spi1 Virulence Regulatory Protein HilD Directly Activates Transcription of the Flagellar Master Operon flhDC. J. Bacteriol. 2014, 196, 1448–1457. [Google Scholar] [CrossRef] [Green Version]
- Doyle, M.E. Multidrug-Resistant Pathogens in the Food Supply. Foodborne Pathog. Dis. 2015, 12, 261–279. [Google Scholar] [CrossRef]
- Brunelle, B.W.; Bearson, B.L.; Bearson, S.M.D.; Casey, T.A. Multidrug-Resistant Salmonella enterica Serovar Typhimurium Isolates Are Resistant to Antibiotics That Influence Their Swimming and Swarming Motility. mSphere 2017, 2, e00306-17. [Google Scholar] [CrossRef] [Green Version]
- Nazek, A.G.; Ridha, B.A. Fluoroquinolones Resistance Salmonella: State of Knowledge. Adv. Biotechnol. Microbiol. 2017, 7, 555717. [Google Scholar] [CrossRef]
- Zhang, C.Z.; Ren, S.Q.; Chang, M.X.; Chen, P.X.; Ding, H.Z.; Jiang, H.X. Resistance mechanisms and fitness of Salmonella Typhimurium and Salmonella Enteritidis mutants evolved under selection with ciprofloxacin in vitro. Sci. Rep. 2017, 7, 9113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fàbrega, A.; Ballesté-Delpierre, C.; Vila, J. Differential impact of ramRA mutations on both ramA transcription and decreased antimicrobial susceptibility in Salmonella Typhimurium. J. Antimicrob. Chemother. 2016, 71, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velhner, M. Mechanisms of Resistance to Quinolones and Epidemiological Significance of Salmonella spp. Acta Vet. 2016, 66, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Cuypers, W.L.; Jacobs, J.; Wong, V.; Klemm, E.J.; Deborggraeve, S.; Puyvelde, S.V. Fluoroquinolone resistance in Salmonella: Insights by whole-genome sequencing. Microbial Genom. 2018, 4, e000195. [Google Scholar] [CrossRef]
- Zhang, C.Z.; Zhang, Y.; Ding, X.M.; Lin, X.L.; Lian, X.L.; Trampari, E.; Thomson, N.M.; Ding, H.Z.; Webber, M.A.; Jiang, H.X. Emergence of ciprofloxacin heteroresistance in foodborne Salmonella enterica serovar Agona. J. Antimicrob. Chemother. 2020, 75, 2773–2779. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Reynolds, J.; Karp, B.E.; Tate, H.; Fedorka-Cray, P.J.; Plumblee, J.R.; Hoekstra, R.M.; Whichard, J.M.; Mahon, B.E. Ceftriaxone-Resistant Nontyphoidal Salmonella from Humans, Retail Meats, and Food Animals in the United States, 1996–2013. Foodborne Pathog. Dis. 2017, 14, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, S.; López-Escarpa, D.; Pucciarelli, M.G.; Cestero, J.J.; Baquero, F.; García-del Portillo, F. An alternative penicillin-binding protein involved in Salmonella relapses following ceftriaxone therapy. EBioMedicine 2020, 55, 102771. [Google Scholar] [CrossRef] [PubMed]
- Thilliez, G.; Kingsley, R.A. Salmonella intracellular adaptation is key to understand cephalosporin treatment relapse. EBioMedicine 2020, 56, 102802. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.; Martínez-Puchol, S.; Palma, N.; Horna, G.; Ruiz-Roldán, L.; Pons, M.J.; Ruiz, J. Macrolide resistance mechanisms in Enterobacteriaceae: Focus on azithromycin. Crit. Rev. Microbiol. 2016, 43, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Al Kraiem, A.A.; Yang, G.; Al Kraiem, F.; Chen, T. Challenges associated with ceftriaxone resistance in Salmonella. Front. Life Sci. 2018, 11, 26–34. [Google Scholar] [CrossRef]
- Djeghout, B.; Saha, S.; Sajib, M.S.I.; Tanmoy, A.M.; Islam, M.; Kay, G.L.; Langridge, G.C.; Endtz, H.P.; Wain, J.; Saha, S.K. Ceftriaxone-resistant Salmonella Typhi carries an IncI1-ST31 plasmid encoding CTX-M-15. J. Med. Microbiol. 2018, 67, 620–627. [Google Scholar] [CrossRef]
- Opperman, T.J.; Nguyen, S.T. Recent advances toward a molecular mechanism of efflux pump inhibition. Front. Microbiol. 2015, 6, 421. [Google Scholar] [CrossRef] [Green Version]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Piddock, L.J.V. Understanding the basis of antibiotic resistance: A platform for drug discovery. Microbiology 2014, 160, 2366–2373. [Google Scholar] [CrossRef] [Green Version]
- Ramos, J.L.; Martinez-Bueno, M.; Molina-Henares, A.J.; Teran, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR Family of Transcriptional Repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [Green Version]
- Colclough, A.L.; Scadden, J.; Blair, J.M.A. TetR-family transcription factors in Gram-negative bacteria: Conservation, variation and implications for efflux-mediated antimicrobial resistance. BMC Genom. 2019, 20, 731. [Google Scholar] [CrossRef] [PubMed]
- Baucheron, S.; Coste, F.; Canepa, S.; Maurel, M.C.; Giraud, E.; Culard, F.; Castaing, B.; Roussel, A.; Cloeckaert, A. Binding of the RamR repressor to wild-type and mutated promoters of the ramA gene involved in efflux-mediated multidrug resistance in Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2012, 56, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Ritter, A.C.; Tondo, E.C.; Siqueira, F.M.; Soggiu, A.; Varela, A.P.M.; Mayer, F.Q.; Brandelli, A. Genome analysis reveals insights into high-resistance and virulence of Salmonella Enteritidis involved in foodborne outbreaks. Int. J. Food Microbiol. 2019, 306, 108269. [Google Scholar] [CrossRef] [PubMed]
- Chinni, S.V.; Raabe, C.A.; Zakaria, R.; Randau, G.; Hoe, C.H.; Zemann, A.; Brosius, J.; Tang, T.H.; Rozhdestvensky, T.S. Experimental identification and characterization of 97 novel npcRNA candidates in Salmonella enterica serovar Typhi. Nucleic Acids Res. 2010, 38, 5893–5908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baucheron, S.; Nishino, K.; Monchaux, I.; Canepa, S.; Maurel, M.C.; Coste, F.; Roussel, A.; Cloeckaert, A.; Giraud, E. Bile-mediated activation of the acrAB and tolC multidrug efflux genes occurs mainly through transcriptional derepression of ramA in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2014, 69, 2400–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Cui, S.; Meng, J. Effect of transcriptional activators RamA and SoxS on expression of multidrug efflux pumps AcrAB and AcrEF in fluoroquinolone-resistant Salmonella Typhimurium. J. Antimicrob. Chemother. 2009, 63, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, S.; Nakashima, R.; Sakurai, K.; Baucheron, S.; Giraud, E.; Doublet, B.; Cloeckaert, A.; Nishino, K. Crystal structure of the multidrug resistance regulator RamR complexed with bile acids. Sci. Rep. 2019, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, E.; Yamaguchi, A.; Nishino, K. AcrAB Multidrug Efflux Pump Regulation in Salmonella enterica serovar Typhimurium by RamA in Response to Environmental Signals. J. Biol. Chem. 2008, 283, 24245–24253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youlden, G.H.; Ricci, V.; Wang-Kan, X.; Piddock, L.J.V.; Jabbari, S.; King, J.R. Time dependent asymptotic analysis of the gene regulatory network of the AcrAB-TolC efflux pump system in gram-negative bacteria. J. Math. Biol. 2021, 82, 31. [Google Scholar] [CrossRef]
- Su, C.C.; Rutherford, D.J.; Yu, E.W. Characterization of the multidrug efflux regulator AcrR from Escherichia coli. Biochem. Biophys. Res. Commun. 2007, 361, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Darzynkiewicz, Z.M.; Green, A.T.; Abdali, N.; Hazel, A.; Fulton, R.L.; Kimball, J.; Gryczynski, Z.; Gumbart, J.C.; Parks, J.M.; Smith, J.C.; et al. Identification of Binding Sites for Efflux Pump Inhibitors of the Escherichia coli AcrAB-TolC component AcrA. Biophys. J. 2019, 116, 648–658. [Google Scholar] [CrossRef] [Green Version]
- Grimsey, E.M.; Weston, N.; Ricci, V.; Stone, J.W.; Piddock, L.J.V. Overexpression of RamA, Which Regulates Production of the Multidrug Resistance Efflux Pump AcrAB-TolC, Increases Mutation Rate and Influences Drug Resistance Phenotype. Antimicrob. Agents Chemother. 2020, 64, e02460-19. [Google Scholar] [CrossRef] [PubMed]
- Ricci, V.; Tzakas, P.; Buckley, A.; Coldham, N.C.; Piddock, L.J.V. Ciprofloxacin-Resistant Salmonella enterica Serovar Typhimurium Strains Are Difficult To Select in the Absence of AcrB and TolC. Antimicrob. Agents Chemother. 2006, 50, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, E.; Shirosaka, I.; Yamaguchi, A.; Nishino, K. Regulation of the AcrAB multidrug efflux pump in Salmonella enterica serovar Typhimurium in response to indole and paraquat. Microbiology 2011, 157, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Manjasetty, B.A.; Halavaty, A.S.; Luan, C.H.; Osipiuk, J.; Mulligan, R.; Kwon, K.; Anderson, W.F.; Joachimiak, A. Loop-to-helix transition in the structure of multidrug regulator AcrR at the entrance of the drug-binding cavity. J. Struct. Biol. 2016, 194, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Regan, E.; Quinn, T.; Pages, J.M.; McCusker, M.; Piddock, L.; Fanning, S. Multiple Regulatory Pathways Associated with High-Level Ciprofloxacin and Multidrug Resistance in Salmonella enterica Serovar Enteritidis: Involvement of ramA and Other Global Regulators. Antimicrob. Agents Chemother. 2009, 53, 1080–1087. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Z.; Chen, P.X.; Yang, L.; Li, W.; Chang, M.X.; Jiang, H.X. Coordinated Expression of acrAB-tolC and Eight Other Functional Efflux Pumps through Activating ramA and marA in Salmonella enterica serovar Typhimurium. Microb. Drug Resist. 2018, 24, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, A. Antibiotic resistome of Salmonella typhi: Molecular determinants for the emergence of drug resistance. Front. Med. 2021, 43. [Google Scholar] [CrossRef]
- King, A.N.; de Mets, F.; Brinsmade, S.R. Who’s in control? Regulation of metabolism and pathogenesis in space and time. Curr. Opin. Microbiol. 2020, 55, 88–96. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, X.; Zhang, D.; Lu, C. Myricanol Inhibits the Type III Secretion System of Salmonella enterica Serovar Typhimurium by Interfering With the DNA-Binding Activity of HilD. Front. Microbiol. 2020, 11, 571217. [Google Scholar] [CrossRef]
- Ferrari, R.G.; Galiana, A.; Cremades, R.; Rodríguez, J.C.; Magnani, M.; Tognim, M.C.B.; Oliveira, T.C.; Royo, G. Expression of the marA, soxS, acrB and ramA genes related to the AcrAB/TolC efflux pump in Salmonella enterica strains with and without quinolone resistance-determining regions gyrA gene mutations. Braz. J. Infect. Dis. 2013, 17, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narm, K.E.; Kalafatis, M.; Slauch, J.M. HilD, HilC, and RtsA Form Homodimers and Heterodimers To Regulate Expression of the Salmonella Pathogenicity Island I Type III Secretion System. J. Bacteriol. 2020, 202, e00012-20. [Google Scholar] [CrossRef] [PubMed]
- Miletic, S.; Goessweiner-Mohr, N.; Marlovits, T.C. The Structure of the Type III Secretion System Needle Complex. Curr. Top. Microbiol. 2020, 427, 67–90. [Google Scholar] [CrossRef]
- Gil, F.; Ipinza, F.; Fuentes, J.; Fumeron, R.; Villarreal, J.M.; Aspée, A.; Mora, G.C.; Vásquez, C.C.; Saavedra, C. The ompW (porin) gene mediates methyl viologen (paraquat) efflux in Salmonella enterica serovar Typhimurium. Res. Microbiol. 2007, 158, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Tian, F.; Cui, S.; Song, J.; Zhao, S.; Brown, E.W.; Meng, J. Differential Gene Expression by RamA in Ciprofloxacin-Resistant Salmonella Typhimurium. PLoS ONE 2011, 6, e22161. [Google Scholar] [CrossRef]
- Hao, Z.; Lou, H.; Zhu, R.; Zhu, J.; Zhang, D.; Zhao, B.S.; Zeng, S.; Chen, X.; Chan, J.; He, C.; et al. The multiple antibiotic resistance regulator MarR is a copper sensor in Escherichia coli. Nat. Chem. Biol. 2014, 10, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Wilks, A. Extracellular heme uptake and the challenges of bacterial cell membranes. Curr. Top. Membr. 2012, 69, 359–392. [Google Scholar] [CrossRef] [Green Version]
- Chubiz, L.M.; Glekas, G.D.; Rao, C.V. Transcriptional cross talk within the mar-sox-rob regulon in Escherichia coli is limited to the rob and marRAB operons. J. Bacteriol. 2012, 194, 4867–4875. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.G.; Bartlett, E.S.; Rosner, J.L.; Wall, M.E. Activation of the Escherichia coli marA/soxS/rob regulon in response to transcriptional activator concentration. J. Mol. Biol. 2008, 380, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Bailey, A.M.; Ivens, A.; Kingsley, R.; Cottell, J.L.; Wain, J.; Piddock, L. RamA, a member of the AraC/XylS family, influences both virulence and efflux in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2010, 192, 1607–1616. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Jamal, S.B.; Hassan, S.S.; Carvalho, P.V.S.D.; Almeida, S.; Barh, D.; Ghosh, P.; Silva, A.; Castro, T.L.P.; Azevedo, V. Two-Component Signal Transduction Systems of Pathogenic Bacteria As Targets for Antimicrobial Therapy: An Overview. Front. Microbiol. 2017, 8, 1878. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Kurushima, J.; Hashimoto, Y.; Tomita, H. Progress Overview of Bacterial Two-Component Regulatory Systems as Potential Targets for Antimicrobial Chemotherapy. Antibiotics 2020, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Murret-Labarthe, C.; Kerhoas, M.; Dufresne, K.; Daigle, F. New Roles for Two-Component System Response Regulators of Salmonella enterica Serovar Typhi during Host Cell Interactions. Microorganisms 2020, 8, 722. [Google Scholar] [CrossRef]
- Tsai, M.H.; Liang, Y.H.; Chen, C.L.; Chiu, C.H. Characterization of Salmonella resistance to bile during biofilm formation. J. Microbiol. Immunol. 2020, 53, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Martynowycz, M.W.; Rice, A.; Andreev, K.; Nobre, T.M.; Kuzmenko, I.; Wereszczynski, J.; Gidalevitz, D. Salmonella membrane structural remodeling increases resistance to antimicrobial peptide LL-37. ACS Infect. Dis. 2019, 5, 1214–1222. [Google Scholar] [CrossRef]
- Kohli, N.; Crisp, Z.; Riordan, R.; Li, M.; Alaniz, R.C.; Jayaraman, A. The microbiota metabolite indole inhibits Salmonella virulence: Involvement of the PhoPQ two-component system. PLoS ONE 2018, 13, e0190613. [Google Scholar] [CrossRef] [Green Version]
- Kapach, G.; Nuri, R.; Schmidt, C.; Danin, A.; Ferrera, S.; Savidor, A.; Gerlach, R.G.; Shai, Y. Loss of the periplasmic chaperone Skp and mutations in the efflux pump AcrAB-TolC play a role in acquired resistance to antimicrobial peptides in Salmonella typhimurium. Front. Microbiol. 2020, 11, 189. [Google Scholar] [CrossRef]
- Zhai, Y.J.; Sun, H.R.; Luo, X.W.; Liu, J.H.; Pan, Y.S.; Wu, H.; Yuan, L.; Liang, J.; He, D.D.; Hu, G.Z. CpxR regulates the colistin susceptibility of Salmonella Typhimurium by a multitarget mechanism. J. Antimicrob. Chemother. 2020, 75, 2780–2786. [Google Scholar] [CrossRef]
- Lin, Y.H.; Yang, T.C.; Hu, W.S. Characterization of stm3030 and stm3031 genes of Salmonella enterica serovar Typhimurium in relation to cephalosporin resistance. J. Microbiol. Immunol. 2019, 52, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Kenney, L.J. The role of acid stress in Salmonella pathogenesis. Curr. Opin. Microbiol. 2019, 47, 45–51. [Google Scholar] [CrossRef]
- Huang, J.; Li, C.; Song, J.; Velkov, T.; Wang, L.; Zhu, Y.; Li, J. Regulating polymyxin resistance in Gram-negative bacteria: Roles of two-component systems PhoPQ and PmrAB. Future Microbiol. 2020. [Google Scholar] [CrossRef]
- Cho, Y.H.; Aziz, M.R.F.; Sutradhar, T.; Bashal, J.; Cojocari, V.; McPhee, J.B. Conserved regulation of omptin proteases by the PhoPQ two-component regulatory system in Enterobacteriaceae. bioRxiv 2020. [Google Scholar] [CrossRef]
- Humphreys, S.; Rowley, G.; Stevenson, A.; Anjum, M.F.; Woodward, M.J.; Gilbert, S.; Kormanec, J.; Roberts, M. Role of the Two-Component Regulator CpxAR in the Virulence of Salmonella enterica Serotype Typhimurium. Infect. Immun. 2004, 72, 4654–4661. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Palmer, A.D.; Vanderpool, C.K.; Slauch, J.M. The sRNA PinT contributes to PhoP-mediated regulation of the SPI1 T3SS in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Palmer, A.D.; Kim, K.; Slauch, J.M. PhoP-mediated repression of the SPI1 T3SS in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2019, 201, e00264-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, V.B.; Vaidyanathan, V.; Mondal, A.; Rajamohan, G. Role of the Two Component Signal Transduction System CpxAR in Conferring Cefepime and Chloramphenicol Resistance in Klebsiella pneumoniae NTUH-K2044. PLoS ONE 2012, 7, e33777. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Pinet, E.; Pagès, J.M. Complex response of the CpxAR two-component system to β-lactams on antibiotic resistance and envelop homeostasis in Enterobacteriaceae. Antimicrob. Agents Chemother. 2020, 64, e00291-20. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.S.; Chen, H.W.; Zhang, R.Y.; Huang, C.Y.; Shen, C.F. The Expression Levels of Outer Membrane Proteins STM1530 and OmpD, Which Are Influenced by the CpxAR and BaeSR Two-Component Systems, Play Important Roles in the Ceftriaxone Resistance of Salmonella enterica Serovar Typhimurium. Antimicrob. Agents Chemother. 2011, 55, 3829–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.S.; Lin, J.F.; Lin, Y.H.; Chang, H.Y. Outer Membrane Protein STM3031 (Ail/OmpX-Like Protein) Plays a Key Role in the Ceftriaxone Resistance of Salmonella enterica Serovar Typhimurium. Antimicrob. Agents Chemother. 2009, 53, 3248–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Nishino, K.; Nikaido, E.; Yamaguchi, A. Regulation of Multidrug Efflux Systems Involved in Multidrug and Metal Resistance of Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2007, 189, 9066–9075. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, P.; Collao, B.; Morales, E.H.; Calderón, I.L.; Ipinza, F.; Parra, S.; Saavedra, C.P.; Gil, F. Characterization of the BaeSR two-component system from Salmonella Typhimurium and its role in ciprofloxacin-induced mdtA expression. Arch. Microbiol. 2012, 194, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Kenney, L.J.; Anand, G.S. EnvZ/OmpR Two-Component Signaling: An Archetype System That Can Function Noncanonically. EcoSal Plus 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Mizusaki, H.; Kenney, L.J. A FRET-Based DNA Biosensor Tracks OmpR-Dependent Acidification of Salmonella during Macrophage Infection. PLoS Biol. 2015, 13, e1002116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Morales, F. Impact of Salmonella enterica Type III Secretion System Effectors on the Eukaryotic Host Cell. ISRN Cell Biol. 2012, 2012, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Gunn, J.S. The Salmonella PmrAB regulon: Lipopolysaccharide modifications, antimicrobial peptide resistance and more. Trends Microbiol. 2008, 16, 284–290. [Google Scholar] [CrossRef]
- Hjort, K.; Nicoloff, H.; Andersson, D.I. Unstable tandem gene amplification generates heteroresistance (variation in resistance within a population) to colistin in Salmonella enterica. Mol. Microbiol. 2016, 102, 274–289. [Google Scholar] [CrossRef]
- Quesada, A.; Porrero, M.C.; Téllez, S.; Palomo, G.; García, M.; Domínguez, L. Polymorphism of genes encoding PmrAB in colistin-resistant strains of Escherichia coli and Salmonella enterica isolated from poultry and swine. J. Antimicrob. Chemother. 2014, 70, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.M.; Strandberg, K.L.; Conroy, M.; Gunn, J.S. Cationic antimicrobial peptides serve as activation signals for the Salmonella Typhimurium PhoPQ and PmrAB regulons in vitro and in vivo. Front. Cell Infect. Microbiol. 2012, 2, 102. [Google Scholar] [CrossRef] [Green Version]
- Molina-Henares, A.J.; Godoy, P.; Duque, E.; Ramos, J.L. A general profile for the MerR family of transcriptional regulators constructed using the semi-automated Provalidator tool. Environ. Microbiol. Rep. 2009, 1, 518–523. [Google Scholar] [CrossRef]
- Brown, N.L.; Stoyanov, J.V.; Kidd, S.P.; Hobman, J.L. The MerR family of transcriptional regulators. FEMS Microbiol. Rev. 2003, 27, 145–163. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez, M.M.; Cerminati, S.; Checa, S.K.; Soncini, F.C. Dissecting the metal selectivity of MerR monovalent metal ion sensors in Salmonella. J. Bacteriol. 2013, 195, 3084–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, N.; Oi, Y.; Takeda-Shitaka, M.; Kanou, K.; Umeyama, H.; Haneda, T.; Miki, T.; Hosoya, S.; Danbara, H. Identification of amino acid residues of Salmonella SlyA that are critical for transcriptional regulation. Microbiology 2007, 153, 548–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, I.C.; Grove, A. Molecular Mechanisms of Ligand-Mediated Attenuation of DNA Binding by MarR Family Transcriptional Regulators. J. Mol. Cell Biol. 2010, 2, 243–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deochand, D.K.; Grove, A. MarR family transcription factors: Dynamic variations on a common scaffold. Crit. Rev. Biochem. Mol. 2017, 52, 595–613. [Google Scholar] [CrossRef]
- Cabezas, C.E.; Briones, A.C.; Aguirre, C.; Pardo-Esté, C.; Castro-Severyn, J.; Salinas, C.R.; Baquedano, M.S.; Hidalgo, A.A.; Fuentes, J.A.; Morales, E.H.; et al. The transcription factor SlyA from Salmonella Typhimurium regulates genes in response to hydrogen peroxide and sodium hypochlorite. Res. Microbiol. 2018, 169, 263–278. [Google Scholar] [CrossRef]
- Tian, S.; Wang, C.; Li, Y.; Bao, X.; Zhang, Y.; Tang, T. The Impact of SlyA on Cell Metabolism of Salmonella typhimurium: A Joint Study of Transcriptomics and Metabolomics. J. Proteome Res. 2020, 20, 184–190. [Google Scholar] [CrossRef]
- Ballesté-Delpierre, C.; Fàbrega, A.; Ferrer-Navarro, M.; Mathur, R.; Ghosh, S.; Vila, J. Attenuation of in vitro host–pathogen interactions in quinolone-resistant Salmonella Typhi mutants. J. Antimicrob. Chemother. 2016, 71, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.; Park, S.; Kwak, M.K.; Lee, J.S. Regulation of gene expression by protein lysine acetylation in Salmonella. J. Microbiol. 2020, 58, 979–987. [Google Scholar] [CrossRef]
- Ali, S.S.; Whitney, J.C.; Stevenson, J.; Robinson, H.; Howell, P.L.; Navarre, W.W. Structural Insights into the Regulation of Foreign Genes in Salmonella by the Hha/H-NS Complex. J. Biol. Chem. 2013, 288, 13356–13369. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, V.H.; Martinez, L.C.; Santana, F.J.; Knodler, L.A.; Steele-Mortimer, O.; Puente, J.L. HilD-mediated transcriptional cross-talk between SPI-1 and SPI-2. Proc. Natl. Acad. Sci. USA 2008, 105, 14591–14596. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Escobar, G.A.; Grépinet, O.; Raymond, P.; Abed, N.; Velge, P.; Virlogeux-Payant, I. H-NS is the major repressor of Salmonella Typhimurium Pef fimbriae expression. Virulence 2019, 10, 849–867. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Groisman, E.A. Salmonella express foreign genes during infection by degrading their silencer. Proc. Natl. Acad. Sci. USA 2020, 117, 8074–8082. [Google Scholar] [CrossRef] [PubMed]
- Bouffartigues, E.; Buckle, M.; Badaut, C.; Travers, A.; Rimsky, S. H-NS cooperative binding to high-affinity sites in a regulatory element results in transcriptional silencing. Nat. Struct. Mol. Biol. 2007, 14, 441–448. [Google Scholar] [CrossRef]
- Lahiri, A.; Das, P.; Chakravortty, D. The LysR-type transcriptional regulator Hrg counteracts phagocyte oxidative burst and imparts survival advantage to Salmonella enterica serovar Typhimurium. Microbiology 2008, 154, 2837–2846. [Google Scholar] [CrossRef] [Green Version]
- Oliver, P.; Peralta-Gil, M.; Tabche, M.L.; Merino, E. Molecular and structural considerations of TF-DNA binding for the generation of biologically meaningful and accurate phylogenetic footprinting analysis: The LysR-type transcriptional regulator family as a study model. BMC Genom. 2016, 17, 686. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Fillat, M.F.; Lanas, Á. Transcriptional regulators: Valuable targets for novel antibacterial strategies. Future Med. Chem. 2018, 10, 541–560. [Google Scholar] [CrossRef] [PubMed]
- Koentjoro, M.P.; Ogawa, N. Structural studies of transcriptional regulation by LysR-type transcriptional regulators in bacteria. Rev. Agric. Sci. 2018, 6, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Passaris, I.; Cambré, A.; Govers, S.K.; Aertsen, A. Bimodal Expression of the Salmonella Typhimurium spv Operon. Genetics 2018, 210, 621–635. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yuan, Y.; Wang, X.; Chen, J.; Wu, Y.; Wang, X.; Xu, L.; Yin, K.; Liu, Z.; Yin, C.; et al. Comparative study of Salmonella enterica serovar Enteritidis genes expressed within avian and murine macrophages via selective capture of transcribed sequences (SCOTS). Appl. Microbiol. Biotechnol. 2018, 102, 6567–6579. [Google Scholar] [CrossRef]
- Dorman, C.J.; Dorman, M.J. Control of virulence gene transcription by indirect readout in Vibrio cholerae and Salmonella enterica serovar Typhimurium. Environ. Microbiol. 2017, 19, 3834–3845. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Lv, R.; Jiang, X.; Cao, H.; Du, Y.; Jiang, L.; Liu, B. Global regulatory function of the low oxygen-induced transcriptional regulator LoiA in Salmonella Typhimurium revealed by RNA sequencing. Biochem. Biophys. Res. Commun. 2018, 503, 2022–2027. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Singh, A.K.; Kumaran, S. Structural and biochemical characterization of ligand recognition by CysB, the master regulator of sulfate metabolism. Biochimie 2017, 142, 112–124. [Google Scholar] [CrossRef]
- Turner, K.A.; Eckert, S.E.; Turner, D.J.; Yasir, M.; Webber, M.A.; Charles, I.G.; Parkhill, J.; Wain, J. A whole-genome screen identifies Salmonella enterica serovar Typhi genes involved in fluoroquinolone susceptibility. J. Antimicrob. Chemother. 2020, 75, 2516–2525. [Google Scholar] [CrossRef] [PubMed]
- Rebollar-Flores, J.E.; Medina-Aparicio, L.; Osio-Becerro, V.E.; Villarreal, J.M.; Mayo, S.; Mendoza, B.D.; Rodríguez-Gutierrez, S.; Olvera, L.; Dávila, S.; Encarnación, S.; et al. The Salmonella enterica Serovar Typhi ltrR Gene Encodes Two Proteins Whose Transcriptional Expression Is Upregulated by Alkaline pH and Repressed at Their Promoters and Coding Regions by H-NS and Lrp. J. Bacteriol. 2020, 202, e00783-19. [Google Scholar] [CrossRef] [PubMed]
- Mandal, R.S.; Ta, A.; Sinha, R.; Theeya, N.; Ghosh, A.; Tasneem, M.; Bhunia, A.; Koley, H.; Das, S. Ribavirin suppresses bacterial virulence by targeting LysR-type transcriptional regulators. Sci. Rep. 2016, 6, 39454. [Google Scholar] [CrossRef]
- Guiney, D.G.; Fierer, J. The role of the spv genes in Salmonella pathogenesis. Front. Microbiol. 2011, 2, 129. [Google Scholar] [CrossRef] [Green Version]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Li, X.; Lv, R.; Feng, L. LoiA directly represses lon gene expression to activate the expression of Salmonella pathogenicity island-1 genes. Res. Microbiol. 2019, 170, 131–137. [Google Scholar] [CrossRef]
- VanDrisse, C.M.; Escalante-Semerena, J.C. In Salmonella enterica, OatA (Formerly YjgM) Uses O-Acetyl-Serine and Acetyl-CoA to Synthesize N,O-Diacetylserine, Which Upregulates Cysteine Biosynthesis. Front. Microbiol. 2018, 9, 2838. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Pathogenicity Island | Virulence Factor | Function | Reference |
|---|---|---|---|
| SPI-1 | SipA, SipB | invasion of intestinal epithelial cells; development of SCV; encoding the effector proteins important for actin cytoskeleton rearrangements; induction of IL-8 and Salmonella-elicited epithelial chemoattractant secretion; membrane ruffling that | [12,33,37,38,39,40,41,42] |
| SPI-2 | SseF, SseG | mediates processes required for bacterial replication within host macrophages; SCV localization; inhibition of fusion between lysosomes and SCVs; avoidance of NADPH oxidase-dependent killing by macrophages; encoding of effector, chaperon and translocon proteins | [12,33,37,38,39,40,43] |
| SPI-3 | MisL | host cell attachment and long-term colonization; intramacrophage survival | [37,38] |
| SPI-4 | SiiE | mediation in adhesion to epithelial cells; involvement in intracellular survival and systemic infection by S. Typhimurium | [12,37,38] |
| SPI-5 | SopB | important for S. Dublin virulence; involvement in intracellular survival and systemic infection by S. Typhimurium; membrane ruffling; activation of pro-inflammatory response | [12,37,38,44] |
| SPI-6 | Tar4, SciG, SciS | combating host resistance to pathogen colonization; involvement in intracellular survival and systemic infection by S. Typhimurium | [37,45] |
| SPI-7 | - | virulence factor for typhoid fever | [46] |
| SPI-11 | PagC, PagD, EnvE, EnvF | involvement in intracellular survival and systemic infection by S. Typhimurium; resistance to antimicrobial peptides; survival within macrophages | [37,38] |
| SPI-12 | SspH2 | intracellular survival | [38] |
| SPI-13 | STM3118, STM3119 | survival within macrophages | [38] |
| SPI-14 | LoiA | encoding the LoiA protein necessary for the invasion of the intestinal epithelium by S. Typhimurium; regulation of SPI-1 genes | [37,38,40] |
| SPI-16 | STM0557 | long-term colonization | [38] |
| SPI-19 | - | necessary for survival within macrophages and initial colonization by S. Pullorum in chicken | [47] |
| TCS System | System Activation Signals | Components of System | Antibiotic Resistance | Function | Reference | |
|---|---|---|---|---|---|---|
| HK | RR | |||||
| PhoPQ | low Mg2+; acidic pH; cationic antimicrobial peptide (CAMP); hyperosmotic stress | PhoQ | PhoP | polymyxin | LPS modifications (by adding 4-aminoarabinose to lipid A); downregulation of the expression of genes located on SPI-1; increasing resistance to both conventional antibiotics and antimicrobial peptides | [116,117,118] |
| CpxAR | acidic pH | CpxA | CpxR | β-lactam | resistance to β-lactam antibiotics in the absence of AcrB efflux pump; downregulation of the MDR-related genes expression; decreasing porin expression | [119] |
| BaeSR | spheroplasting and exposure to indole, tannin, zinc, or cooper | BaeS | BaeR | ceftriaxone | upregulation of MDR efflux pumps; drug resistance by regulating the gene expression encoding drug transporters | [120] |
| EnvZ/OmpR | acidic pH | EnvZ | OmpR | β-lactam | activation of the ssrA transcription and production of SsrA and SsrB; decreasing porin expression, upregulation of MDR efflux pumps | [8,121] |
| PmrAB | acidic pH; high Fe3+/Al3+; activation by the PhoPQ system | PmrA | PmrB | polymyxin | LPS modifications (modification of lipid A) | [122] |
| LTTR System | Function | Reference |
|---|---|---|
| SpvR | enhancing bacterial virulence by inhibiting autophagy; inducing the spv operon expression in the intracellular environment of host cells | [160,161] |
| LeuO | forming tetramer residues involved in oligomerization, DNA binding, transcriptional regulation | [158,162] |
| LoiA | encoded by SPI-14; essential for the Salmonella invasion of intestinal epithetical cells | [40,163] |
| STM0030 | regulating genes essential for Salmonella intracellular replication and virulence | [40] |
| CysB | activator of the Cys regulon under the sulfur limitation | [164,165] |
| LtrR | indirect regulation of the porin synthesis; bacterial transformation and bile resistance | [166] |
| Hrg | protection against oxidative stress; bacterial growth and survival within macrophages | [167] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wójcicki, M.; Świder, O.; Daniluk, K.J.; Średnicka, P.; Akimowicz, M.; Roszko, M.Ł.; Sokołowska, B.; Juszczuk-Kubiak, E. Transcriptional Regulation of the Multiple Resistance Mechanisms in Salmonella—A Review. Pathogens 2021, 10, 801. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10070801
Wójcicki M, Świder O, Daniluk KJ, Średnicka P, Akimowicz M, Roszko MŁ, Sokołowska B, Juszczuk-Kubiak E. Transcriptional Regulation of the Multiple Resistance Mechanisms in Salmonella—A Review. Pathogens. 2021; 10(7):801. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10070801
Chicago/Turabian StyleWójcicki, Michał, Olga Świder, Kamila J. Daniluk, Paulina Średnicka, Monika Akimowicz, Marek Ł. Roszko, Barbara Sokołowska, and Edyta Juszczuk-Kubiak. 2021. "Transcriptional Regulation of the Multiple Resistance Mechanisms in Salmonella—A Review" Pathogens 10, no. 7: 801. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10070801