
Phaeohyphomycosis due to Exophiala in Aquarium-Housed Lumpfish (Cyclopterus lumpus): Clinical Diagnosis and Description
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Antemortem Cytology
2.3. Antemortem Microbiology
2.4. Necropsy and Histopathology
2.5. Fungus Identification and Antifungal Drug Susceptibility Testing
2.6. DNA Extraction and PCR Detection of Exophiala from Formalin-Fixed Paraffin-Embedded Tissues
3. Results
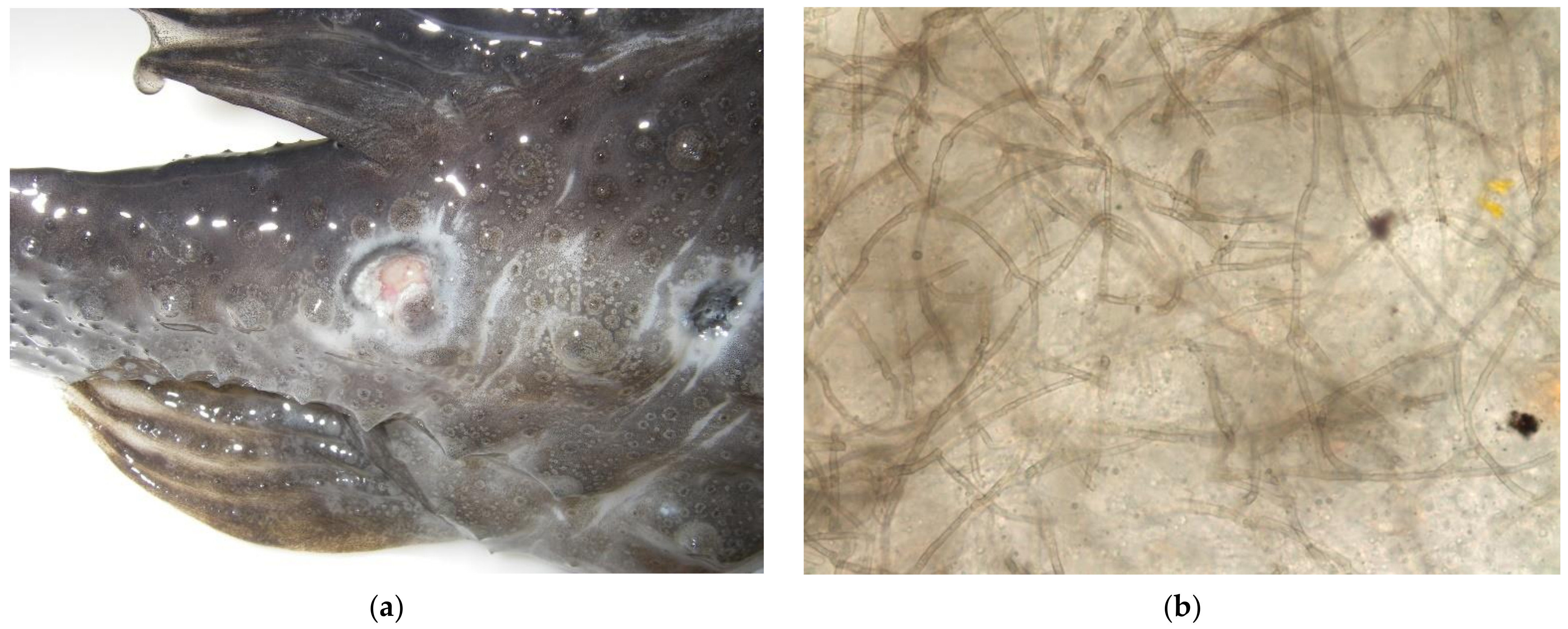
3.1. Observed Clinical Signs
3.2. Antemortem Diagnostics and Treatments
3.2.1. Diagnostics
3.2.2. Attempted Treatments
3.3. Necropsy and Histopathology
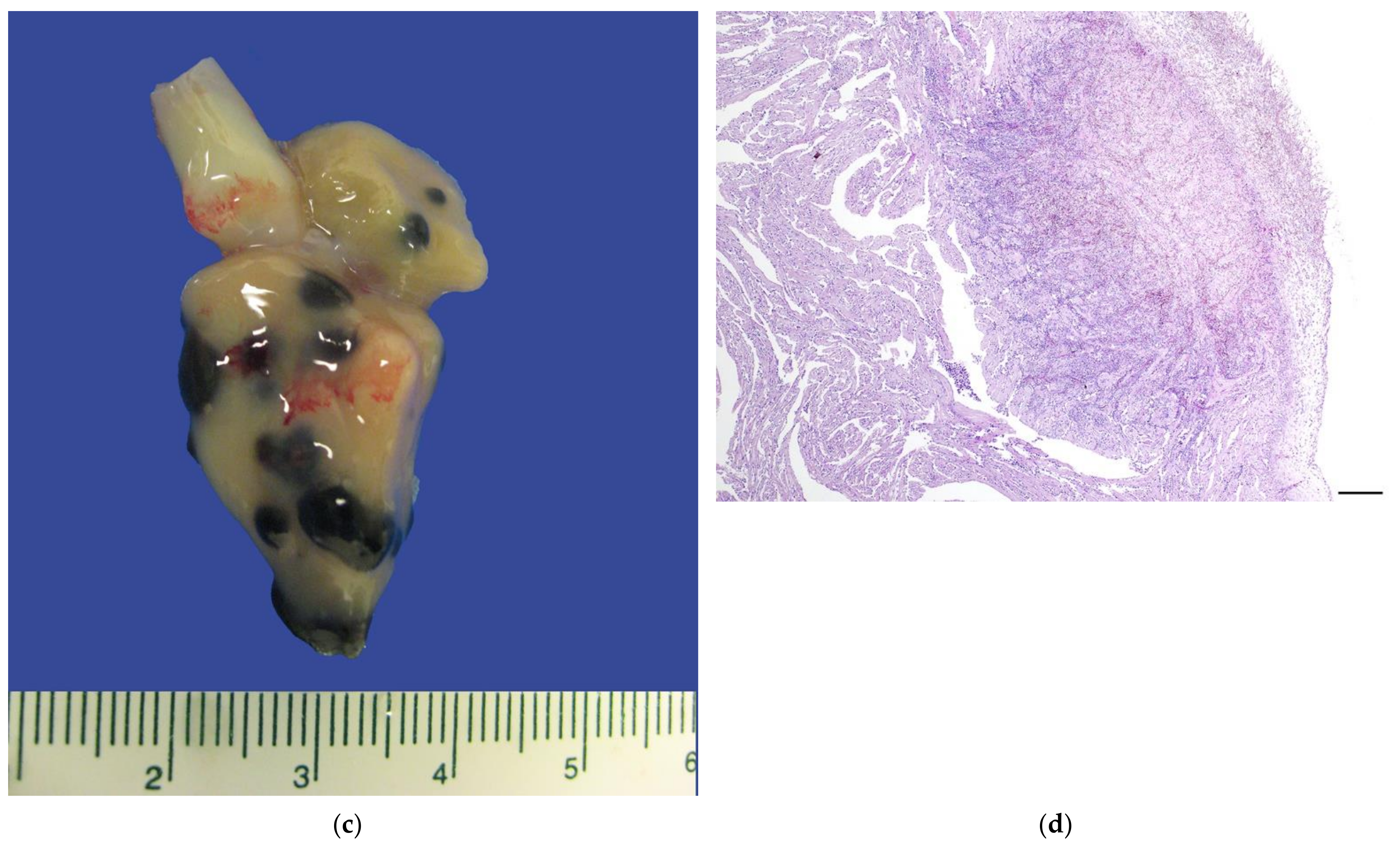
3.3.1. Gross Necropsy
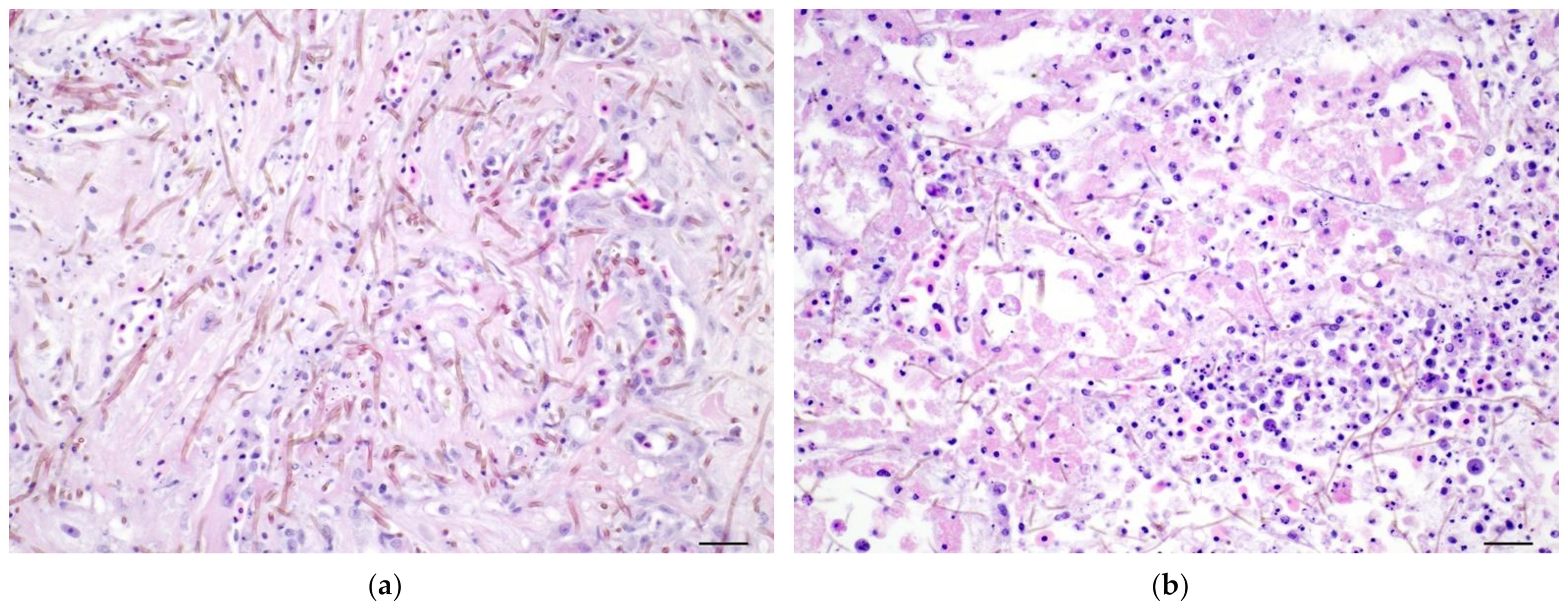
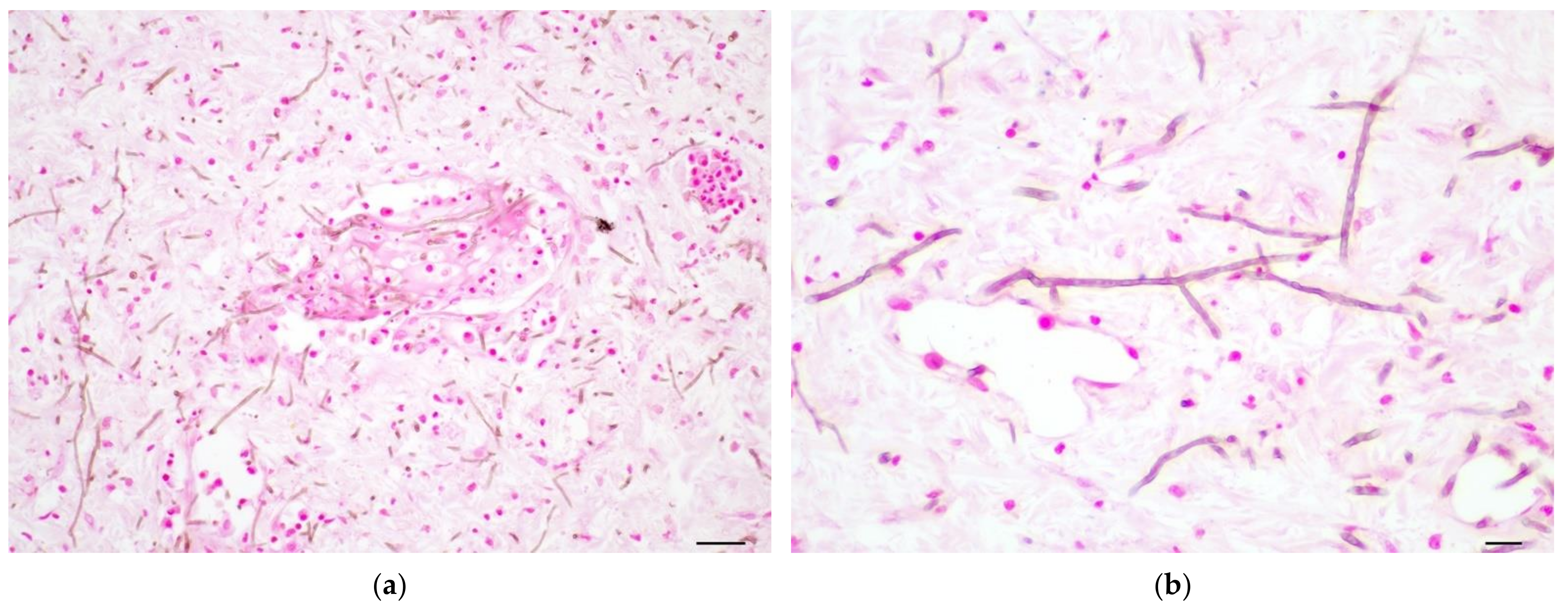
3.3.2. Histopathology
3.4. Fungus Identification and Antifungal Drug Susceptibility Testing
3.5. PCR Detection of Exophiala from Formalin-Fixed Paraffin-Embedded Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
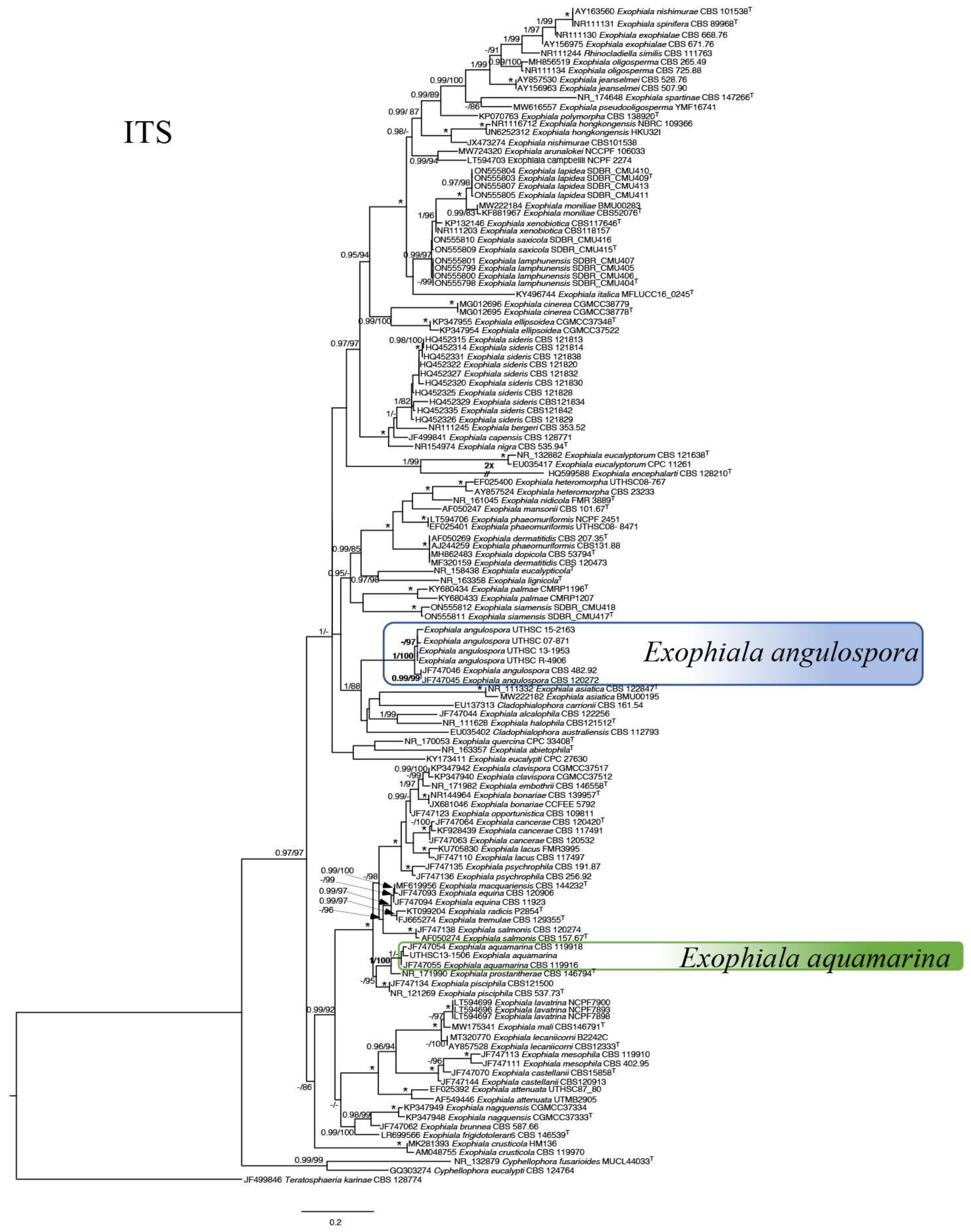{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Target | Amplicon Size (bp) |
|---|---|---|---|
| Exophiala-F1 | ACCYCTTGTTGCTTCGGC | ITS1 | 181 |
| Exophiala-R1 | GCCAGAACCAAGAGATCCGT | ||
| Lumpfish-F1 | AATCATAATCGGGGGCTTTG | COX1 | 245 |
| Lumpfish-R1 | CCGCGAGGTGTAAAGAAAAG |

References
- Eriksen, E.; Durif, C.M.; Prozorkevich, D. Lumpfish (Cyclopterus lumpus) in the Barents Sea: Development of biomass and abundance indices, and spatial distribution. ICES J. Mar. Sci. 2014, 71, 2398–2402. [Google Scholar] [CrossRef] [Green Version]
- Pampoulie, C.; Skirnisdottir, S.; Olafsdottir, G.; Helyar, S.J.; Thorsteinsson, V.; Jónsson, S.Þ.; Fréchet, A.; Durif, C.M.; Sherman, S.; Lampart-Kałużniacka, M. Genetic structure of the lumpfish Cyclopterus lumpus across the North Atlantic. ICES J. Mar. Sci. 2014, 71, 2390–2397. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, S.; Baird, J. The Fishery for Lumpfish [Cyclopterus lumpus] in Newfoundland Waters; Canadian Technical Report of Fisheries and Aquatic Sciences; Fisheries and Oceans Canada: St. John’s, NL, Canada, 1988; pp. IV + 26p. [Google Scholar]
- Kasper, J.; Bertelsen, B.; Ólafsson, H.; Holst, J.; Sturlaugsson, J.; Jónsson, S. Observations of growth and postspawning survival of lumpfish Cyclopterus lumpus from mark-recapture studies. J. Fish Biol. 2014, 84, 1958–1963. [Google Scholar] [CrossRef] [PubMed]
- Hedeholm, R.; Blicher, M.E.; Grønkjær, P. First estimates of age and production of lumpsucker (Cyclopterus lumpus) in Greenland. Fish. Res. 2014, 149, 1–4. [Google Scholar] [CrossRef]
- Powell, A.; Treasurer, J.W.; Pooley, C.L.; Keay, A.J.; Lloyd, R.; Imsland, A.K.; Garcia De Leaniz, C. Use of lumpfish for sea-lice control in salmon farming: Challenges and opportunities. Rev. Aquac. 2018, 10, 683–702. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.; Beckmann, M.J.; Pflaum, S.; Pearson, M.; Carcajona, D.; Treasurer, J.W.; Van West, P. Exophiala angulospora infection in hatchery-reared lumpfish (Cyclopterus lumpus) broodstock. J. Fish Dis. 2019, 42, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Armwood, A.R.; Cañete-Gibas, C.F.; Dill-Okubo, J.A.; Wiederhold, N.P.; Camus, A.C. Retrospective study of phaeohyphomycosis in aquarium-housed fish, with first descriptions of Exophiala lecanii-corni and Neodevriesia cladophorae in fish. J. Fish Dis. 2021, 44, 1563–1577. [Google Scholar] [CrossRef]
- De Hoog, G.S.; Vicente, V.A.; Najafzadeh, M.J.; Harrak, M.J.; Badali, H.; Seyedmousavi, S. Waterborne Exophiala species causing disease in cold-blooded animals. Pers.-Mol. Phylogeny Evol. Fungi 2011, 27, 46–72. [Google Scholar] [CrossRef] [Green Version]
- Nyaoke, A.; Weber, E.S.; Innis, C.; Stremme, D.; Dowd, C.; Hinckley, L.; Gorton, T.; Wickes, B.; Sutton, D.; De Hoog, S.; et al. Disseminated Phaeohyphomycosis in Weedy Seadragons (Phyllopteryx taeniolatus) and Leafy Seadragons (Phycodurus eques) Caused by Species of Exophiala, Including a Novel Species. J. Vet. Diagn. Investig. 2009, 21, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Gaskins, J.E.; Cheung, P.J. Exophiala pisciphila. Mycopathologia 1986, 93, 173–184. [Google Scholar] [CrossRef]
- Otis, E.; Wolke, R.; Blazer, V. Infection of Exophiala salmonis in Atlantic salmon (Salmo salar L.). J. Wildl. Dis. 1985, 21, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.H.; Holliman, A.; Helgason, S. Exophiala salmonis infection in Atlantic salmon Salmo salar L. J. Fish Dis. 1978, 1, 357–368. [Google Scholar] [CrossRef]
- Langdon, J. Cranial Exophiala pisciphila infection in Salmo salar in Australia. Bull. Eur. Assoc. Fish Pathol. 1987, 7, 35–37. [Google Scholar]
- McGinnis, M.R.; Ajello, L. A New Species of Exophiala Isolated From Channel Catfish. Mycologia 1974, 66, 518–520. [Google Scholar] [CrossRef]
- Řehulka, J.; Kolařík, M.; Hubka, V. Disseminated infection due to Exophiala pisciphila in Cardinal tetra, Paracheirodon axelrodi. J. Fish Dis. 2017, 40, 1015–1024. [Google Scholar] [CrossRef]
- Řehulka, J.; Kubátová, A.; Hubka, V. Swim bladder mycosis in pretty tetra (Hemigrammus pulcher) caused by Exophiala pisciphila and Phaeophleospora hymenocallidicola, and experimental verification of pathogenicity. J. Fish Dis. 2018, 41, 487–500. [Google Scholar] [CrossRef]
- Camus, A.; Berliner, A.; Hyatt, M.; Hatcher, N.; Clauss, T. Exophiala xenobiotica aerocystitis in a Queensland grouper Epinephelus lanceolatus (Bloch). J. Fish Dis. 2015, 38, 221–225. [Google Scholar] [CrossRef]
- Munchan, C.; Kurata, O.; Wada, S.; Hatai, K.; Sano, A.; Kamei, K.; Nakaoka, N. Exophiala xenobiotica infection in cultured striped jack, Pseudocaranx dentex (Bloch & Schneider), in Japan. J. Fish Dis. 2009, 32, 893–900. [Google Scholar] [CrossRef]
- Komine, T.; Srivorakul, S.; Tomaru, K.; Tanaka, Y.; Inohana, M.; Fukano, H.; Kurata, O.; Wada, S. Outbreak of Nontuberculous Mycobacteriosis with Dematiaceous Fungus Co-infection in Aquarium-reared Sharphead Flyingfish (Hirundichthys oxycephalus). Fish Pathol. 2021, 56, 187–198. [Google Scholar] [CrossRef]
- Gjessing, M.; Davey, M.; Kvellestad, A.; Vrålstad, T. Exophiala angulospora causes systemic inflammation in Atlantic cod Gadus morhua. Dis. Aquat. Org. 2011, 96, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Kanchan, C.; Muraosa, Y.; Hatai, K. Exophiala angulospora infection found in cultured Japanese flounder Paralichthys olivaceus in Japan. Bull. Eur. Assoc. Fish Pathol. 2014, 34, 187–194. [Google Scholar]
- Overy, D.P.; Groman, D.; Giles, J.; Duffy, S.; Rommens, M.; Johnson, G. Exophiala angulospora Causes Systemic Mycosis in Atlantic Halibut: A Case Report. J. Aquat. Anim. Health 2015, 27, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Madan, V.; Bisset, D.; Harris, P.; Howard, S.; Beck, M.H. Phaeohyphomycosis caused by Exophiala salmonis. Br. J. Dermatol. 2006, 155, 1082–1084. [Google Scholar] [CrossRef] [PubMed]
- Sughayer, M.; DeGirolami, P.C.; Khettry, U.; Korzeniowski, D.; Grumney, A.; Pasarell, L.; McGinnis, M.R. Human infection caused by Exophiala pisciphila: Case report and review. Rev. Infect. Dis. 1991, 13, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.S.; Sutton, D.A.; Fothergill, A.W.; Rinaldi, M.G.; Harrak, M.J.; Hoog, G.S.d. Spectrum of Clinically Relevant Exophiala Species in the United States. J. Clin. Microbiol. 2007, 45, 3713–3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, R.; Hutchinson, W.; Ham, J.; Davis, S. Exophiala sp. infection in captured King George whiting (Sillaginodes punctata). Bull. Eur. Assoc. Fish Pathol. 2003, 23, 128–134. [Google Scholar]
- Najafzadeh, M.J.; Saradeghi Keisari, M.; Vicente, V.A.; Feng, P.; Shamsian, S.A.A.; Rezaei-Matehkolaei, A.; de Hoog, G.S.; Curfs-Breuker, I.; Meis, J.F. In Vitro Activities of Eight Antifungal Drugs against 106 Waterborne and Cutaneous Exophiala Species. Antimicrob. Agents Chemother. 2013, 57, 6395–6398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyedmousavi, S.; Guillot, J.; de Hoog, G.S. Phaeohyphomycoses, Emerging Opportunistic Diseases in Animals. Clin. Microbiol. Rev. 2013, 26, 19–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Food and Drug Administration (USDA). Approved Aquaculture Drugs. Available online: https://www.fda.gov/animal-veterinary/aquaculture/approved-aquaculture-drugs (accessed on 8 November 2022).
- Johnson, K.; Freeman, M.; Laxdal, B.; Kristmundsson, Á. Aetiology and histopathology of a systemic phaeohyphomycosis in farmed lumpfish, Cyclopterus lumpus. Bull. Eur. Assoc. Fish Pathol. 2018, 38, 187–194. [Google Scholar]
- Scholz, F.; Ruane, N.; Marcos-Lopez, M.; Mitchell, S.; Bolton-Warberg, M.; O’Connor, I.; Rodger, H. Systemic mycoses in lumpfish (Cyclopterus lumpus L.) in Ireland: Aetiology and clinical presentation. Bull. Eur. Assoc. Fish Pathol. 2018, 38, 202–212. [Google Scholar]
- Heuch, P.; Øines, Ø.; Knutsen, J.; Schram, T. Infection of wild fishes by the parasitic copepod Caligus elongatus on the south east coast of Norway. Dis. Aquat. Org. 2007, 77, 149–158. [Google Scholar] [CrossRef]
- Alarcón, M.; Thoen, E.; Poppe, T.T.; Bornø, G.; Mohammad, S.N.; Hansen, H. Co-infection of Nucleospora cyclopteri (Microsporidia) and Kudoa islandica (Myxozoa) in farmed lumpfish, Cyclopterus lumpus L., in Norway: A case report. J. Fish Dis. 2016, 39, 411–418. [Google Scholar] [CrossRef]
- Cavin, J.M.; Donahoe, S.L.; Frasca, S.; Innis, C.J.; Kinsel, M.J.; Kurobe, T.; Naples, L.M.; Nyaoke, A.; Poll, C.P.; Weber, E.P.S. Myxobolus albi infection in cartilage of captive lumpfish (Cyclopterus lumpus). J. Vet. Diagn. Investig. 2012, 24, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Freeman, M.A.; Kasper, J.M.; Kristmundsson, Á. Nucleospora cyclopteri n. sp., an intranuclear microsporidian infecting wild lumpfish, Cyclopterus lumpus L., in Icelandic waters. Parasites Vectors 2013, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Mullins, J.; Powell, M.; Speare, D.J.; Cawthorn, R. An intranuclear microsporidian in lumpfish Cyclopterus lumpus. Dis. Aquat. Org. 1994, 20, 7–13. [Google Scholar] [CrossRef]
- Scholz, F.; Fringuelli, E.; Bolton-Warberg, M.; Marcos-López, M.; Mitchell, S.; Prodhol, P.; Moffet, D.; Savage, P.; Murphy O’Sullivan, S.; O′Connor, I.; et al. First record of Tetramicra brevifilum in lumpfish (Cyclopterus lumpus, L.). J. Fish Dis. 2017, 40, 757–771. [Google Scholar] [CrossRef]
- Einarsdottir, T.; Sigurdardottir, H.; Bjornsdottir, T.S.; Einarsdottir, E. Moritella viscosa in lumpfish (Cyclopterus lumpus) and Atlantic salmon (Salmo salar). J. Fish Dis. 2018, 41, 1751–1758. [Google Scholar] [CrossRef]
- Grove, S.; Wiik-Nielsen, C.; Lunder, T.; Tunsjø, H.; Tandstad, N.; Reitan, L.; Marthinussen, A.; Sørgaard, M.; Olsen, A.; Colquhoun, D. Previously unrecognised division within Moritella viscosa isolated from fish farmed in the North Atlantic. Dis. Aquat. Org. 2010, 93, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Marcos-López, M.; Ruane, N.M.; Scholz, F.; Bolton-Warberg, M.; Mitchell, S.O.; Murphy O’Sullivan, S.; Irwin Moore, A.; Rodger, H.D. Piscirickettsia salmonis infection in cultured lumpfish (Cyclopterus lumpus L.). J. Fish Dis. 2017, 40, 1625–1634. [Google Scholar] [CrossRef]
- Rønneseth, A.; Haugland, G.T.; Colquhoun, D.J.; Brudal, E.; Wergeland, H.I. Protection and antibody reactivity following vaccination of lumpfish (Cyclopterus lumpus L.) against atypical Aeromonas salmonicida. Fish Shellfish Immunol. 2017, 64, 383–391. [Google Scholar] [CrossRef]
- Rothacker, T.; Jaffey, J.A.; Rogers, E.R.; Fales, W.H.; Gibas, C.F.C.; Wiederhold, N.P.; Sanders, C.; Mele, J.; Fan, H.; Cohn, L.A.; et al. Novel Penicillium species causing disseminated disease in a Labrador Retriever dog. Med. Mycol. 2020, 58, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, G.; de Hoog, G.S.; Haase, G. Barcode identifiers as a practical tool for reliable species assignment of medically important black yeast species. J. Clin. Microbiol. 2012, 50, 3023–3030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- CLSI Standard M38; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi. CLSI: Wayne, PA, USA, 2017.
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Ito, T. Systemic Exophiala sp. Infection and Diagnostic Efficacy of Ultrasonography in Lumpfish (Cyclopterus lumpus). In Proceedings of the IAAAM, Gold Coast, Australia, 17–21 May 2014. [Google Scholar]
- Madrid, H.; Hernández-Restrepo, M.; Gené, J.; Cano, J.; Guarro, J.; Silva, V. New and interesting chaetothyrialean fungi from Spain. Mycol. Prog. 2016, 15, 1179–1201. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, M.W. Everything Old is New Again: Successful Treatment of Cutaneous Exophiala Phaeohyphomycosis in Two Valentini Pufferfish (Canthigaster valentini) Using Mohs’ Paste IAAAM 2018. In Proceedings of the IAAAM, Long Beach, CA, USA, 19–23 May 2018. [Google Scholar]
- Kenney, R.T.; Kwon-Chung, K.; Waytes, A.T.; Melnick, D.A.; Pass, H.I.; Merino, M.J.; Gallin, J.I. Successful treatment of systemic Exophiala dermatitidis infection in a patient with chronic granulomatous disease. Clin. Infect. Dis. 1992, 14, 235–242. [Google Scholar] [CrossRef]
- Sekiguchi, R.; Urabe, N.; Sakamoto, S.; Sasaki, M.; Homma, S.; Kishi, K. Exophiala dermatitidis pneumonia with bronchiectasis required prolonged voriconazole treatment. Respirol. Case Rep. 2021, 9, e00783. [Google Scholar] [CrossRef]
- Revankar, S.G.; Sutton, D.A. Melanized Fungi in Human Disease. Clin. Microbiol. Rev. 2010, 23, 884–928. [Google Scholar] [CrossRef] [Green Version]
- Gold, W.L.; Vellend, H.; Salit, I.E.; Campbell, I.; Summerbell, R.; Rinaldi, M.; Simor, A.E. Successful treatment of systemic and local infections due to Exophiala species. Clin. Infect. Dis. 1994, 19, 339–341. [Google Scholar] [CrossRef]




| Institution | House ID | Cutaneous Lesions Present? | Direct Cytology Performed? | Fungal Elements on Skin Scrape | Biopsy | Antemortem Fungal culture | Culture Source | Culture Result | Isolate Identification by FTL on Antemortem Culture | Isolate Identification on Postmortem FFPE | MIC Data Available |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NEAq | 3152 | Y | Y | Y | N | Y | Caudal tail lesion, Swab | Pigmented mold growth | Exophiala angulospora | None | Y |
| N | Y | Dorsal skin lesion, Swab | Pigmented mold growth | Exophiala angulospora | None | Y | |||||
| NEAq | 3189 | Y | Y | Y | N | Y | Skin lesion, Swab | Pigmented mold growth | Exophiala angulospora ** | ND | Y |
| NEAq | 3197 | Y | Y | N | N | N | Closest match to Cyphellophora guyanensis | ||||
| Quebec | 09N032 | N | N | N/A | N | N | ND | ||||
| Quebec | 09N038 | N | N | N/A | N | N | ND | ||||
| Quebec | 09N051 | Y | N | N/A | N | N | ND | ||||
| NEAq | 6192 | Y | Y | Y | Y (not used for culture) | Y | Skin lesion, Swab | No growth | Closest match to Cyphellophora guyanensis | ||
| NEAq | 6193 | Y | Y | Y | N | Y | Aspirate from Skin Cyst, Swab | No growth | ND | ||
| N | Y | Skin lesion, Swab | Pigmented mold growth | N/A | N | ||||||
| Quebec | 11N210 | Y | Y | Y | N | Y | Skin Lesion, Swab | No growth | ND | ||
| Aq of the Bay | 11028 | Y | Y | Y | N | N | N/A | ||||
| WHSA | 1103 | Y | Y | Y | N | N | None | ||||
| Quebec | 12N091 | N | N | N/A | N | N | ND | ||||
| NADC | T12069 | Y | Y | Y | Y | Y | Skin Lesion, Swab | No growth | ND | ||
| Y | Skin lesion, Biopsy | Pigmented mold growth | Exophiala aquamarina | Identical to Exophiala aquamarina | Y | ||||||
| NADC | T12068 | Y | Y | Y | Y | Y | Skin lesion, Swab | No growth, bacterial contamination | ND | ||
| Y | Skin lesion, Biopsy | Pigmented mold growth | Exophiala angulospora | Identical to Exophiala aquamarina | N | ||||||
| NEAq | 9404 | Y | Y | Y | Y | Y | Skin lesion, Biopsy | Pigmented mold growth | Exophiala angulospora | ND | N |
| Institution | House ID | Integument/Muscle | Gill | Kidney (Cranial and/or Caudal) | Liver | Heart | Spleen | Intestine | Coelomic Cavity/Serosa | Gonad | Stomach | Eye | Mesentery |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NEAq | 3152 | X | X | X | X | X | X | ||||||
| NEAq | 3189 | X | |||||||||||
| NEAq | 3197 | X | X | X | X | X | X | X | |||||
| Quebec | 09N032 | X | X | X | |||||||||
| Quebec | 09N038 | X | X | X | X | X | |||||||
| Quebec | 09N051 | X | X | X | X | ||||||||
| NEAq | 6192 | X | X | X | X | X | X | ||||||
| NEAq | 6193 | X | X | ||||||||||
| Quebec | 11N210 | X | |||||||||||
| Aq of the Bay | 11028 | X | X | X | |||||||||
| WHSA | 1103 | X | |||||||||||
| Quebec | 12N091 | X | X | X | X | X | X | X | X | ||||
| NADC | T12069 | X | |||||||||||
| NADC | T12068 | X | X | X | X | ||||||||
| NEAq | 9404 | X | X | X | X | X | X | X | X | X | |||
| Total: | 11 | 9 | 9 | 7 | 6 | 6 | 4 | 3 | 2 | 2 | 1 | 1 |
| Institution | House ID | Species | Location | AMB | FLC | ITC | VRC | TRB | MON |
|---|---|---|---|---|---|---|---|---|---|
| NEAq | 3152 | E. angulospora | Caudal tail | 0.25 | 32–64 | 0.125 | 0.5 | 2 | ~ |
| E. angulospora | Dorsum | 0.25 | 32 | 0.06 | 0.5 | 2 | ~ | ||
| NEAq | 3189 | E. angulospora | Skin lesion | 2 | 32–64 | 0.25 | 0.5 | 2 | ~ |
| NADC | T12069 | E. aquamarina | Skin lesion | ~ | ~ | 0.5 | ~ | 0.03 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McDermott, C.T.; Innis, C.J.; Nyaoke, A.C.; Tuxbury, K.A.; Cavin, J.M.; Weber, E.S.; Edmunds, D.; Lair, S.; Spangenberg, J.V.; Hancock-Ronemus, A.L.; et al. Phaeohyphomycosis due to Exophiala in Aquarium-Housed Lumpfish (Cyclopterus lumpus): Clinical Diagnosis and Description. Pathogens 2022, 11, 1401. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121401
McDermott CT, Innis CJ, Nyaoke AC, Tuxbury KA, Cavin JM, Weber ES, Edmunds D, Lair S, Spangenberg JV, Hancock-Ronemus AL, et al. Phaeohyphomycosis due to Exophiala in Aquarium-Housed Lumpfish (Cyclopterus lumpus): Clinical Diagnosis and Description. Pathogens. 2022; 11(12):1401. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121401
Chicago/Turabian StyleMcDermott, Colin T., Charles J. Innis, Akinyi C. Nyaoke, Kathryn A. Tuxbury, Julie M. Cavin, E. Scott Weber, Deana Edmunds, Stéphane Lair, Jill V. Spangenberg, Amy L. Hancock-Ronemus, and et al. 2022. "Phaeohyphomycosis due to Exophiala in Aquarium-Housed Lumpfish (Cyclopterus lumpus): Clinical Diagnosis and Description" Pathogens 11, no. 12: 1401. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121401