
The Novel Zoonotic Pathogen, Anaplasma capra, Infects Human Erythrocytes, HL-60, and TF-1 Cells In Vitro

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathogen Collection and Identification
2.2. Human Erythrocytes Isolation and Preserved In Vitro
2.3. Thawing and Cultivation of HL-60 and TF-1 Cells
2.4. Pathogen Inoculation
2.5. Cell Infection Identification
2.5.1. Wright–Giemsa Staining
2.5.2. Immunocytochemistry
2.5.3. Chromogenic In Situ Hybridization (CISH)
2.5.4. Transmission Electron Microscopy (TEM)
3. Results
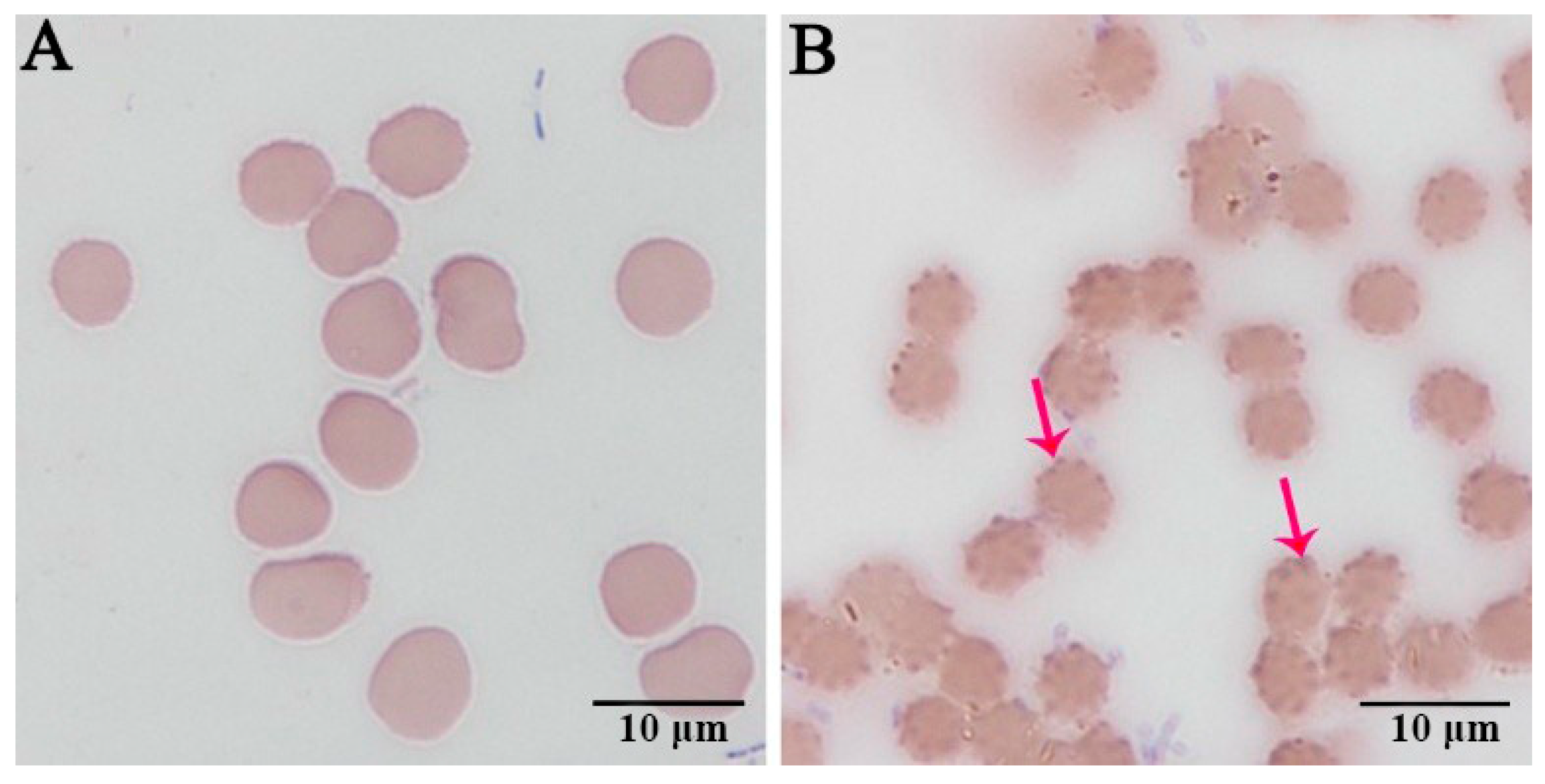
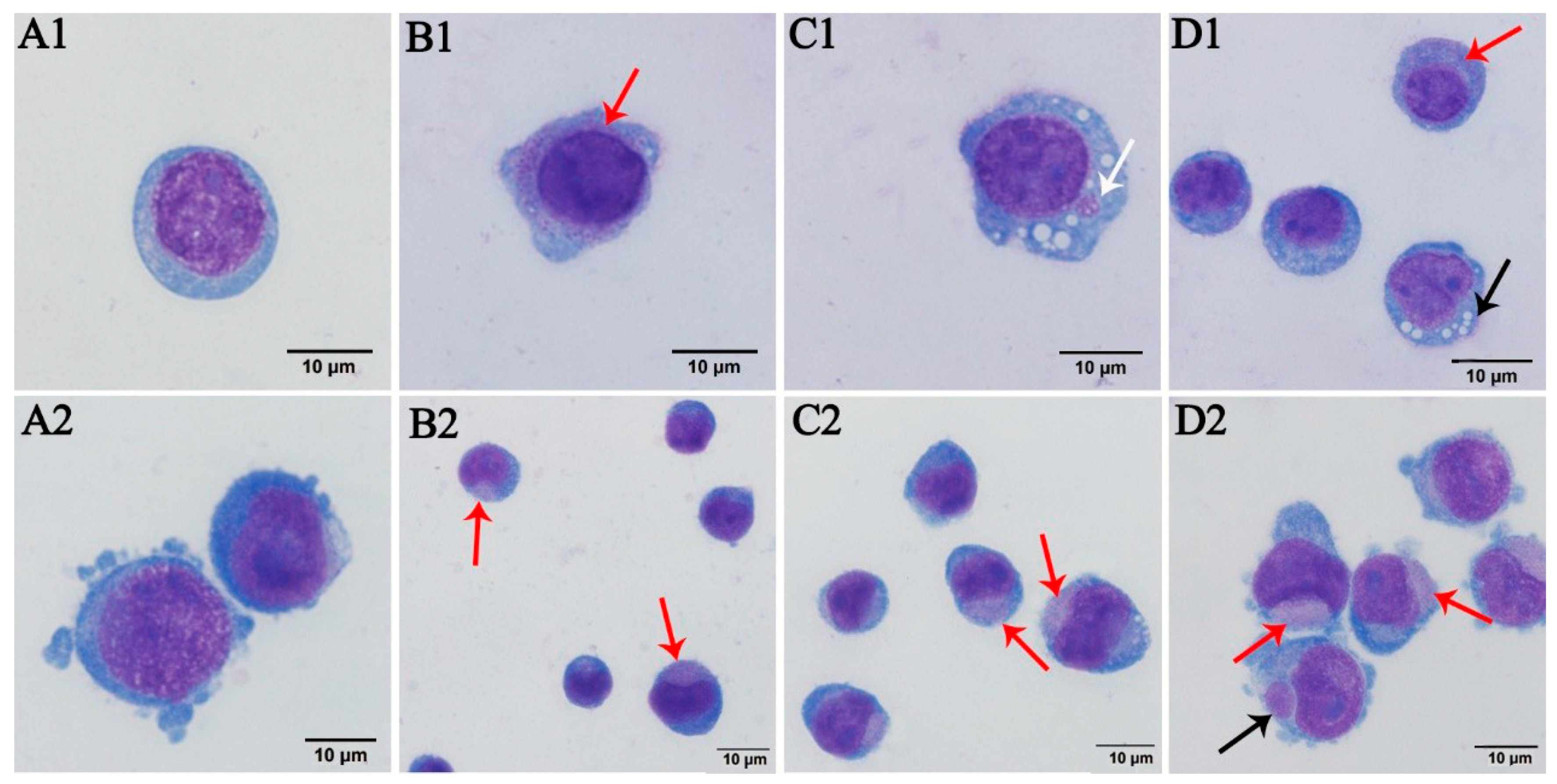
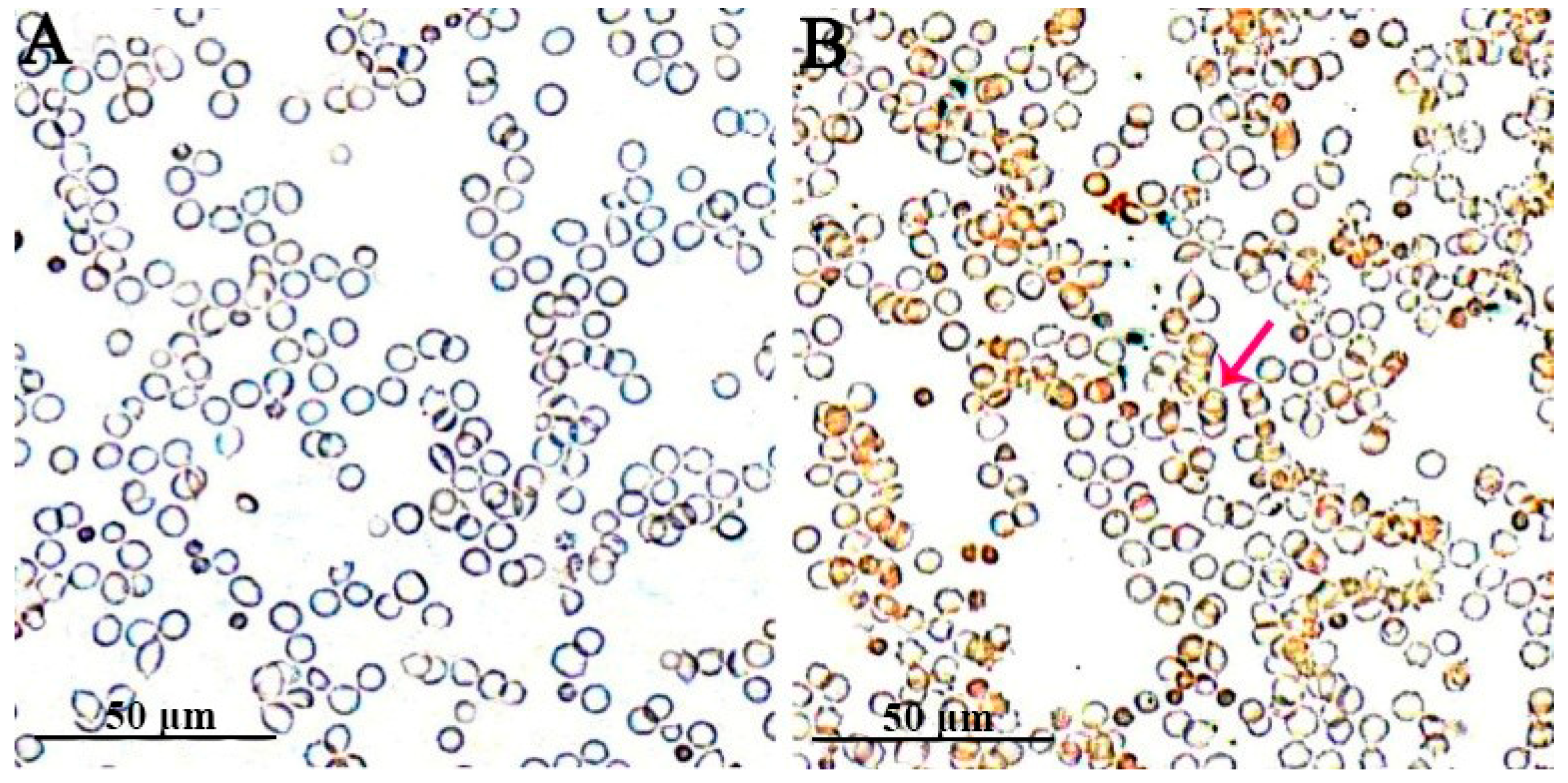
3.1. Wright–Giemsa Staining Results
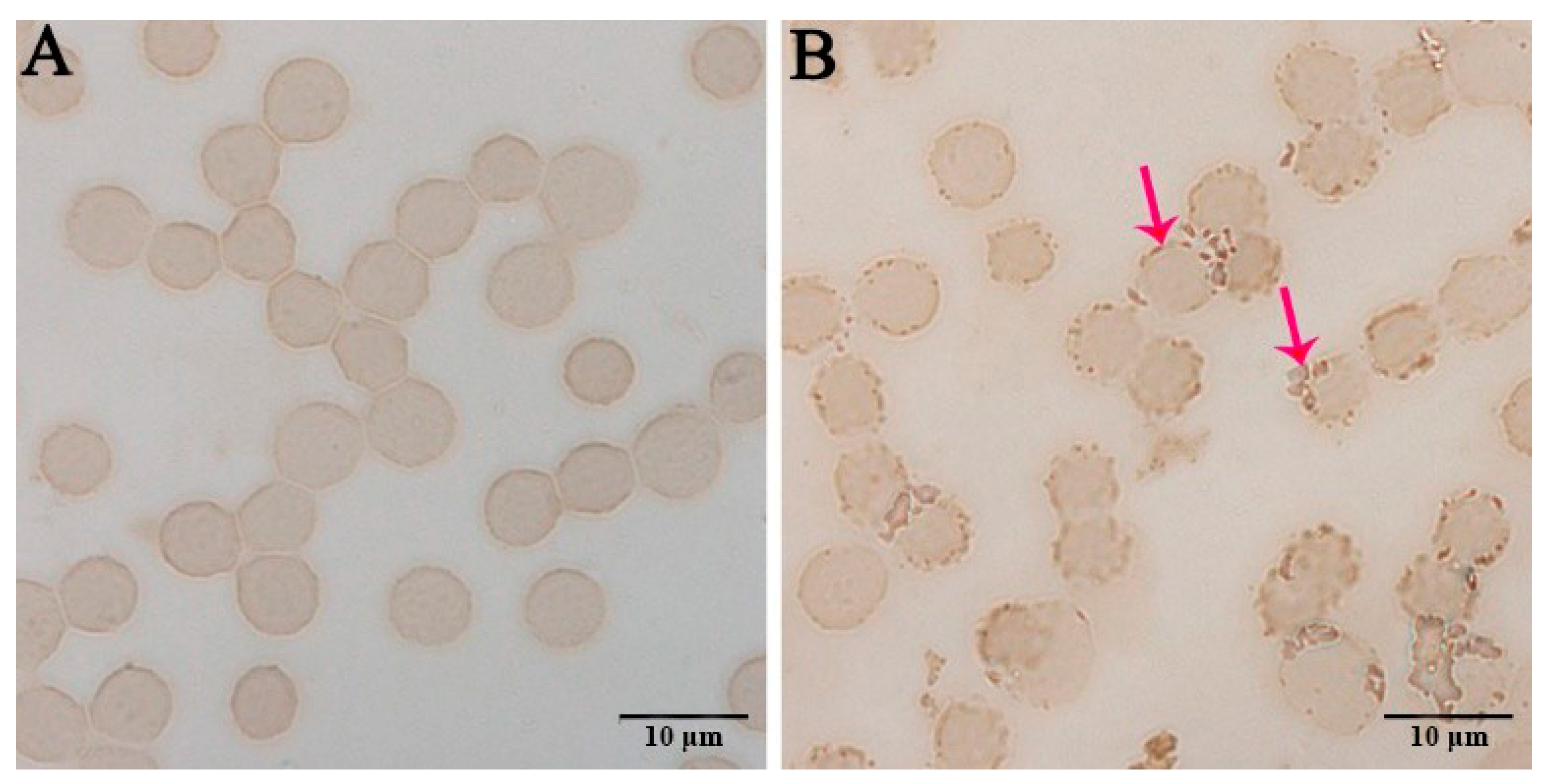
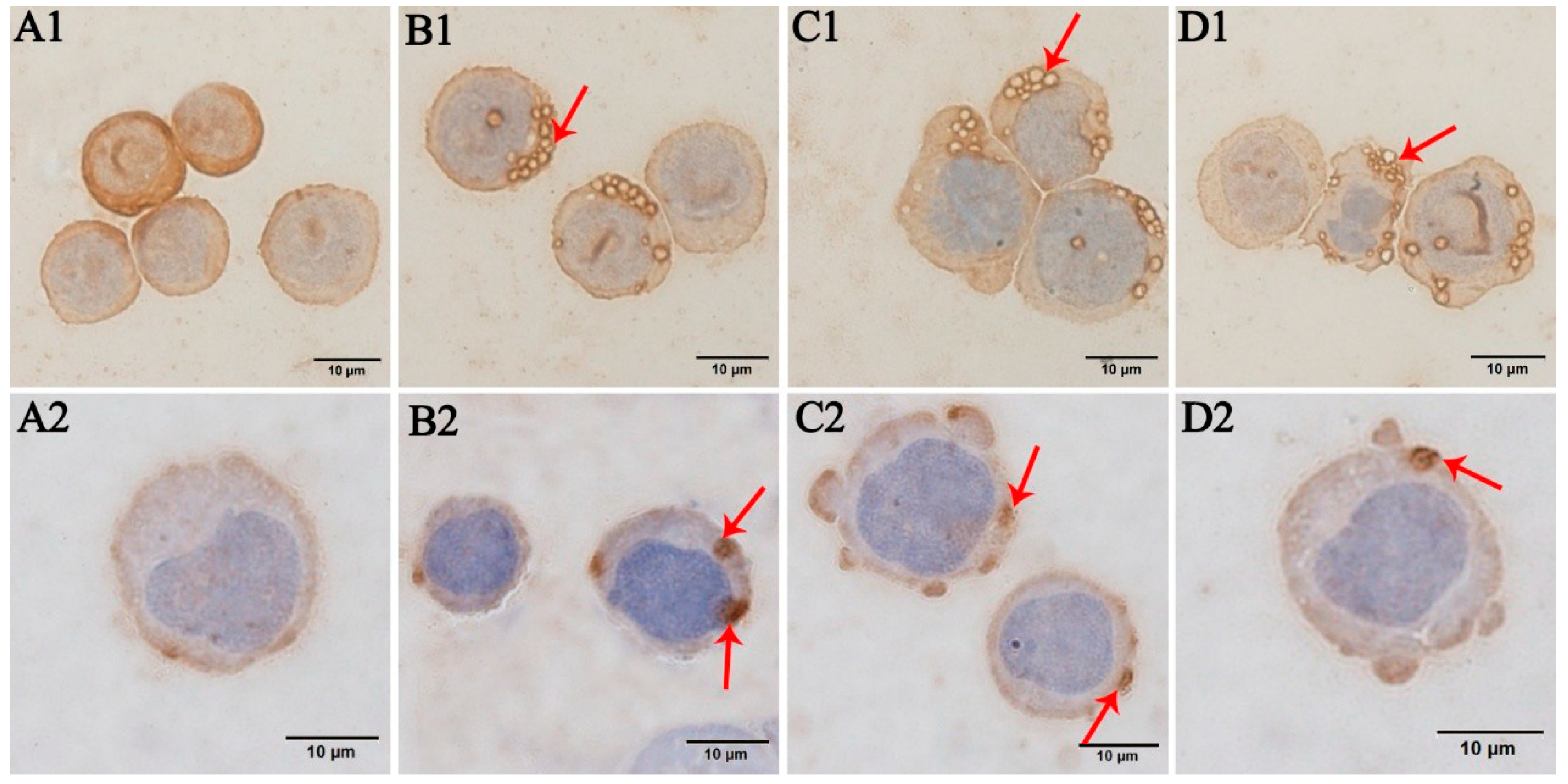
3.2. Immunocytochemistry Results
3.3. CISH Results
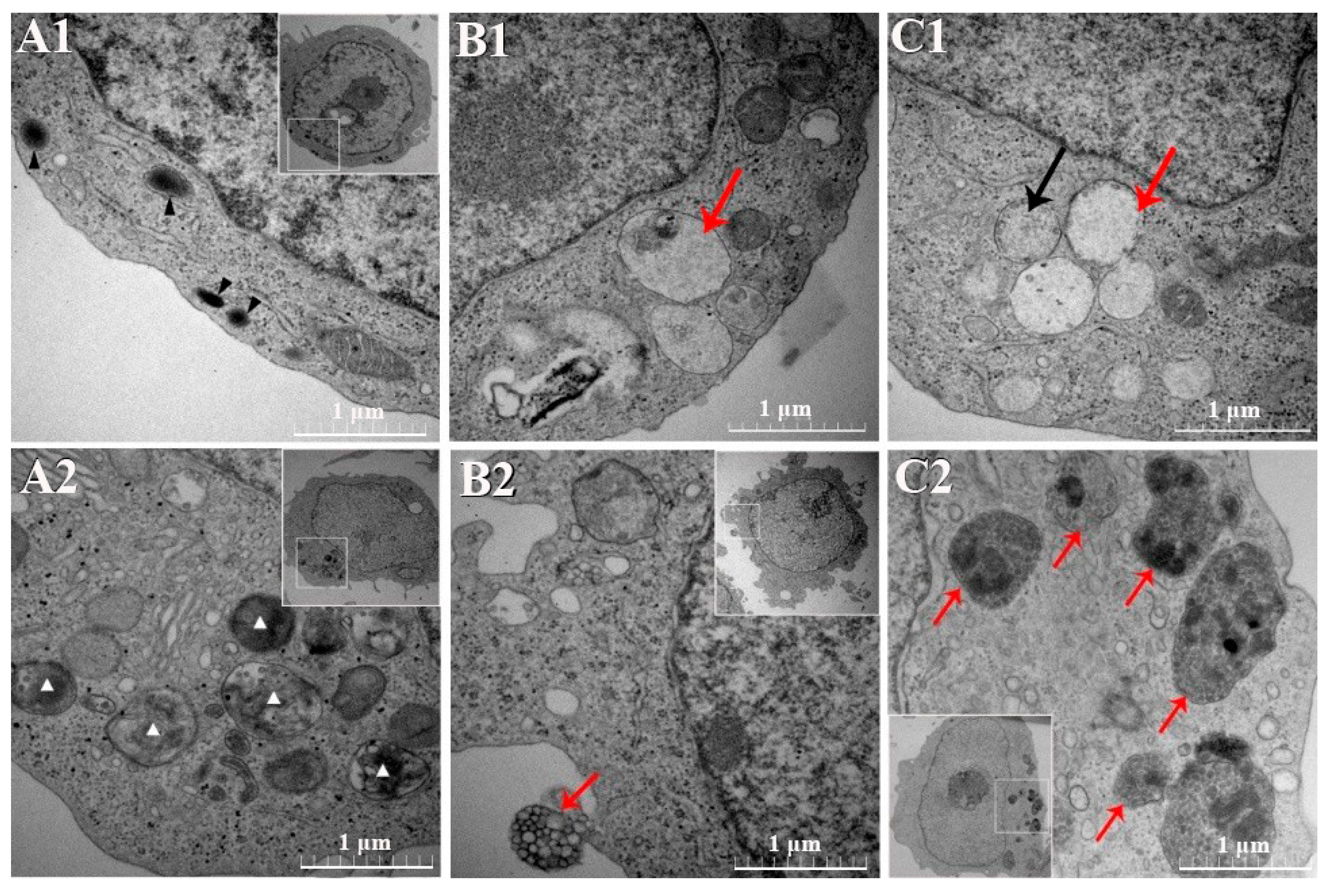
3.4. TEM Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dehhaghi, M.; Panahi, H.K.S.; Holmes, E.C.; Hudson, B.J.; Guillemin Gilles, J. Human Tick-borne diseases in Australia. Front. Cell. Infect. Microbiol. 2019, 9, 3. [Google Scholar] [CrossRef]
- Zhou, Z.; Nie, K.; Tang, C.; Wang, Z.; Zhou, R.; Hu, S.; Zhang, Z. Phylogenetic analysis of the genus Anaplasma in southwestern China based on 16S rRNA sequence. Res. Vet. Sci. 2010, 89, 262–265. [Google Scholar] [CrossRef]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G. Human infection with a novel tick-borne Anaplasma species in China, a surveillance study. Lancet Infect. Dis. 2015, 15, 663–670. [Google Scholar] [CrossRef]
- Wei, R.; Liu, H.B.; Jongejan, F.; Jiang, B.G.; Chang, Q.C.; Fu, X.; Jiang, J.F.; Jia, N.; Cao, W.C. Cultivation of Anaplasma ovis in the HL-60 human promyelocytic leukemia cell line. Emerg. Microbes Infect. 2017, 6, e83. [Google Scholar]
- Lee, S.; Shin, N.; Kim, C.; Park, S.; Yun, N.R.; Kim, D.M.; Jung, D.S. First identification of Anaplasma phagocytophilum in both a biting tick Ixodes nipponensis and a patient in Korea: A case report. BMC Infect. Dis. 2020, 20, 826. [Google Scholar] [CrossRef]
- Shi, Y.; Yang, J.; Guan, G.; Liu, Z.; Song, M. Molecular investigation of Anaplasma species in sheep from Heilongjiang Province, northeast China identified four Anaplasma species and a novel genotype of Anaplasma capra. Parasitol. Int. 2020, 76, 102072. [Google Scholar] [CrossRef]
- Guo, W.P.; Zhang, B.; Wang, Y.H.; Xu, G.; Zhou, E.M. Molecular identification and characterization of Anaplasma capra and Anaplasma platys-like in Rhipicephalus microplus in Ankang, northwest China. BMC Infect. Dis. 2019, 19, 434. [Google Scholar] [CrossRef]
- Qin, X.R.; Han, F.J.; Luo, L.M.; Zhao, F.M.; Han, H.J.; Zhang, Z.T.; Liu, J.W.; Xue, Z.F.; Liu, M.M.; Ma, D.Q.; et al. Anaplasma species detected in Haemaphysalis longicornis tick from China. Ticks Tick Borne Dis. 2018, 9, 840–843. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Z.; Niu, Q.; Liu, J.; Han, R.; Liu, G.; Shi, Y.; Luo, J.; Yin, H. Molecular survey and characterization of a novel Anaplasma species closely related to Anaplasma capra in ticks, northwestern China. Parasite Vector 2016, 9, 603. [Google Scholar] [CrossRef] [Green Version]
- Han, R.; Yang, J.F.; Mukhtar, M.U.; Chen, Z.; Niu, Q.L.; Lin, Y.Q.; Liu, G.Y.; Luo, J.X.; Yin, H.; Liu, Z.J. Molecular detection of Anaplasma infections in Ixodid ticks from the Qinghai-Tibet Plateau. Infect. Dis. Poverty 2019, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Genotypic analysis of Piroplasms and associated pathogens from ticks infesting cattle in Korea. Microorganisms 2020, 8, 728. [Google Scholar] [CrossRef]
- Seo, M.G.; Ouh, I.O.; Kwon, O.D.; Kwak, D. Molecular detection of Anaplasma phagocytophilum-like Anaplasma spp. and pathogenic A. phagocytophilum in cattle from south Korea. Mol. Phylogenet. Evol. 2018, 126, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Borjesson, D.L. Culture, isolation, and labeling of Anaplasma phagocytophilum for subsequent infection of human neutrophils. Methods Mol. Biol. 2008, 431, 159–171. [Google Scholar]
- Truchan, H.K.; Cockburn, C.L.; May, L.J.; Lauren, V.B.; Jason, C. Anaplasma phagocytophilum-occupied vacuole interactions with the host cell cytoskeleton. Vet. Sci. 2016, 3, 25. [Google Scholar] [CrossRef]
- Truchan, H.K.; Cockburn, C.L.; Hebert, K.S.; Magunda, F.; Noh, S.M.; Carlyon, J.A. The pathogen-occupied vacuoles of Anaplasma phagocytophilum and Anaplasma marginale interact with the endoplasmic reticulum. Front. Cell. Infect. Microbiol. 2016, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Massung, R.F.; Levin, M.L.; Munderloh, U.G.; Silverman, D.J.; Lynch, M.J.; Gaywee, J.K.; Kurtti, T.J. Isolation and propagation of the Ap-Variant 1 strain of Anaplasma phagocytophilum in a tick cell line. J. Clin. Microbiol. 2007, 45, 2138–2143. [Google Scholar] [CrossRef] [Green Version]
- Bell-Sakyi, L.; Zweygarth, E.; Blouin, E.F.; Gould, E.A.; Jongejan, F. Tick cell lines: Tools for tick and tick-borne disease research. Trends Parasitol. 2007, 23, 450–457. [Google Scholar] [CrossRef]
- Bastos, C.V.; Passos, L.M.; Vasconcelos, M.M.; Ribeiro, M.F. In vitro establishment and propagation of a Brazilian strain of Anaplasma marginale with appendage in IDE8 (Ixodes scapularis) cells. Braz. J. Microbiol. 2009, 40, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, T.; Tange, T.; Terasawa, T.; Chiba, S.; Kuwaki, T.; Miyagawa, K.; Piao, Y.F.; Miyazono, K.; Urabe, A.; Takaku, F. Establishment and characterization of a unique human cell line that proliferates dependently on GM-CSF, IL-3, or erythropoietin. J. Cell. Physiol. 1989, 140, 323–334. [Google Scholar] [CrossRef]
- Peng, Y.; Lu, C.; Yan, Y.; Shi, K.; Chen, Q.; Zhao, C.; Wang, R.; Zhang, L.; Jian, F.; Ning, C. The first detection of Anaplasma capra, an emerging zoonotic sp., in erythrocytes. Emerg. Microbes Infect. 2021, 10, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.P.; Huang, B.; Zhao, Q.; Xu, G.; Zhou, E.M. Human-pathogenic Anaplasma spp., and Rickettsia spp. in animals in Xi’an, China. PLoS Negl. Trop. Dis. 2018, 12, e0006916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, R.; Westblade, L.F.; Kessler, D.A.; Sfeir, M.; Slavinski, S.; Backenson, B.; Gebhardt, L.; Kane, K.; Laurence, J.; Scherr, D.; et al. Death from Transfusion-Transmitted Anaplasmosis, New York, USA, 2017. Emerg. Microbes Infect. 2018, 24, 1548–1550. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, S.; Lee, Y.S.; Lee, H.K.; Hwang, S.D. Diagnosis and molecular characteristics of human infections caused by Anaplasma phagocytophilum in south Korea. J. Microbiol. 2018, 56, 847–853. [Google Scholar] [CrossRef]
- Yoshikawa, Y.; Sugimoto, K.; Ochiai, Y.; Ohashi, N. Intracellular proliferation of Anaplasma phagocytophilum is promoted via modulation of endoplasmic reticulum stress signaling in host cells. Microbiol. Immunol. 2020, 64, 270–279. [Google Scholar] [CrossRef]
- Shimada, M.; Takamoto, N.; Su, H.; Sasahara, H.; Shimamura, Y.; Ando, S.; Ohashi, N. Predominant shift of different P44-expressing Anaplasma phagocytophilum in infected HL-60, THP-1, NB4, and RF/6A cell lines. Jpn. J. Infect. Dis. 2019, 72, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Baeta, B.A.; Ribeiro, C.C.; Teixeira, R.C.; Cabezas-Cruz, A.; Lygia Passos, M.F.; Zweygarth, E.; Fonseca, A.H. Characterization of two strains of Anaplasma marginale isolated from cattle in Rio de Janeiro, Brazil, after propagation in tick cell culture. Ticks Tick Borne Dis. 2015, 6, 141–145. [Google Scholar] [CrossRef]
- Lis, K.; Najm, N.; de la Fuente, J.; Isabel, F.D.M.; Zweygarth, E.; Pfister, K.; Passos, L.M.F. Use of percoll gradients to purify Anaplasma marginale (Rickettsiales: Anaplasmataceae) from tick cell cultures. Ticks Tick Borne Dis. 2014, 5, 511–515. [Google Scholar] [CrossRef]
- Passos, L.M. In vitro cultivation of Anaplasma marginale and A. phagocytophilum in tick cell lines: A review. Rev. Bras. Parasitol. Vet. 2012, 21, 81–86. [Google Scholar] [CrossRef]
- Lasmar, P.V.; Carvalho, A.U.; Facury Filho, E.J.; Bastos, C.V.; Ribeiro, M.F.B. Evaluating the effectiveness of an inactivated vaccine from Anaplasma marginale derived from tick cell culture. Rev. Bras. Parasitol. Vet. 2012, 21, 112–117. [Google Scholar] [CrossRef]
- Samish, M.; Pipano, E.; Hana, B. Cultivation of Anaplasma marginale from cattle in a Dermacentor cell line. Am. J. Vet. Res. 1988, 49, 254–256. [Google Scholar]
- Battilani, M.; De Arcangeli, S.; Balboni, A.; Dondi, F. Genetic diversity and molecular epidemiology of Anaplasma. Infect. Genet. Evol. 2017, 49, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, M.; Wang, Z.; Wang, J.; Peng, Y.; Li, Y.; Guan, G.; Luo, J.; Yin, H. Molecular survey and genetic identification of Anaplasma species in goats from central and southern China. Appl. Environ. Microbiol. 2012, 78, 464–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, M.B.; Hayes, S.F.; Goodman, J.L. Monocytic differentiation inhibits infection and granulocytic differentiation potentiates infection by the agent of Human granulocytic ehrlichiosis. Infect. Immun. 1998, 66, 3410–3415. [Google Scholar] [CrossRef] [Green Version]
- Trong, T.P.N.; Wang, J.; Majzoub-Altweck, M.; Fell, S.; Straubinger, R.K. Recovering host cell-free Anaplasma phagocytophilum from HL-60 cells by using rock tumbler grit in comparison to the syringe lysis method. Ticks Tick Borne Dis. 2019, 10, 280–285. [Google Scholar] [CrossRef]
- Magunda, F.; Thompson, C.W.; Schneider, D.A.; Noh, S.M. Anaplasma marginale actively modulates vacuolar maturation during intracellular infection of its tick vector, Dermacentor andersoni. Appl. Environ. Microbiol. 2016, 82, 4715–4731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattila, J.T.; Munderloh, U.G.; Kurtti, T.J. Phagocytosis of the Lyme disease spirochete, Borrelia burgdorferi, by cells from the ticks, Ixodes scapularis and Dermacentor andersoni, infected with an endosymbiont, Rickettsia peacockii. J. Insect. Sci. 2007, 7, 58. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, P.; Ayllón, N.; Cabezas-Cruz, A.; Bell-Sakyi, L.; Zweygarth, E.; Stuen, S.; de la Fuente, J. Infection of Ixodes spp. tick cells with different Anaplasma phagocytophilum isolates induces the inhibition of apoptotic cell death. Ticks Tick Borne Dis. 2015, 6, 758–767. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Y.; Lu, C.; Yan, Y.; Song, J.; Pei, Z.; Gong, P.; Wang, R.; Zhang, L.; Jian, F.; Ning, C. The Novel Zoonotic Pathogen, Anaplasma capra, Infects Human Erythrocytes, HL-60, and TF-1 Cells In Vitro. Pathogens 2021, 10, 600. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050600
Peng Y, Lu C, Yan Y, Song J, Pei Z, Gong P, Wang R, Zhang L, Jian F, Ning C. The Novel Zoonotic Pathogen, Anaplasma capra, Infects Human Erythrocytes, HL-60, and TF-1 Cells In Vitro. Pathogens. 2021; 10(5):600. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050600
Chicago/Turabian StylePeng, Yongshuai, Chenyang Lu, Yaqun Yan, Jinxing Song, Zhiyang Pei, Pihong Gong, Rongjun Wang, Longxian Zhang, Fuchun Jian, and Changshen Ning. 2021. "The Novel Zoonotic Pathogen, Anaplasma capra, Infects Human Erythrocytes, HL-60, and TF-1 Cells In Vitro" Pathogens 10, no. 5: 600. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10050600