

A First Report of Molecular Typing, Virulence Traits, and Phenotypic and Genotypic Resistance Patterns of Newly Emerging XDR and MDR Aeromonas veronii in Mugil seheli

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Sampling
2.3. Clinical and Postmortem Examinations
2.4. Isolation and Identification of A. veronii
2.5. 16S rRNA Gene Sequencing and Phylogenetic Analyses
2.6. Antibiogram of the Recovered A. veronii Isolates
2.7. Determination of Virulence and Antimicrobial Resistance Genes in the Recovered A. veronii Strains
2.8. Pathogenicity Test
2.8.1. Fish Sampling and Accommodation Period
2.8.2. Experimental Setup
2.9. Statistical Analyses
3. Results
3.1. Clinical and Post-Mortem Findings
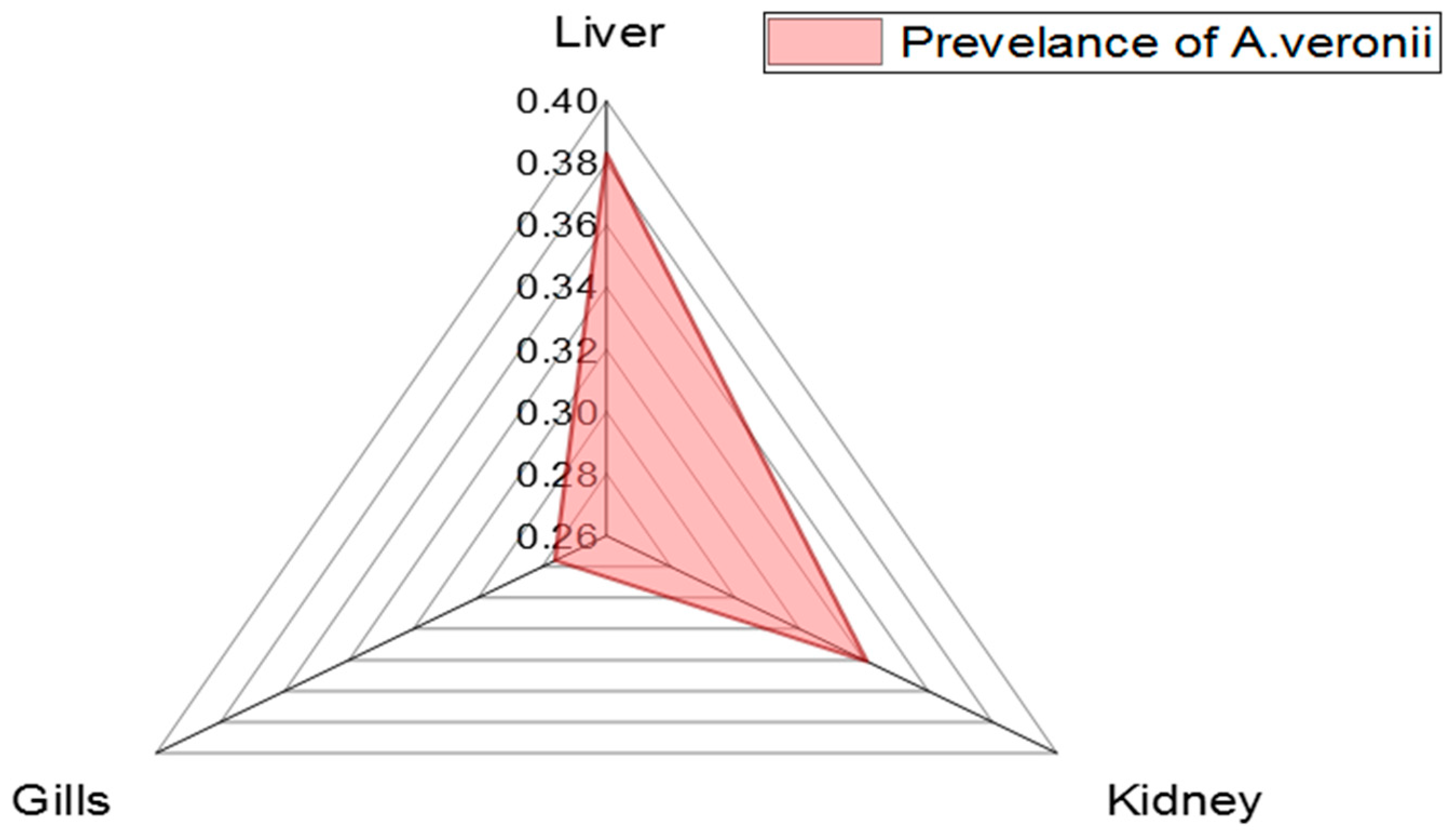
3.2. Phenotypic Features and the Prevalence of A. veronii in the Examined M. seheli
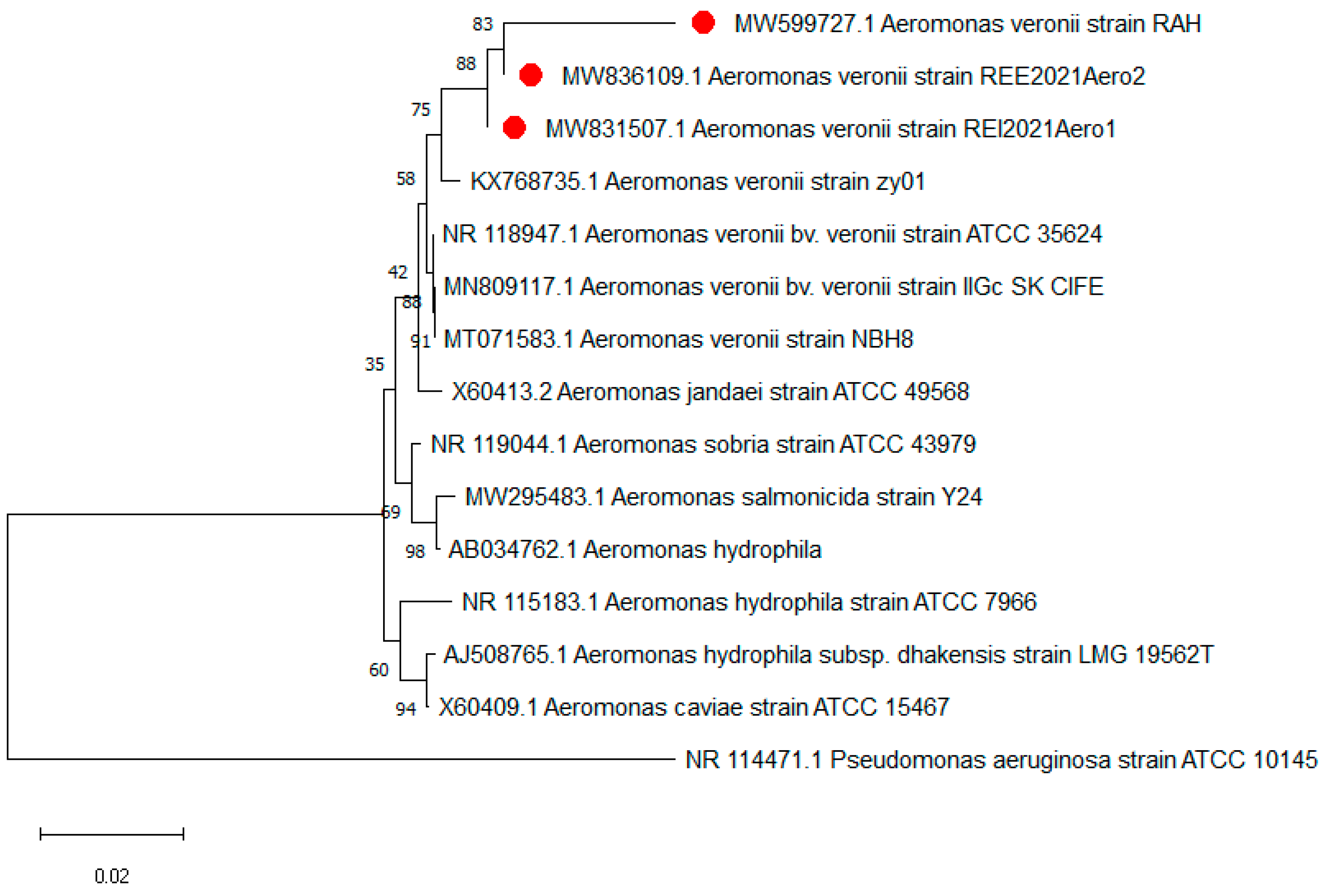
3.3. 16srRNA Gene Sequencing and Phylogenetic Analyses
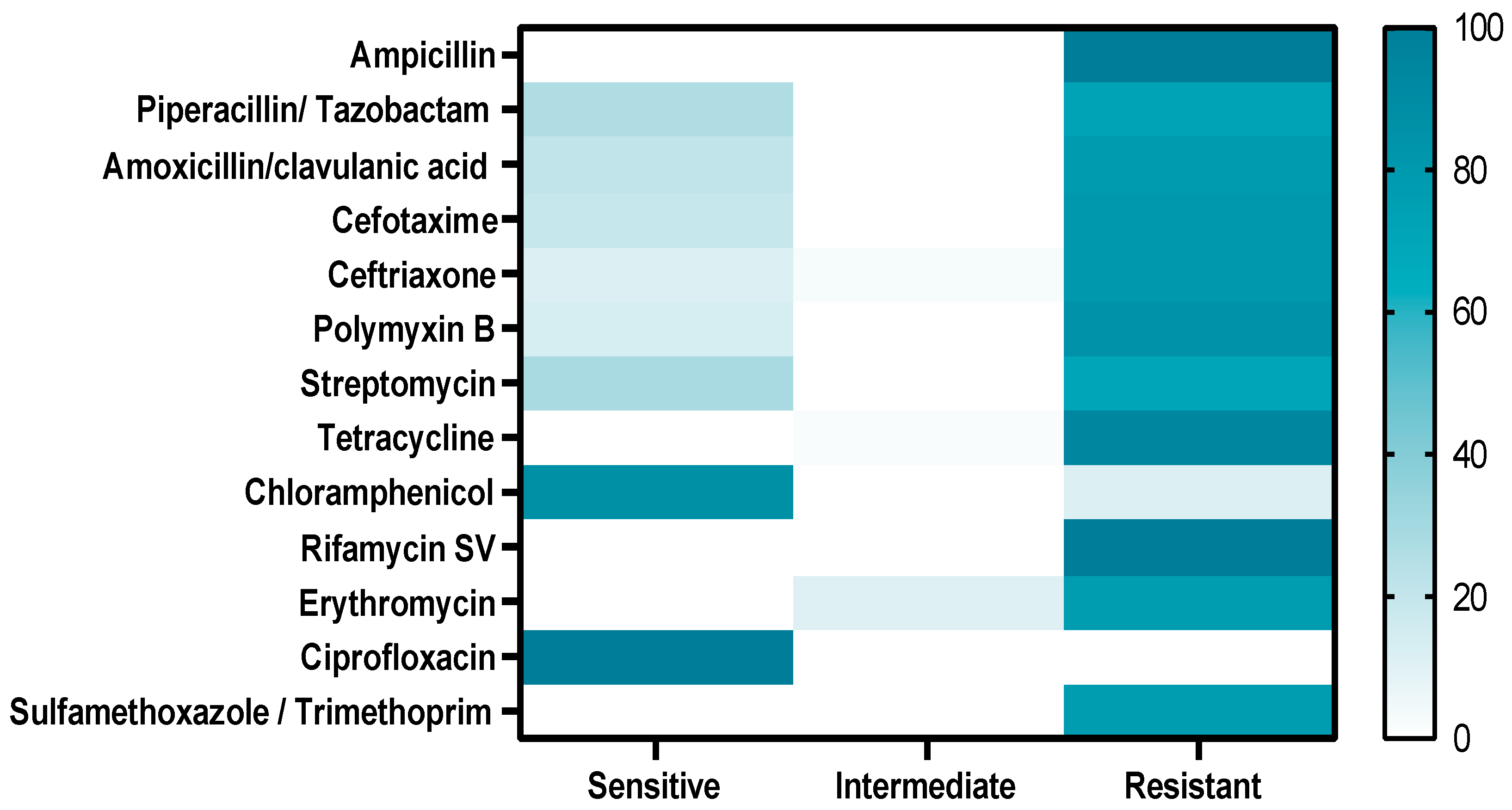
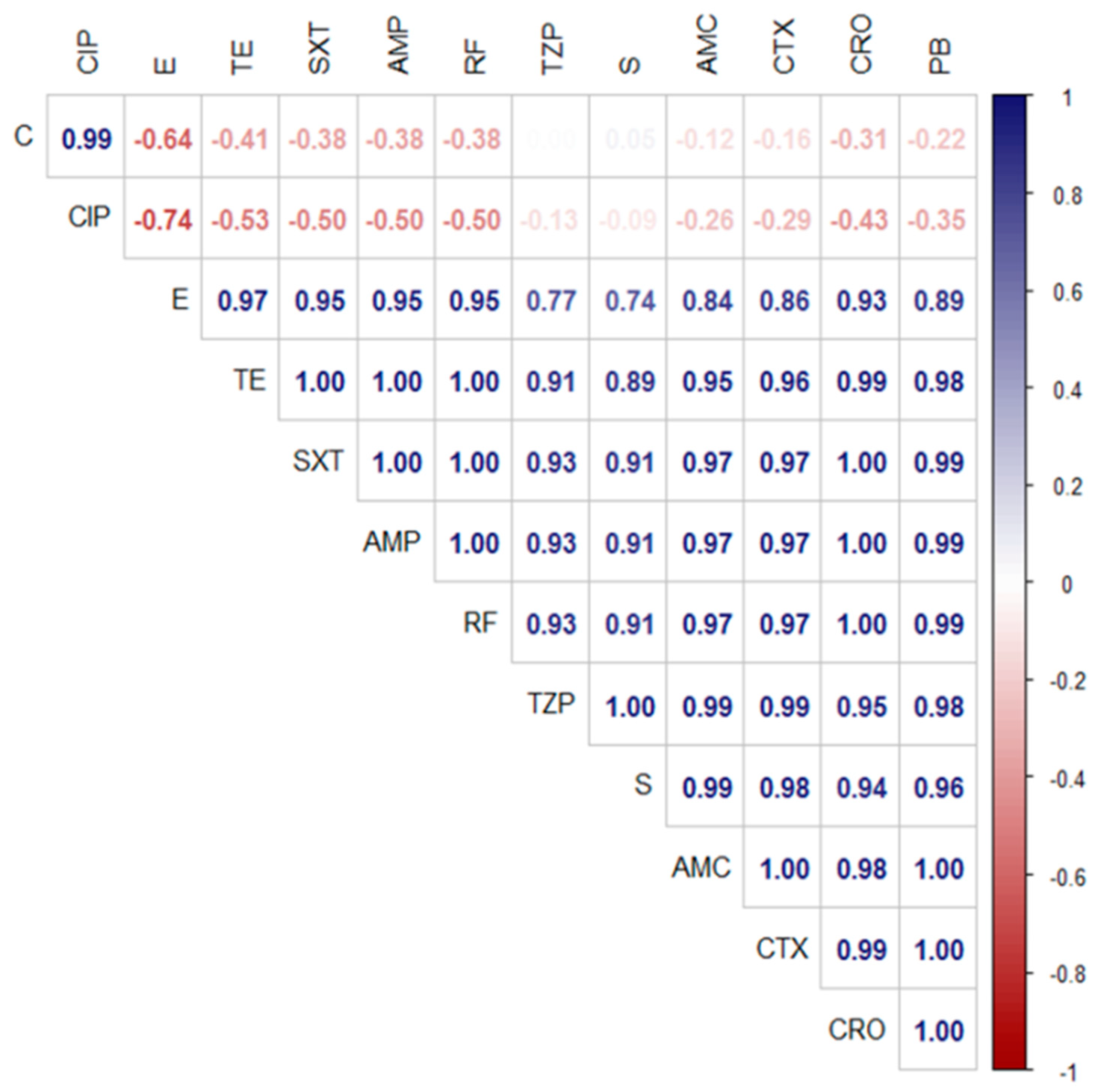
3.4. Antimicrobial Resistance Profiles of the Retrieved A. veronii Isolates
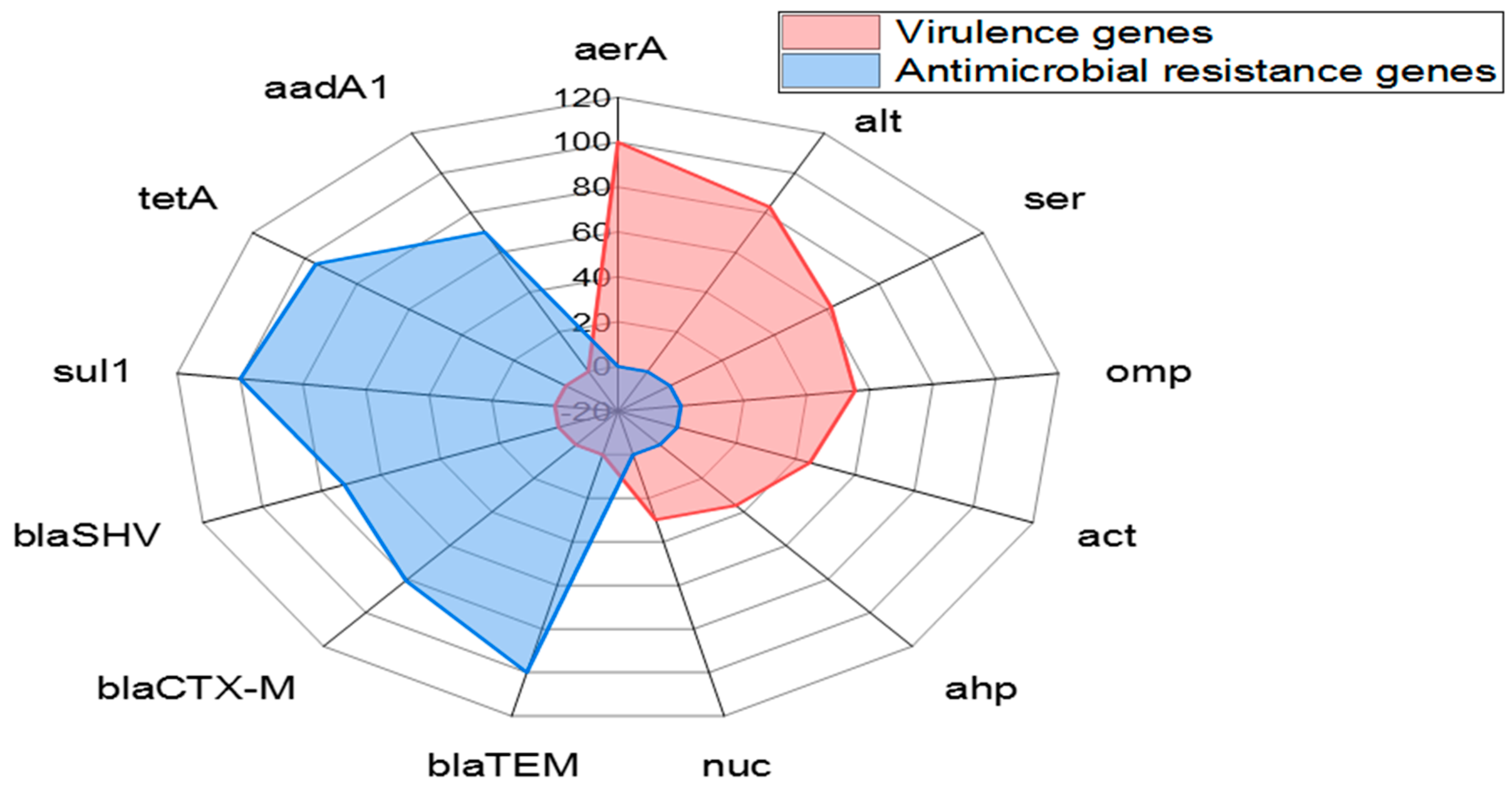
3.5. Dissemination of Virulence-Determinant and Antimicrobial Resistance Genes in the Emerging A. veronii Strains
3.6. Genotypic and Phenotypic Multidrug-Resistance Patterns of the Emerging A. veronii Strains
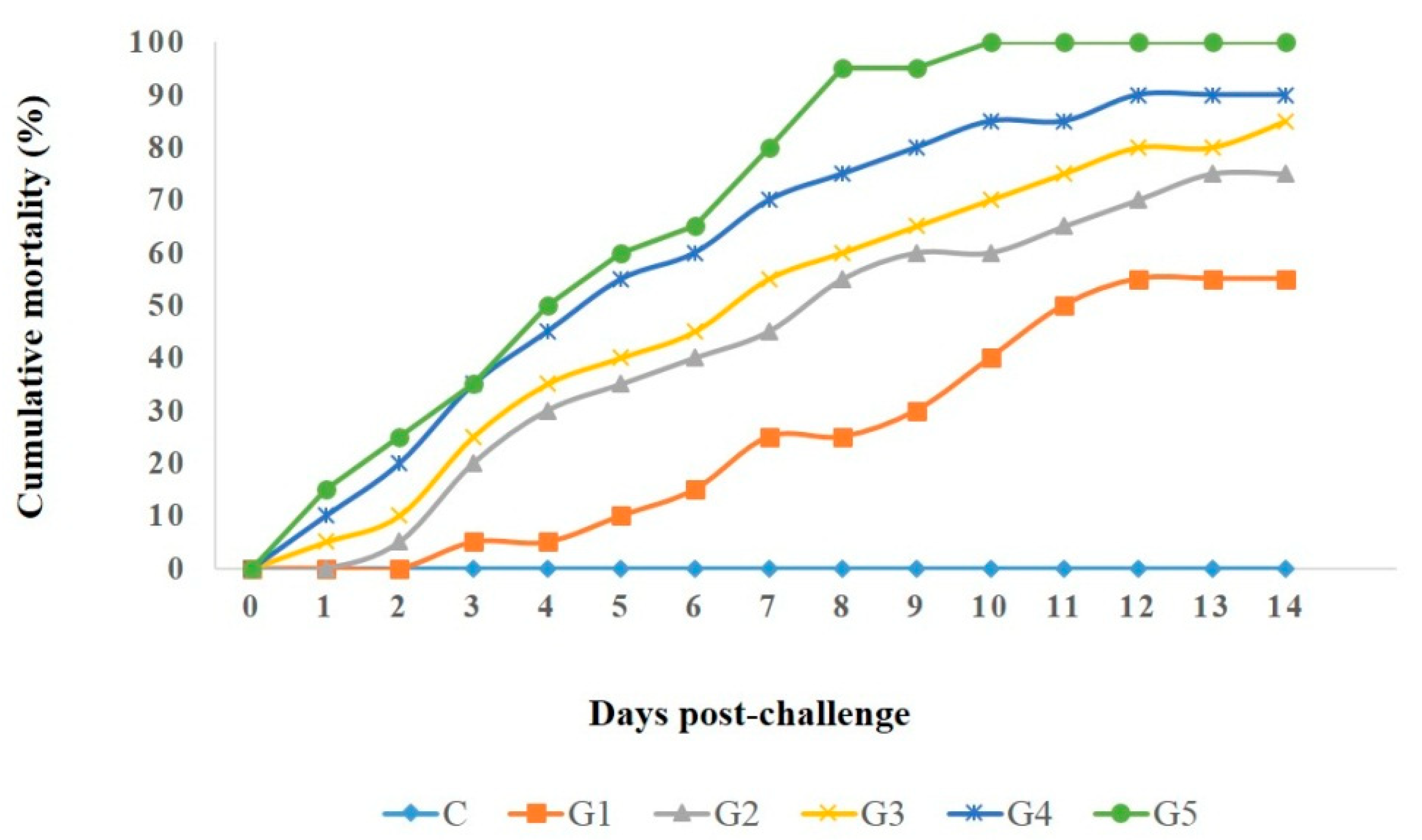
3.7. Pathogenicity Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture. Contributing to Food Security and Nutrition for All; FAO: Rome, Italy, 2016. [Google Scholar]
- Abouelmaatti, R.R.; Algammal, A.M.; LI, X.; MA, J.; Abdelnaby, E.A.; Elfeil, W.M. Cloning and analysis of Nile tilapia Toll-like receptors type-3 mRNA. Centr. Eur. J. Immunol. 2013, 38, 277–282. [Google Scholar] [CrossRef]
- Shaalan, M.; El-Mahdy, M.; Saleh, M.; El-Matbouli, M. Aquaculture in Egypt: Insights on the Current Trends and Future Perspectives for Sustainable Development. Rev. Fish. Sci. Aquac. 2018, 26, 99–110. [Google Scholar] [CrossRef]
- Mehanna, S.F.; El-Sherbeny, A.S.; El-Mor, M.; Eid, N.M. Age, Growth and Mortality of Liza Carinata Valenciennes, 1836 (Pisces: Mugilidae) in Bitter Lakes, Suez Canal, Egypt. Egypt. J. Aquat. Biol. Fish. 2019, 23, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Nelson, J.S. Fishes of the World, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Abd El Latif, A.M.A.; Elabd, H.; Amin, A.; Eldeen, A.I.N.; Shaheen, A.A. High Mortalities Caused by Aeromonas veronii: Identification, Pathogenicity, and Histopathological studies in Oreochromis niloticus. Aquac. Int. 2019, 27, 1725–1737. [Google Scholar] [CrossRef]
- Nawaz, M.; Khan, S.A.; Khan, A.A.; Sung, K.; Tran, Q.; Kerdahi, K.; Steele, R. Detection and Characterization of Virulence Genes and Integrons in Aeromonas veronii Isolated from Catfish. Food Microbiol. 2010, 27, 327–331. [Google Scholar] [CrossRef]
- Hoai, T.; Trang, T.; Van Tuyen, N.; Aquaculture, N.G. Aeromonas veronii Caused Disease and Mortality in Channel Catfish in Vietnam. Aquaculture 2019, 513, 734425. [Google Scholar] [CrossRef]
- Shameena, S.S.; Kumar, K.; Kumar, S.; Kumar, S.; Rathore, G. Virulence Characteristics of Aeromonas veronii Biovars Isolated from Infected Freshwater Goldfish (Carassius Auratus). Aquaculture 2020, 518, 734819. [Google Scholar] [CrossRef]
- Cai, S.; Wu, Z.; Jian, J.; Lu, Y.; Tang, J. Characterization of Pathogenic Aeromonas veronii Bv. Veronii Associated with Ulcerative Syndrome from Chinese Longsnout Catfish (Leiocassis Longirostris Günther). SciELO Bras. 2012, 43, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Raza, S.H.A.; Yang, B.; Sun, Y.; Wang, G.; Sun, W.; Qian, A.; Wang, C.; Kang, Y.; Shan, X. Aeromonas veronii Infection in Commercial Freshwater Fish: A Potential Threat to Public Health. Animals 2020, 10, 608. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Abbott, S.L. The Genus Aeromonas: Taxonomy, Pathogenicity, and Infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef]
- Fernandez-Bravo, A.; Fort-Gallifa, I.; Ballester, F.; Pujol, I.; Gomez-Bertomeu, F.; Dominguez, M.; Micó, M.; Alcoceba, E.; Simó-Sisó, J.M.; Figueras, M.J. A Case of Aeromonas Trota in an Immunocompromised Patient with Diarrhea. Microorganisms 2020, 8, 399. [Google Scholar] [CrossRef] [Green Version]
- Algammal, A.; Mohamed, M.; Tawfiek, B.; Pathogens, W.H. Molecular Typing, Antibiogram and PCR-RFLP Based Detection of Aeromonas Hydrophila Complex Isolated from Oreochromis niloticus. Pathogens 2020, 9, 238. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zhang, X.; Gao, X.; Jiang, Q.; Wen, Y.; Lin, L. Characterization of Virulence Properties of Aeromonas veronii Isolated from Diseased Gibel Carp (Carassius Gibelio). Int. J. Mol. Sci. 2016, 17, 496. [Google Scholar] [CrossRef]
- Hossain, S.; De-Silva, B.C.J.; Dahanayake, P.S.; Heo, G.J. Characterization of Virulence Properties and Multi-Drug Resistance Profiles in Motile Aeromonas Spp. Isolated from Zebrafish (Danio Rerio). Lett. Appl. Microbiol. 2018, 67, 598–605. [Google Scholar] [CrossRef]
- Zhang, D.X.; Kang, Y.H.; Song, M.F.; Shu, H.P.; Guo, S.N.; Jia, J.P.; Tao, L.T.; Zhao, Z.L.; Zhang, L.; Wang, C.F.; et al. Identity and Virulence Properties of Aeromonas Isolates from Healthy Northern Snakehead (Channa Argus) in China. Lett. Appl. Microbiol. 2019, 69, 100–109. [Google Scholar] [CrossRef]
- Abu-Elala, N.; Abdelsalam, M.; Marouf, S.; Setta, A.; Abu-Elala, N.; Abdel-Salam, M. Comparative Analysis of Virulence Genes, Antibiotic Resistance and GyrB-based Phylogeny of Motile Aeromonas Species Isolates from Nile Tilapia and Domestic Fowl. Lett. Appl. Microbiol. 2015, 61, 429–436. [Google Scholar] [CrossRef]
- Castro-Escarpulli, G.; Aguilera, G.; Castro-Escarpulli, G.; Figueras, M.J.; Aguilera-Arreola, G.; Soler, L.; Fernández-Rendón, E.; Aparicio, G.O.; Guarro, J.; Chacón, M.R. Characterisation of Aeromonas Spp. Isolated from Frozen Fish Intended for Human Consumption in Mexico. Int. J. Food Microbiol. 2003, 84, 41–49. [Google Scholar] [CrossRef]
- Algammal, A.M.; Hashem, H.R.; Al-Otaibi, A.S.; Alfifi, K.J.; El-Dawody, E.M.; Mahrous, E.; Hetta, H.F.; El-Kholy, A.W.; Ramadan, H.; El-Tarabili, R.M. Emerging MDR-Mycobacterium avium subsp. avium in house-reared domestic birds as the first report in Egypt. BMC Microbiol. 2021, 21, 237. [Google Scholar] [CrossRef]
- Kareem, S.M.; Al-kadmy, I.M.S.; Kazaal, S.S.; Mohammed Ali, A.N.; Aziz, S.N.; Makharita, R.R.; Algammal, A.M.; Al-Rejaie, S.; Behl, T.; Batiha, G.E.-S.; et al. Detection of gyrA and parC Mutations and Prevalence of Plasmid-Mediated Quinolone Resistance Genes in Klebsiella pneumoniae. Infect. Drug Resist. 2021, 14, 555–563. [Google Scholar] [CrossRef]
- Algammal, A.M.; Alfifi, K.J.; Mabrok, M.; Alatawy, M.; Abdel-moneam, D.A.; Alghamdi, S.; Azab, M.M.; Ibrahim, R.A.; Hetta, H.F.; El-Tarabili, R.M. Newly Emerging MDR B. cereus in Mugil Seheli as the First Report Commonly Harbor Nhe, Hbl, CytK, and Pc-Plc Virulence Genes and bla1, bla2, tetA, and ermA. Infect. Drug Resist. 2022, 15, 2167–2185. [Google Scholar] [CrossRef]
- Cabello, F.C. Heavy Use of Prophylactic Antibiotics in Aquaculture: A Growing Problem for Human and Animal Health and for the Environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- WHO. Report of a Joint FAO/OIE/WHO Expert Consultation on Antimicrobial Use in Aquaculture and Antimicrobial Resistance; WHO: Seoul, Korea, 2006.
- Colquhoun, D.J.; Aarflot, L.; Melvold, C.F. GyrA and Par C Mutations and Associated Quinolone Resistance in Vibrio anguillarum Serotype O2b Strains Isolated from Farmed Atlantic Cod (Gadus Morhua) in Norway. Antimicrob. Agents Chemother. 2007, 51, 2597–2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, B.; Austin, D. Characteristics of the pathogens: Gram-negative bacteria. In Bacterial Fish Pathogens; Springer: Berlin/Heidelberg, Germany, 2007; pp. 81–150. [Google Scholar]
- Noga, E.J. Fish Disease: Diagnosis and Treatment; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Monir, M.S.; Borty, S.C.; Bagum, N.; Rahman, M.K.; Islam, M.A.; Mahmud, Y. Identification of Pathogenic Bacteria Isolated from Diseased Stinging Catfish, Shing (Heteropneustes Fossilis) Cultured in Greater Mymensingh, Bangladesh. Asian Australas. J. Biosci. Biotechnol. 2016, 1, 116–124. [Google Scholar]
- Eissa, I.A.M.; Maather, E.; Mona, S.; Desuky, E.; Mona, Z.; Bakry, M. Aeromonas veronii biovar sobria a causative agent of mass mortalities in cultured Nile Tilapia in El-Sharkia governorate, Egypt. Life Sci. J. 2015, 12, 90–97. [Google Scholar]
- Gordon, L.; Giraud, E.; Ganiè, J.-P.; Armand, F.; Bouju-Albert, A.; De La Cotte, N.; Mangion, C.; Le Bris, H. Antimicrobial Resistance Survey in a River Receiving Effluents from Freshwater Fish Farms. J. Appl. Microbiol. 2007, 102, 1167–1176. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for Inferring Very Large Phylogenies by Using the Neighbor-Joining Method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- CLSI, Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing for Bacteria Isolated from Animals, 4th ed.; CLSI Supplement VET08; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Krumperman, P.H. Multiple Antibiotic Resistance Indexing of Escherichia Coli to Identify High-Risk Sources of Fecal Contamination of Foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, S.T.L.; Veneroni-Gouveia, G.; Costa, M.M. Molecular Characterization of Virulence Factors in Aeromonas hydrophila Obtained from Fish. Pesqui. Vet. Bras. 2012, 32, 701–706. [Google Scholar] [CrossRef]
- Nam, I.; Joh, K. Rapid Detection of Virulence Factors of Aeromonas Isolated from a Trout Farm by Hexaplex-PCR. J. Microbiol. 2007, 45, 297–304. [Google Scholar]
- Hu, M.; Wang, N.; Pan, Z.H.; Lu, C.P.; Liu, Y.J. Identity and Virulence Properties of Aeromonas Isolates from Diseased Fish, Healthy Controls and Water Environment in China. Lett. Appl. Microbiol. 2012, 55, 224–233. [Google Scholar] [CrossRef]
- Wang, H. Comparative Study between Biological Characteristics of the Different Animal Species Aeromonas veronii and Four Virulence Genes. Master’s Thesis, Agricultural University, Jilin, China, 2014. [Google Scholar]
- Colom, K.; Pèrez, J.; Alonso, R.; Fernández-Aranguiz, A.; Lariňo, E.; Cisterna, R. Simple and reliable multiplex PCR assay for detection of blaTEM, blaSHV and blaOXA-1 genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef]
- Archambault, M.; Petrov, P.; Hendriksen, R.S.; Asseva, G.; Bangtrakulnonth, A.; Hasman, H.; Aarestrup, F.M. Molecular Characterization and Occurrence of Extended-Spectrum β-Lactamase Resistance Genes among Salmonella Enterica Serovar Corvallis from Thailand, Bulgaria, and Denmark. Microb. Drug Resist. 2006, 12, 192–198. [Google Scholar] [CrossRef]
- Randall, L.P.; Cooles, S.W.; Osborn, M.K.; Piddock, L.J.V.; Woodward, M.J. Antibiotic Resistance Genes, Integrons and Multiple Antibiotic Resistance in Thirty-Five Serotypes of Salmonella Enterica isolated from Humans and Animals in the UK. J. Antimicrob. Chemother. 2004, 53, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Ibekwe, A.M.; Murinda, S.E.; Graves, A.K. Genetic Diversity and Antimicrobial Resistance of Escherichia coli from Human and Animal Sources Uncovers Multiple Resistances from Human Sources. PLoS ONE 2011, 6, e20819. [Google Scholar] [CrossRef] [Green Version]
- El-Bahar, H.M.; Ali, N.G.; Aboyadak, I.M.; Khalil, S.A.E.S.; Ibrahim, M.S. Virulence Genes Contributing to Aeromonas hydrophila Pathogenicity in Oreochromis Niloticus. Int. Microbiol. 2019, 22, 479–490. [Google Scholar] [CrossRef]
- Azzam-Sayuti, M.; Ina-Salwany, M.Y.; Zamri-Saad, M.; Annas, S.; Yusof, M.T.; Monir, M.S.; Mohamad, A.; Muhamad-Sofie, M.H.N.; Lee, J.Y.; Chin, Y.K.; et al. Comparative Pathogenicity of Aeromonas spp. in Cultured Red Hybrid Tilapia (Oreochromis niloticus × O. mossambicus). Biology 2021, 10, 1192. [Google Scholar] [CrossRef]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas Species Infections and Their Significance in Public Health. Sci. World J. 2012, 2012, 625023. [Google Scholar] [CrossRef] [Green Version]
- Jagoda, S.S.; Wijewardana, T.G.; Arulkanthan, A.; Igarashi, Y.; Tan, E.; Kinoshita, S.; Watabe, S.; Asakawa, S. Characterization and antimicrobial susceptibility of motile aeromonads isolated from freshwater ornamental fish showing signs of septicaemia. Dis. Aquat. Org. 2014, 109, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Monir, M.S.; Bagum, N.; Kabir, S.L.; Borty, S.C.; Doulah, M.A. Isolation, Molecular Identification and Characterization of Aeromonas hydrophila from Infected Air-Breathing Catfish Magur (Clarias Batrachus) Cultured in Mymensingh, Bangladesh. Asian Australas. J. Food Saf. Secur. 2017, 1, 17–24. [Google Scholar] [CrossRef]
- Abdel-moneam, D.A.; Ibrahim, R.A.; Nashaat, M.; Shaalan, M. Multifactorial Causes of Mass Mortality in Oreochromis niloticus in Kafr El-Sheikh, Egypt, 198. Bull. Eur. Ass. Fish Pathol. 2021, 41, 2021. [Google Scholar]
- Ling, S.H.M.; Wang, X.H.; Lim, T.M.; Leung, K.Y. Green Fluorescent Protein -Tagged Edwardsiella tarda Reveals Portal of Entry in Fish. FEMS Microbiol. Lett. 2001, 194, 239–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsheshtawy, A.; Yehia, N.; Elkemary, M.; Soliman, H. Investigation of Nile tilapia Summer Mortality in Kafr El-Sheikh Governorate, Egypt. Genet. Aquat. Org. 2019, 3, 17–25. [Google Scholar] [CrossRef]
- Abbott, S.L.; Cheung, W.K.W.; Janda, J.M. The Genus Aeromonas: Biochemical Characteristics, Atypical Reactions, and Phenotypic Identification Schemes. J. Clin. Microbiol. 2003, 41, 2348–2357. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Murcia, A.J.; Benlloch, S.; Collins, M.D. Phylogenetic interrelationships of members of the genera Aeromonas and Plesiomonas as determined by 16S ribosomal DNA sequencing: Lack of congruence with results of DNA-DNA hybridizations. Int J. Syst. Bacteriol. 1992, 42, 412–421. [Google Scholar] [CrossRef] [Green Version]
- Smyrli, M.; Prapas, A.; Rigos, G.; Kokkari, C.; Pavlidis, M.; Katharios, P. Aeromonas veronii infection associated with high morbidity and mortality in farmed European seabass Dicentrarchus labrax in the Aegean Sea, Greece. Fish Pathol. 2017, 52, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Algammal, A.M.; Mabrok, M.; Ezzat, M.; Alfifi, K.J.; Aboelkheir, M.; Esawy, A.M.; Elmasry, N.; El-Tarabili, R.M. Prevalence, antimicrobial resistance (AMR) pattern, virulence determinant and AMR genes of emerging multi-drug resistant Edwardsiella tarda in Nile tilapia and African catfish. Aquaculture 2022, 548, 737643. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, X.R.; Li, J.; Li, G.Y.; Liu, Z.P.; Mo, Z.L. Identification and Virulence Properties of Aeromonas veronii Bv. Sobria Isolates Causing an Ulcerative Syndrome of Loach Misgurnus Anguillicaudatus. J. Fish Dis. 2016, 39, 777–781. [Google Scholar]
- Yogananth, N.; Bhakyaraj, R.; Chanthuru, A.; Anbalagan, T.; Nila, K.M. Detection of Virulence Gene in Aeromonas hydrophila Isolated from Fish Samples Using PCR Technique. Glob. J. Biotechnol. Biochem. 2009, 4, 51–53. [Google Scholar]
- Sha, J.; Kozlova, E.V.; Chopra, A. Role of Various Enterotoxins in Aeromonas hydrophila-Induced Gastroenteritis: Generation of Enterotoxin Gene-Deficient Mutants and Evaluation of Their Enterotoxic Activity. Infect. Immun. 2002, 70, 1924–1935. [Google Scholar] [CrossRef] [Green Version]
- Beaz-Hidalgo, R.; Figueras, M.J. Aeromonas spp. whole genomes and virulence factors implicated in fish disease. J. Fish Dis. 2013, 36, 371–388. [Google Scholar] [CrossRef]
- Namba, A.; Mano, N.; Takano, H.; Beppu, T.; Ueda, K.; Hirose, H. OmpA Is an Adhesion Factor of Aeromonas veronii, an Optimistic Pathogen That Habituates in Carp Intestinal Tract. J. Appl. Microbiol. 2008, 105, 1441–1451. [Google Scholar] [CrossRef]
- Piotrowska, M.; Popowska, M. The Prevalence of Antibiotic Resistance Genes among Aeromonas Species in Aquatic Environments. Ann. Microbiol. 2014, 64, 921–934. [Google Scholar] [CrossRef]
- Elgendy, M.Y.; Shaalan, M.; Abdelsalam, M.; Eissa, A.E.; El-Adawy, M.M.; Seida, A.A. Antibacterial activity of silver nanoparticles against antibiotic-resistant Aeromonas veronii infections in Nile tilapia, Oreochromis niloticus (L.), in vitro and in vivo assay. Aquac. Res. 2022, 53, 901–920. [Google Scholar] [CrossRef]
- Ali, S.E.; Mahana, O.; Mohan, C.V.; Delamare-Deboutteville, J.; Elgendy, M.Y. Genetic characterization and antimicrobial profiling of bacterial isolates collected from Nile tilapia (Oreochromis niloticus) affected by summer mortality syndrome. J. Fish Dis. 2022, 00, 1–15. [Google Scholar] [CrossRef]
- Tekedar, H.C.; Arick, M.A.; Hsu, C.Y.; Thrash, A.; Blom, J.; Lawrence, M.L.; Abdelhamed, H. Identification of Antimicrobial Resistance Determinants in Aeromonas veronii Strain MS-17-88 Recovered from Channel Catfish (Ictalurus Punctatus). Front. Cell. Infect. Microbiol. 2020, 10, 348. [Google Scholar] [CrossRef]
- Fauzi, F.; Hamdan, R.H.; Mohamed, M.; Tan, L.P. Detection of Integron and Antibiotic Resistance Genes of Aeromonas Spp. Isolated from Freshwater Fish in Pahang, Malaysia. Charting Sustain. In Charting the Sustainable Future of ASEAN in Science and Technology; Springer: Berlin/Heidelberg, Germany, 2020; pp. 473–486. [Google Scholar] [CrossRef]
- Chen, F.; Sun, J.; Han, Z.; Yang, X.; Xian, J.A.A.; Lv, A.; Hu, X.; Shi, H. Isolation, Identification and Characteristics of Aeromonas veronii from Diseased Crucian Carp (Carassius Auratus Gibelio). Front. Microbiol. 2019, 10, 2742. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Cycling Conditions (35 Cycles) | Amplified Product | Reference | ||
|---|---|---|---|---|---|---|
| Denatur. | Annealing | Extension | ||||
| 16SrRNA Aeromonas species | F: CTACTTTTGCCGGCGAGCGG | 94 °C for 30 s | 50 °C for 40 s | 72 °C for 50 s | 953 bp | [30] |
| R: TGATTCCCGAAGGCACTCCC | ||||||
| aerA (Aerolysin) | F: CCTATGGCCTGAGCGAGAAG | 94 °C for 30 s | 55.5 °C for 30 s | 72 °C for 30 s | 431 bp | [36] |
| R: CCAGTTCCAGTCCCACCACT | ||||||
| ser (Serine protease) | F: ACGGAGTGCGTTCTTCCTACTCCAG | 94 °C for 1 min | 64 °C for 30 s | 72 °C for 45 s | 211 bp | [37] |
| R: CCGTTCATCACACCGTTGTAGTCG | ||||||
| nuc (Nuclease) | F: CAGGATCTGAACCGCCTCTATCAGG | 94 °C for 1 min | 64 °C for 30 s | 72 °C for 45 s | 504 | |
| R: GTCCCAAGCTTCGAACAGTTTACGC | ||||||
| act Cytotoxic enterotoxin | F: GAGAAGGTGACCACCAAGAACA | 94 °C for 4 min | 59 °C for 30 s | 72 °C for 1 min | 232 bp | [18] |
| R: AACTGACATCGGCCTTGAACTC | ||||||
| alt (Heat-labile cytotonic enterotoxin) | F: TGACCCAGTCCTGGCACGGC | 94 °C for 4 min | 59 °C for 30 s | 72 °C for 1 min | 442 bp | [38] |
| R: GGTGATCGATCACCACCAGC | ||||||
| ahp (Serine protease) | F: ATTGGATCCCTGCCTA | 94 °C for 4 min | 59 °C for 30 s | 72 °C for 1 min | 911 bp | |
| R: GCTAAGCTTGCATCCG | ||||||
| ompAII (Outer membrane protein II) | F:GCTGAATTCATGAAACTCAAAATGGCTC | 94 °C for 1 min | 55 °C for 1 min | 72 °C for 1 min | 1001 | [39] |
| R: GCGAAGCTTTTACTGTTGTACTTGC | ||||||
| blaTEM (Penicillin-resistance) | F: ATCAGCAATAAACCAGC | 94 °C 30 s | 54 °C 40 s | 72 °C 45 s | 516 | [40] |
| R: CCCCGAAGAACGTTTTC | ||||||
| blaSHV (Penicillin-resistance) | F: AGGATTGACTGCCTTTTTG | 94 °C 30 s | 54 °C 40 s | 72 °C 40 s | 392 | |
| R: ATTTGCTGATTTCGCTCG | ||||||
| blaCTX-M (Cephalosporines-resistance) | F: ATGTGCAGYACCAGTAARGTKATGGC | 94 °C 30 s | 54 °C 40 s | 72 °C 45 s | 593 | [41] |
| R: TGGGTRAARTARGTSACCAGAAYC AGC GG | ||||||
| aadA1 (Aminoglycosides-resistance) | F: TATCAGAGGTAGTTGGCGTCAT | 94 °C 30 s | 54 °C 40 s | 72 °C 45 s | 484 | [42] |
| R: GTTCCTAGCGTTAAGGTTTCATT | ||||||
| tetA (Tetracycline-resistance) | F: GGTTCACTCGAACGACGTCA | 94 °C 30 s | 50 °C 40 s | 72 °C 45 s | 576 | |
| R: CTGTCCGACAAGTTGCATGA | ||||||
| sul1 (sulfonamide-resistance) | F: CGGCGTGGGCTACCTGAACG | 94 °C 30 s | 54 °C 40 s | 72 °C 45 s | 433 | [43] |
| R: GCCGATCGCGTGAAGTTCCG | ||||||
| Organ | No. of Positive Isolates | Percentage of Positive Isolates | Chi Square p Value |
|---|---|---|---|
| Liver | 18 | 38.3% | 0.80851 0.6675 NS * |
| Kidneys | 16 | 34.1% | |
| Gills | 13 | 27.6% | |
| Total | 47 | 100% |
| Antimicrobial Class | Antimicrobial Agents | Sensitive | Intermediate | Resistant | |||
|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | ||
| Penicillin | Ampicillin | - | - | - | - | 47 | 100 |
| Piperacillin/ Tazobactam | 13 | 27.7 | - | - | 34 | 72.3 | |
| β-Lactam-β-lactamase-inhibitor combination | Amoxicillin/clavulanic acid | 10 | 21.2 | - | - | 37 | 78.8 |
| Cephalosporin | Cefotaxime | 9 | 19.1 | - | - | 38 | 80.9 |
| Ceftriaxone | 6 | 12.7 | 3 | 6.4 | 38 | 80.9 | |
| Polymyxin | Polymyxin B | 7 | 14.9 | - | - | 40 | 85.1 |
| Aminoglycosides | Streptomycin | 14 | 29.8 | - | - | 33 | 70.2 |
| Tetracycline | Tetracycline | - | - | 2 | 4.3 | 45 | 95.7 |
| Phenicols | Chloramphenicol | 41 | 87.3 | - | - | 6 | 12.7 |
| Ansamycin | Rifamycin SV | - | - | - | - | 47 | 100 |
| Macrolides | Erythromycin | - | - | 11 | 23.4 | 36 | 76.5 |
| Fluroquinolones | Ciprofloxacin | 47 | 100 | - | - | - | - |
| Sulfonamides | Sulfamethoxazole/Trimethoprim | - | - | - | - | 47 | 100 |
| Chi square p value | 252.82 <0.0001 | 92.875 <0.001 | 76.817 <0.0001 | ||||
| Type of Genes | Genes | No of Positive Isolates | % | Chi-Square p-Value |
|---|---|---|---|---|
| Virulence genes | aerA | 47 | 100 | 30.891 <0.0001 |
| alt | 39 | 82.9 | ||
| ser | 29 | 61.7 | ||
| ompAII | 26 | 55.3 | ||
| act | 21 | 44.7 | ||
| ahp | 17 | 36.17 | ||
| nuc | 14 | 29.8 | ||
| Antimicrobial resistance genes | blaTEM | 47 | 100 | 5.1475 0.3981 NS * |
| blaCTX-M | 38 | 80.9 | ||
| blaSHV | 34 | 72.3 | ||
| sul1 | 47 | 100 | ||
| tetA | 45 | 95.7 | ||
| aadA1 | 33 | 70.2 |
| No. of Isolates | % | Type of Resistance | Phenotypic Resistance | Resistance Genes | MAR |
|---|---|---|---|---|---|
| 14 | 29.8% | XDR | Nine classes:
| blaTEM,blaCTX-M, blaSHV,tetA, aadA1, and sul1 | 0.84 |
| 9 | 19.1% | MDR | Eight classes:
| blaTEM, blaCTX-M, blaSHV, tetA, aadA1, and sul1 | 0.76 |
| 7 | 14.9% | MDR | Seven classes:
| blaTEM,blaSHV, tetA, and sul1 | 0.61 |
| 6 | 12.8% | XDR | Nine classes:
| blaTEM, blaCTX-M,aadA1, tetA, and sul1 | 0.76 |
| 5 | 10.6% | MDR | Seven classes:
| blaTEM, blaCTX-M, tetA, and sul1 | 0.61 |
| 4 | 8.5% | MDR | Eight classes:
| blaTEM blaCTX-M,blaSHV,aadA1, tetA, and sul1 | 0.76 |
| 2 | 4.2% | MDR | Four classes:
| blaTEM and sul1 | 0.31 |
| A. veronii Strains | Virulence Genes | Corresponding Group | Cumulative Mortality % |
|---|---|---|---|
| Strain 5 | aerA, alt, ahp, act, ser, nuc, and ompAII | G5 | 100 |
| Strain 4 | aerA, ompAII, alt, ahp, and act | G4 | 90 |
| Strain 3 | aerA, ahp, ser, and nuc | G3 | 85 |
| Strain 2 | aerA and ser | G2 | 75 |
| Strain 1 | aerA | G1 | 55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Algammal, A.M.; Ibrahim, R.A.; Alfifi, K.J.; Ghabban, H.; Alghamdi, S.; Kabrah, A.; Khafagy, A.R.; Abou-Elela, G.M.; Abu-Elala, N.M.; Donadu, M.G.; et al. A First Report of Molecular Typing, Virulence Traits, and Phenotypic and Genotypic Resistance Patterns of Newly Emerging XDR and MDR Aeromonas veronii in Mugil seheli. Pathogens 2022, 11, 1262. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11111262
Algammal AM, Ibrahim RA, Alfifi KJ, Ghabban H, Alghamdi S, Kabrah A, Khafagy AR, Abou-Elela GM, Abu-Elala NM, Donadu MG, et al. A First Report of Molecular Typing, Virulence Traits, and Phenotypic and Genotypic Resistance Patterns of Newly Emerging XDR and MDR Aeromonas veronii in Mugil seheli. Pathogens. 2022; 11(11):1262. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11111262
Chicago/Turabian StyleAlgammal, Abdelazeem M., Reham A. Ibrahim, Khyreyah J. Alfifi, Hanaa Ghabban, Saad Alghamdi, Ahmed Kabrah, Ahmed R. Khafagy, Gehan M. Abou-Elela, Nermeen M. Abu-Elala, Matthew Gavino Donadu, and et al. 2022. "A First Report of Molecular Typing, Virulence Traits, and Phenotypic and Genotypic Resistance Patterns of Newly Emerging XDR and MDR Aeromonas veronii in Mugil seheli" Pathogens 11, no. 11: 1262. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11111262