
Canine Morbillivirus from Colombian Lineage Exhibits In Silico and In Vitro Potential to Infect Human Cells

 , and
, and 
Abstract
:1. Introduction
2. Results
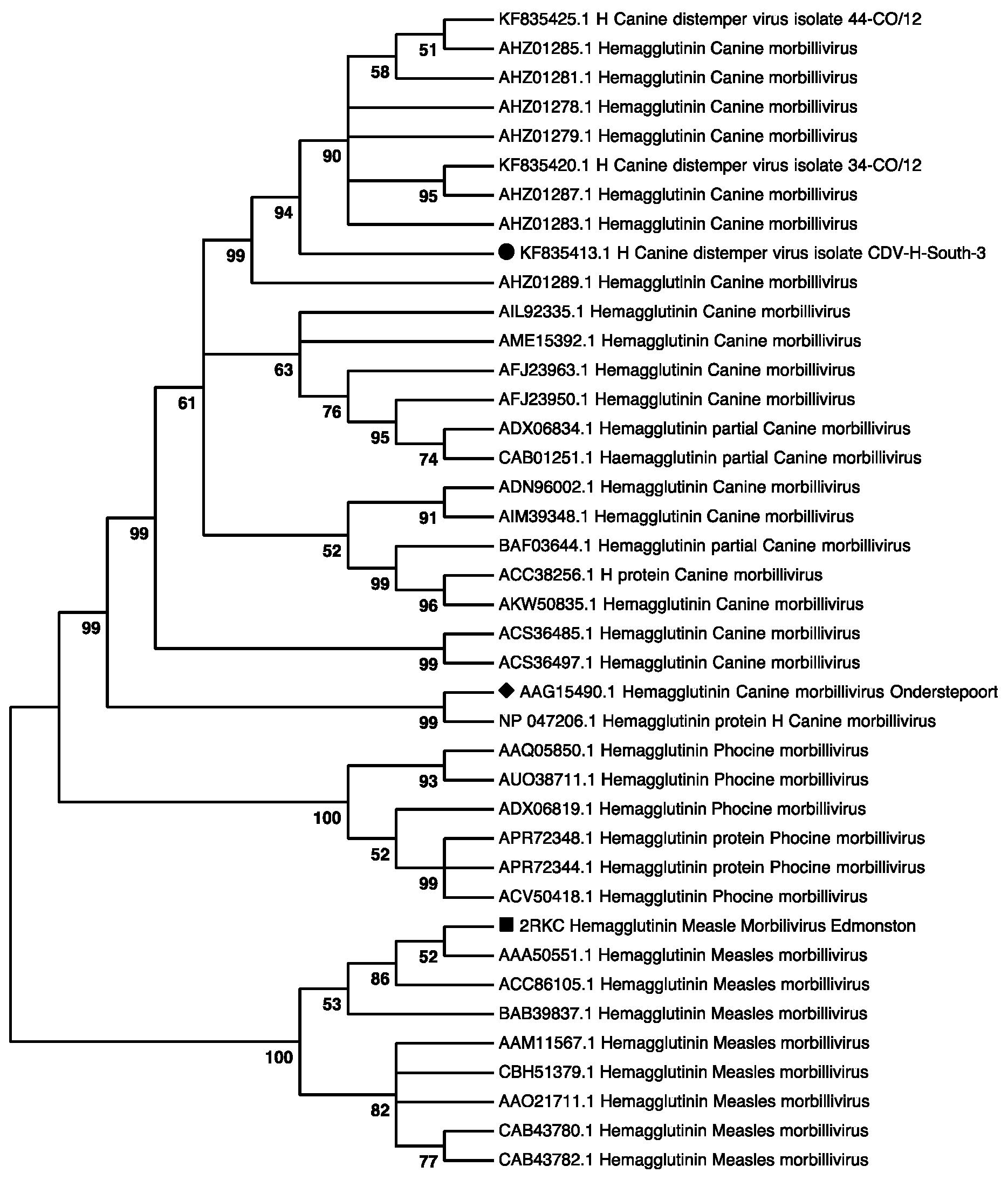
2.1. CDV-H-South-3 Protein Diverges from the MeV-H Protein Sequence
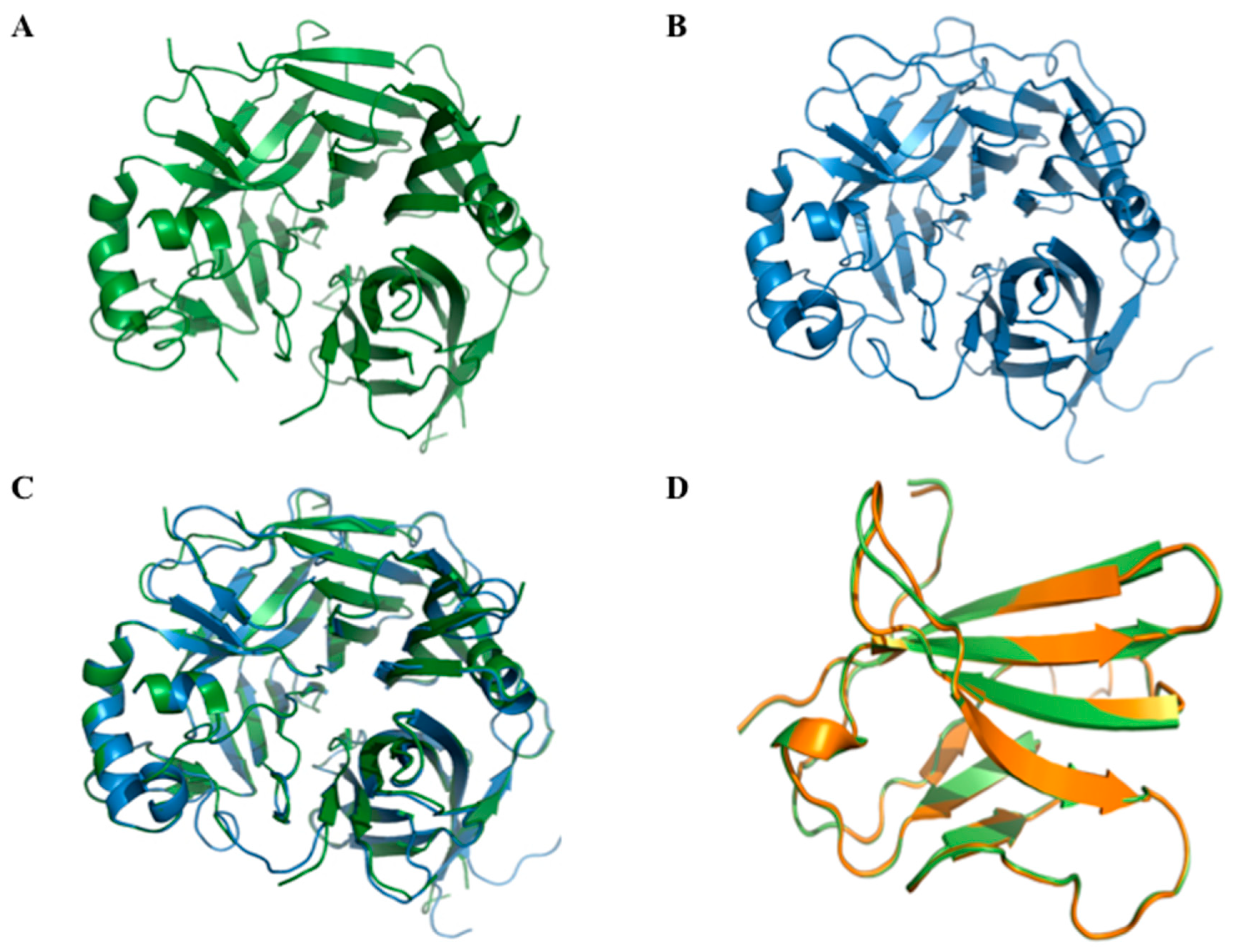
2.2. CDV-H-South-3 Homology 3D Model and Validation
2.3. Molecular Docking and Dynamics between Colombian CDV-H Protein and Cellular Receptors Showed a Potential Interaction with Orthologous Human Cellular Receptors
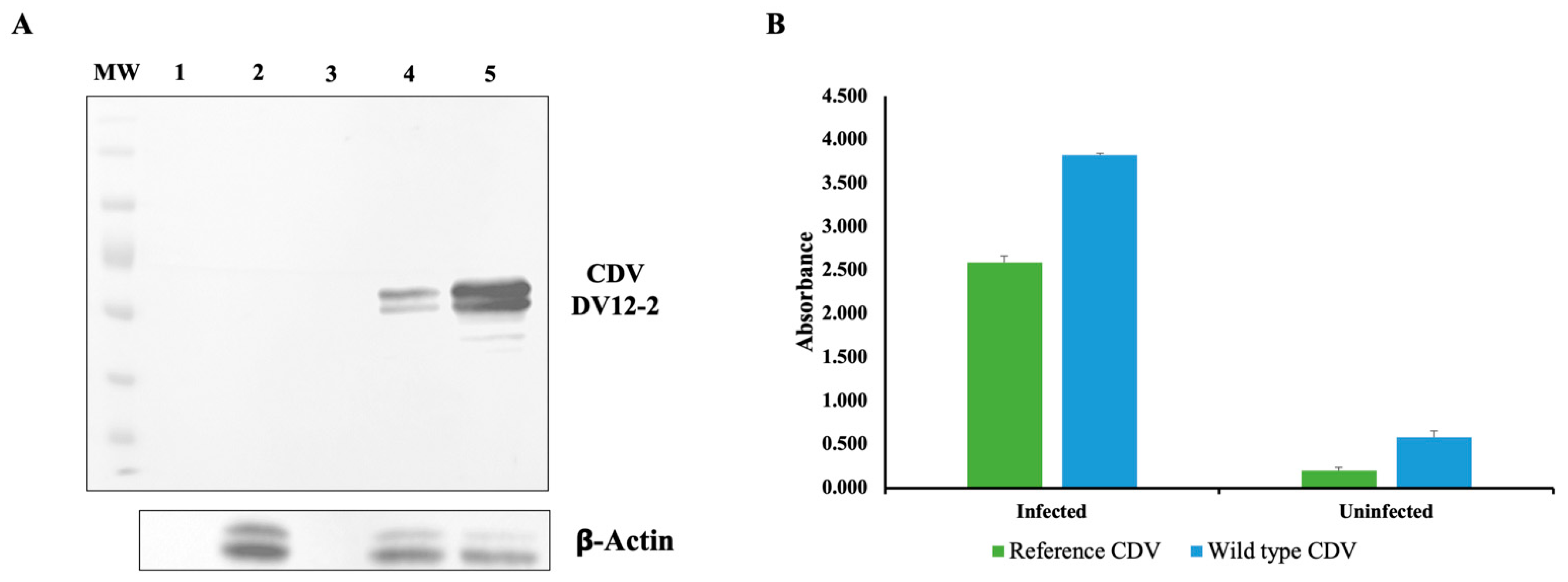
2.4. CDV from Colombian Circulating Strain Was Isolated from a Viremic Patient
2.5. CDV Proteins from Colombian Isolate Was Immunodetected in Cell Monolayers by Monoclonal Antibodies
2.6. CDV Colombian Strain Could Infect Human Cell Lines Efficiently
3. Discussion
4. Materials and Methods
4.1. H Protein Primary Structure Analysis, Homology Modelling and Models Validation
4.2. Molecular Docking Analysis
4.3. Molecular Dynamics Simulations
4.4. Cell Lines, Virus Isolation, and Viral Titration
4.5. Molecular Detection of Wild-Type Colombian CDV Strain by RT-PCR
4.6. CDV Protein Detection by CELL-ELISA and Western Blot
4.7. Reference and Wild-Type Colombian CDV Strain Infection in Human and Canine Cell Lines
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacLachlan, N.; Dubovi, E.; Fenner, F. Fenner’s Veterinary Virology, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 299–325. [Google Scholar]
- Martinez-Gutierrez, M.; Ruiz-Saenz, J. Diversity of susceptible hosts in canine distemper virus infection: A systematic review and data synthesis. BMC Vet. Res. 2016, 12, 78. [Google Scholar] [CrossRef]
- da Fontoura Budaszewski, R.; Streck, A.F.; Nunes Weber, M.; Maboni Siqueira, F.; Muniz Guedes, R.L.; Wageck Canal, C. Influence of vaccine strains on the evolution of canine distemper virus. Infect. Genet. Evol. 2016, 41, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae. In Fields Virology, 6th ed.; Wolters Kluwer Health Adis (ESP): Philadelphia, PA, USA, 2013. [Google Scholar]
- von Messling, V.; Zimmer, G.; Herrler, G.; Haas, L.; Cattaneo, R. The hemagglutinin of canine distemper virus determines tropism and cytopathogenicity. J. Virol. 2001, 75, 6418–6427. [Google Scholar] [CrossRef] [PubMed]
- von Messling, V.; Oezguen, N.; Zheng, Q.; Vongpunsawad, S.; Braun, W.; Cattaneo, R. Nearby clusters of hemagglutinin residues sustain SLAM-dependent canine distemper virus entry in peripheral blood mononuclear cells. J. Virol. 2005, 79, 5857–5862. [Google Scholar] [CrossRef]
- Pratakpiriya, W.; Seki, F.; Otsuki, N.; Sakai, K.; Fukuhara, H.; Katamoto, H.; Hirai, T.; Maenaka, K.; Techangamsuwan, S.; Lan, N.T.; et al. Nectin4 is an epithelial cell receptor for canine distemper virus and involved in neurovirulence. J. Virol. 2012, 86, 10207–10210. [Google Scholar] [CrossRef]
- da Fontoura Budaszewski, R.; von Messling, V. Morbillivirus Experimental Animal Models: Measles Virus Pathogenesis Insights from Canine Distemper Virus. Viruses 2016, 8, 274. [Google Scholar] [CrossRef]
- Noyce, R.S.; Bondre, D.G.; Ha, M.N.; Lin, L.T.; Sisson, G.; Tsao, M.S.; Richardson, C.D. Tumor cell marker PVRL4 (nectin 4) is an epithelial cell receptor for measles virus. PLoS Pathog. 2011, 7, e1002240. [Google Scholar] [CrossRef]
- Tatsuo, H.; Ono, N.; Yanagi, Y. Morbilliviruses use signaling lymphocyte activation molecules (CD150) as cellular receptors. J. Virol. 2001, 75, 5842–5850. [Google Scholar] [CrossRef] [PubMed]
- Rendon-Marin, S.; da Fontoura Budaszewski, R.; Canal, C.W.; Ruiz-Saenz, J. Tropism and molecular pathogenesis of canine distemper virus. Virol. J. 2019, 16, 30. [Google Scholar] [CrossRef] [PubMed]
- Langedijk, J.P.; Janda, J.; Origgi, F.C.; Orvell, C.; Vandevelde, M.; Zurbriggen, A.; Plattet, P. Canine distemper virus infects canine keratinocytes and immune cells by using overlapping and distinct regions located on one side of the attachment protein. J. Virol. 2011, 85, 11242–11254. [Google Scholar] [CrossRef]
- Duque-Valencia, J.; Sarute, N.; Olarte-Castillo, X.A.; Ruiz-Saenz, J. Evolution and Interspecies Transmission of Canine Distemper Virus-An Outlook of the Diverse Evolutionary Landscapes of a Multi-Host Virus. Viruses 2019, 11, 582. [Google Scholar] [CrossRef]
- Mochizuki, M.; Hashimoto, M.; Hagiwara, S.; Yoshida, Y.; Ishiguro, S. Genotypes of canine distemper virus determined by analysis of the hemagglutinin genes of recent isolates from dogs in Japan. J. Clin. Microbiol. 1999, 37, 2936–2942. [Google Scholar] [CrossRef]
- Anis, E.; Newell, T.K.; Dyer, N.; Wilkes, R.P. Phylogenetic analysis of the wild-type strains of canine distemper virus circulating in the United States. Virol. J. 2018, 15, 118. [Google Scholar] [CrossRef] [PubMed]
- Blixenkrone-Moller, M.; Svansson, V.; Appel, M.; Krogsrud, J.; Have, P.; Orvell, C. Antigenic relationships between field isolates of morbilliviruses from different carnivores. Arch. Virol. 1992, 123, 279–294. [Google Scholar] [CrossRef] [PubMed]
- Espinal, M.A.; Diaz, F.J.; Ruiz-Saenz, J. Phylogenetic evidence of a new canine distemper virus lineage among domestic dogs in Colombia, South America. Vet. Microbiol. 2014, 172, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Haas, L.; Martens, W.; Greiser-Wilke, I.; Mamaev, L.; Butina, T.; Maack, D.; Barrett, T. Analysis of the haemagglutinin gene of current wild-type canine distemper virus isolates from Germany. Virus Res. 1997, 48, 165–171. [Google Scholar] [CrossRef]
- Harder, T.C.; Kenter, M.; Vos, H.; Siebelink, K.; Huisman, W.; van Amerongen, G.; Orvell, C.; Barrett, T.; Appel, M.J.; Osterhaus, A.D. Canine distemper virus from diseased large felids: Biological properties and phylogenetic relationships. J. Gen. Virol. 1996, 77, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Iwatsuki, K.; Tokiyoshi, S.; Hirayama, N.; Nakamura, K.; Ohashi, K.; Wakasa, C.; Mikami, T.; Kai, C. Antigenic differences in the H proteins of canine distemper viruses. Vet. Microbiol. 2000, 71, 281–286. [Google Scholar] [CrossRef]
- Panzera, Y.; Calderon, M.G.; Sarute, N.; Guasco, S.; Cardeillac, A.; Bonilla, B.; Hernandez, M.; Francia, L.; Bedo, G.; La Torre, J.; et al. Evidence of two co-circulating genetic lineages of canine distemper virus in South America. Virus Res. 2012, 163, 401–404. [Google Scholar] [CrossRef]
- Riley, M.C.; Wilkes, R.P. Sequencing of emerging canine distemper virus strain reveals new distinct genetic lineage in the United States associated with disease in wildlife and domestic canine populations. Virol. J. 2015, 12, 219. [Google Scholar] [CrossRef]
- Nikolin, V.M.; Olarte-Castillo, X.A.; Osterrieder, N.; Hofer, H.; Dubovi, E.; Mazzoni, C.J.; Brunner, E.; Goller, K.V.; Fyumagwa, R.D.; Moehlman, P.D.; et al. Canine distemper virus in the Serengeti ecosystem: Molecular adaptation to different carnivore species. Mol. Ecol. 2017, 26, 2111–2130. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.J.; Yan, X.J.; Chai, X.L.; Martella, V.; Luo, G.L.; Zhang, H.L.; Gao, H.; Liu, Y.X.; Bai, X.; Zhang, L.; et al. Phylogenetic analysis of the haemagglutinin gene of canine distemper virus strains detected from breeding foxes, raccoon dogs and minks in China. Vet. Microbiol. 2010, 140, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Radtanakatikanon, A.; Keawcharoen, J.; Charoenvisal, N.T.; Poovorawan, Y.; Prompetchara, E.; Yamaguchi, R.; Techangamsuwan, S. Genotypic lineages and restriction fragment length polymorphism of canine distemper virus isolates in Thailand. Vet. Microbiol. 2013, 166, 76–83. [Google Scholar] [CrossRef]
- Duque-Valencia, J.; Forero-Munoz, N.R.; Diaz, F.J.; Martins, E.; Barato, P.; Ruiz-Saenz, J. Phylogenetic evidence of the intercontinental circulation of a Canine distemper virus lineage in the Americas. Sci. Rep. 2019, 9, 15747. [Google Scholar] [CrossRef]
- Duque-Valencia, J.; Diaz, F.J.; Ruiz-Saenz, J. Phylogenomic Analysis of Two Co-Circulating Canine Distemper Virus Lineages in Colombia. Pathogens 2019, 9, 26. [Google Scholar] [CrossRef]
- Ke, G.M.; Ho, C.H.; Chiang, M.J.; Sanno-Duanda, B.; Chung, C.S.; Lin, M.Y.; Shi, Y.Y.; Yang, M.H.; Tyan, Y.C.; Liao, P.C.; et al. Phylodynamic analysis of the canine distemper virus hemagglutinin gene. BMC Vet. Res. 2015, 11, 164. [Google Scholar] [CrossRef]
- Hashiguchi, T.; Ose, T.; Kubota, M.; Maita, N.; Kamishikiryo, J.; Maenaka, K.; Yanagi, Y. Structure of the measles virus hemagglutinin bound to its cellular receptor SLAM. Nat. Struct. Mol. Biol. 2011, 18, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lu, G.; Qi, J.; Li, Y.; He, Y.; Xu, X.; Shi, J.; Zhang, C.W.; Yan, J.; Gao, G.F. Structure of measles virus hemagglutinin bound to its epithelial receptor nectin-4. Nat. Struct. Mol. Biol. 2013, 20, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, Y.; Ochikubo, F.; Matsubara, Y.; Tsuruoka, H.; Ishii, M.; Shirota, K.; Nomura, Y.; Sugiyama, M.; Yamanouchi, K. Natural infection with canine distemper virus in a Japanese monkey (Macaca fuscata). Vet. Microbiol. 1989, 20, 193–205. [Google Scholar] [CrossRef]
- Otsuki, N.; Nakatsu, Y.; Kubota, T.; Sekizuka, T.; Seki, F.; Sakai, K.; Kuroda, M.; Yamaguchi, R.; Takeda, M. The V protein of canine distemper virus is required for virus replication in human epithelial cells. PLoS ONE 2013, 8, e82343. [Google Scholar]
- Fukuhara, H.; Ito, Y.; Sako, M.; Kajikawa, M.; Yoshida, K.; Seki, F.; Mwaba, M.H.; Hashiguchi, T.; Higashibata, M.A.; Ose, T.; et al. Specificity of Morbillivirus Hemagglutinins to Recognize SLAM of Different Species. Viruses 2019, 11, 761. [Google Scholar] [CrossRef]
- Bieringer, M.; Han, J.W.; Kendl, S.; Khosravi, M.; Plattet, P.; Schneider-Schaulies, J. Experimental adaptation of wild-type canine distemper virus (CDV) to the human entry receptor CD150. PLoS ONE 2013, 8, e57488. [Google Scholar]
- Krieger, E.; Nabuurs, S.B.; Vriend, G. Homology modeling. Methods Biochem. Anal. 2003, 44, 509–523. [Google Scholar] [PubMed]
- Quintero-Gil, C.; Rendon-Marin, S.; Martinez-Gutierrez, M.; Ruiz-Saenz, J. Origin of Canine Distemper Virus: Consolidating Evidence to Understand Potential Zoonoses. Front. Microbiol. 2019, 10, 1982. [Google Scholar] [CrossRef]
- Cosby, S.L.; Weir, L. Measles vaccination: Threat from related veterinary viruses and need for continued vaccination post measles eradication. Hum. Vaccin Immunother. 2018, 14, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Seki, F.; Yamamoto, Y.; Nao, N.; Tokiwa, H. Animal morbilliviruses and their cross-species transmission potential. Curr. Opin. Virol. 2020, 41, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Chattopadhyay, K.; Lazar-Molnar, E.; Yan, Q.; Rubinstein, R.; Zhan, C.; Vigdorovich, V.; Ramagopal, U.A.; Bonanno, J.; Nathenson, S.G.; Almo, S.C. Sequence, structure, function, immunity: Structural genomics of costimulation. Immunol Rev. 2009, 229, 356–386. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.K.; Rajak, K.K.; Bhatt, M.; Kumar, A.; Chakravarti, S.; Sankar, M.; Muthuchelvan, D.; Kumar, R.; Khulape, S.; Singh, R.P.; et al. Comparative sequence analysis of morbillivirus receptors and its implication in host range expansion. Can. J. Microbiol. 2019, 65, 783–794. [Google Scholar] [CrossRef]
- de Vries, R.D.; Ludlow, M.; Verburgh, R.J.; van Amerongen, G.; Yuksel, S.; Nguyen, D.T.; McQuaid, S.; Osterhaus, A.D.; Duprex, W.P.; de Swart, R.L. Measles vaccination of nonhuman primates provides partial protection against infection with canine distemper virus. J. Virol. 2014, 88, 4423–4433. [Google Scholar] [CrossRef] [PubMed]
- Nambulli, S.; Sharp, C.R.; Acciardo, A.S.; Drexler, J.F.; Duprex, W.P. Mapping the evolutionary trajectories of morbilliviruses: What, where and whither. Curr. Opin. Virol. 2016, 16, 95–105. [Google Scholar] [CrossRef]
- Otsuki, N.; Sekizuka, T.; Seki, F.; Sakai, K.; Kubota, T.; Nakatsu, Y.; Chen, S.; Fukuhara, H.; Maenaka, K.; Yamaguchi, R.; et al. Canine distemper virus with the intact C protein has the potential to replicate in human epithelial cells by using human nectin4 as a receptor. Virology 2013, 435, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Uhl, E.W.; Kelderhouse, C.; Buikstra, J.; Blick, J.P.; Bolon, B.; Hogan, R.J. New world origin of canine distemper: Interdisciplinary insights. Int. J. Paleopathol. 2019, 24, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Nagata, N.; Ami, Y.; Seki, F.; Suzaki, Y.; Iwata-Yoshikawa, N.; Suzuki, T.; Fukushi, S.; Mizutani, T.; Yoshikawa, T.; et al. Lethal canine distemper virus outbreak in cynomolgus monkeys in Japan in 2008. J. Virol. 2013, 87, 1105–1114. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Nakano, S.; Seki, F.; Shigeta, Y.; Ito, S.; Tokiwa, H.; Takeda, M. Computational Analysis Reveals a Critical Point Mutation in the N-Terminal Region of the Signaling Lymphocytic Activation Molecule Responsible for the Cross-Species Infection with Canine Distemper Virus. Molecules 2021, 26, 1262. [Google Scholar] [CrossRef]
- Seki, F.; Yamamoto, Y.; Fukuhara, H.; Ohishi, K.; Maruyama, T.; Maenaka, K.; Tokiwa, H.; Takeda, M. Measles Virus Hemagglutinin Protein Establishes a Specific Interaction With the Extreme N-Terminal Region of Human Signaling Lymphocytic Activation Molecule to Enhance Infection. Front. Microbiol. 2020, 11, 1830. [Google Scholar] [CrossRef] [PubMed]
- Pfeffermann, K.; Dorr, M.; Zirkel, F.; von Messling, V. Morbillivirus Pathogenesis and Virus-Host Interactions. Adv. Virus Res. 2018, 100, 75–98. [Google Scholar] [PubMed]
- Reddy, T.; Sansom, M.S. Computational virology: From the inside out. Biochim. Biophys. Acta. 2016, 1858, 1610–1618. [Google Scholar] [CrossRef]
- Huber, R.G.; Marzinek, J.K.; Holdbrook, D.A.; Bond, P.J. Multiscale molecular dynamics simulation approaches to the structure and dynamics of viruses. Prog. Biophys. Mol. Biol. 2017, 128, 121–132. [Google Scholar] [CrossRef]
- Behera, A.K.; Chandra, I.; Cherian, S.S. Molecular dynamics simulation of the effects of single (S221P) and double (S221P and K216E) mutations in the hemagglutinin protein of influenza A H5N1 virus: A study on host receptor specificity. J. Biomol. Struct. Dyn. 2016, 34, 2054–2067. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Botlani, M.; Varma, S. Water dynamics at protein–protein interfaces: Molecular dynamics study of virus–host receptor complexes. J. Phys. Chem. B. 2014, 118, 14795–14807. [Google Scholar]
- Chan, K.W.; Hsieh, H.H.; Wang, H.C.; Lee, Y.J.; Sung, M.H.; Wong, M.L.; Hsu, W.L. Identification, expression and antigenic analysis of recombinant hemagglutinin proteins of canine distemper virus. J. Virol. Methods. 2009, 155, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.J.; Shek, W.R.; Shesberadaran, H.; Norrby, E. Measles virus and inactivated canine distemper virus induce incomplete immunity to canine distemper. Arch. Virol. 1984, 82, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.L.; Vitamvas, J.A.; Merry, D.L., Jr.; Beckenhauer, W.H. Immune response of pups to modified live-virus canine distemper-measles vaccine. Am. J. Vet. Res. 1972, 33, 1447–1456. [Google Scholar] [PubMed]
- Qiu, W.; Zheng, Y.; Zhang, S.; Fan, Q.; Liu, H.; Zhang, F.; Wang, W.; Liao, G.; Hu, R. Canine distemper outbreak in rhesus monkeys, China. Emerg. Infect. Dis. 2011, 17, 1541–1543. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef]
- Zhang, Y.; Skolnick, J. TM-align: A protein structure alignment algorithm based on the TM-score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef]
- Berger, J.J.; Barnard, R.J. Effect of diet on fat cell size and hormone-sensitive lipase activity. J. Appl. Physiol. 1999, 87, 227–232. [Google Scholar] [CrossRef]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B., 3rd; de Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Calpha geometry: Phi,psi and Cbeta deviation. Proteins 2003, 50, 437–450. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Vajda, S.; Yueh, C.; Beglov, D.; Bohnuud, T.; Mottarella, S.E.; Xia, B.; Hall, D.R.; Kozakov, D. New additions to the ClusPro server motivated by CAPRI. Proteins 2017, 85, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein-protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Visser, I.K.; Mamaev, L.; Goatley, L.; van Bressem, M.F.; Osterhaust, A.D. Dolphin and porpoise morbilliviruses are genetically distinct from phocine distemper virus. Virology 1993, 193, 1010–1012. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Z Value | Favorable Region (%) | TM Value | Aligned AA |
|---|---|---|---|---|
| CDV-H-South-3 | −7.00 | 96.9 | 0.983 | 397 |
| CYN07-dV | −6.82 | 98.1 | 0.986 | 399 |
| Onderstepoort | −6.53 | 97.2 | 0.985 | 397 |
| cSLAM | −4.56 | 97.2 | 0.992 | 104 |
| cNectin-4 | −4.55 | 98.2 | 0.998 | 114 |
| Complex | Cluster | Members |
|---|---|---|
| cSLAM–CDV-H-South-3 | 2 | 49 |
| hSLAM–CDV-H- South-3 | 6 | 32 |
| cNectin-4–CDV-H- South-3 | 3 | 54 |
| hNectin-4–CDV-H- South-3 | 4 | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rendon-Marin, S.; Quintero-Gil, C.; Guerra, D.; Muskus, C.; Ruiz-Saenz, J. Canine Morbillivirus from Colombian Lineage Exhibits In Silico and In Vitro Potential to Infect Human Cells. Pathogens 2021, 10, 1199. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10091199
Rendon-Marin S, Quintero-Gil C, Guerra D, Muskus C, Ruiz-Saenz J. Canine Morbillivirus from Colombian Lineage Exhibits In Silico and In Vitro Potential to Infect Human Cells. Pathogens. 2021; 10(9):1199. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10091199
Chicago/Turabian StyleRendon-Marin, Santiago, Carolina Quintero-Gil, Diego Guerra, Carlos Muskus, and Julian Ruiz-Saenz. 2021. "Canine Morbillivirus from Colombian Lineage Exhibits In Silico and In Vitro Potential to Infect Human Cells" Pathogens 10, no. 9: 1199. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens10091199