
The Tapeworm Hymenolepis diminuta as an Important Model Organism in the Experimental Parasitology of the 21st Century


{kind=link}
{kind=link}
{kind=link}
Abstract
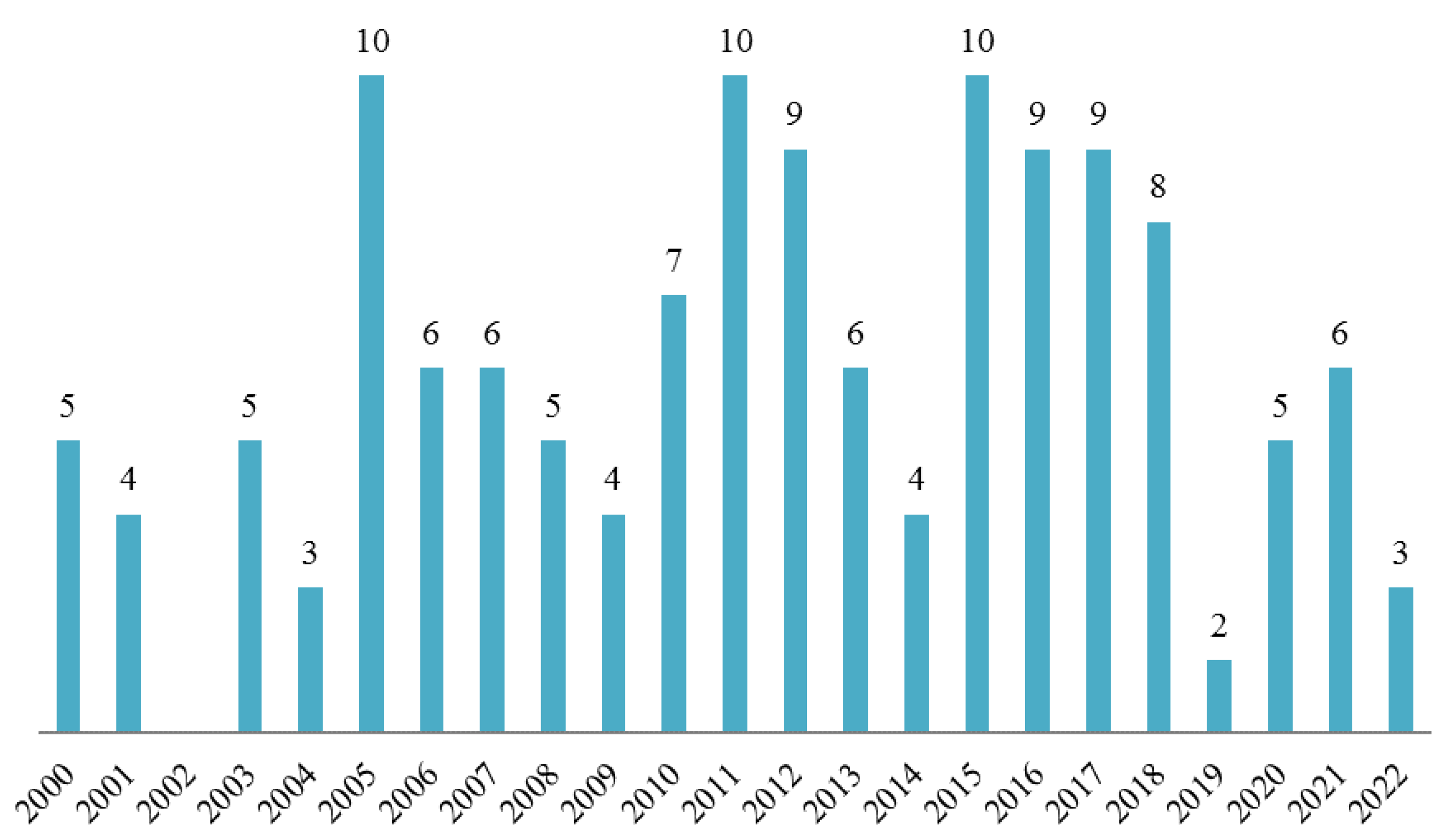
:1. Introduction
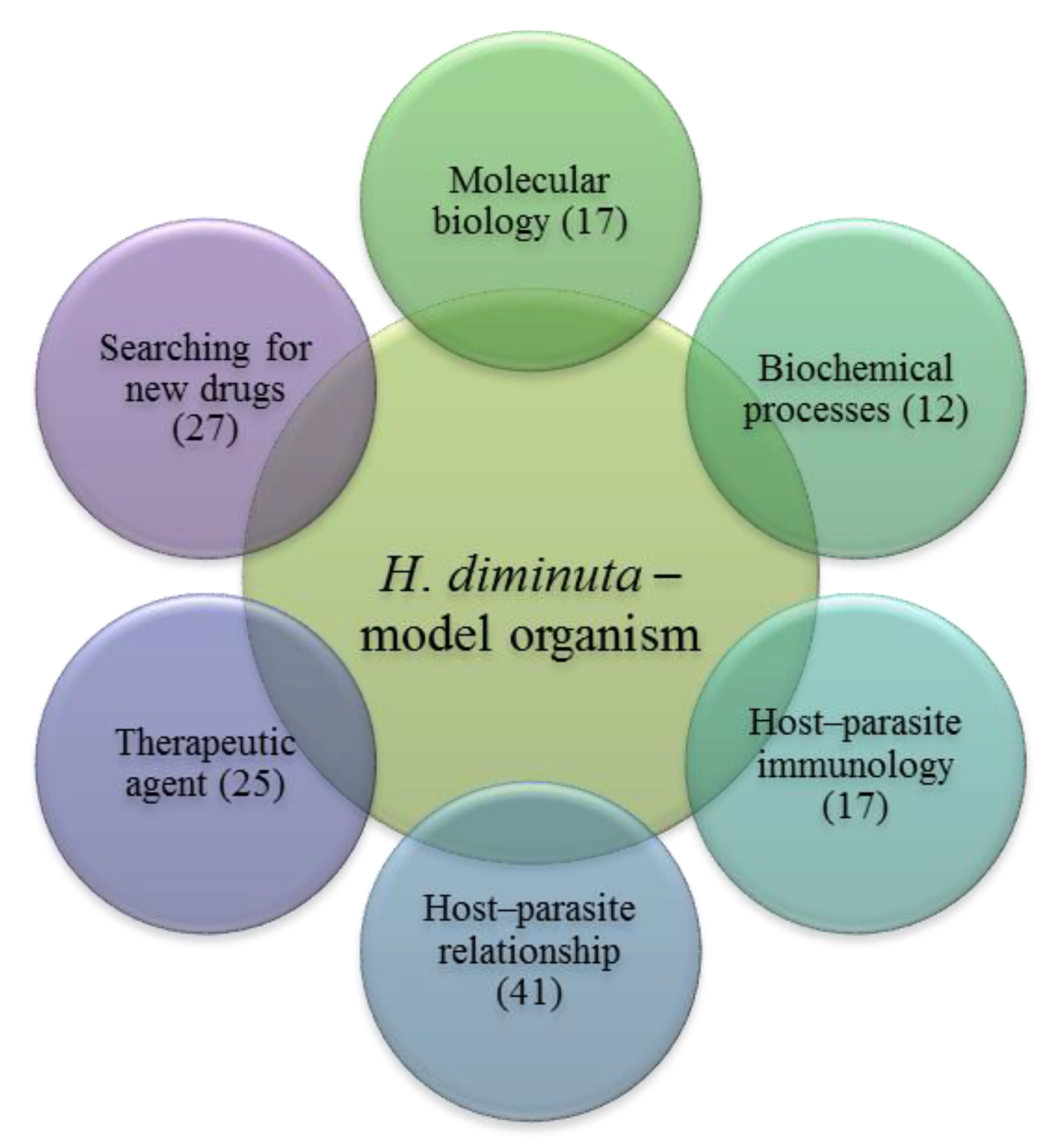
2. Molecular Biology
3. Biochemistry
4. The Host–Parasite Immunology
5. H. diminuta as a Therapeutic Agent
6. The Host–Parasite Relationship
7. Treatments
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunter, P. The paradox of model organisms. The use of model organisms in research will continue despite their shortcomings. EMBO Rep. 2008, 9, 717–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuitton, D.A.; Gottstein, B. Echinococcus multilocularis and its intermediate host: A model of parasite-host interplay. J. Biomed. Biotechnol. 2010, 2010, 923193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, I.J.; Zarowiecki, M.; Holroyd, N.; Garciarrubio, A.; Sanchez-Flores, A.; Brooks, K.L.; Tracey, A.; Bobes, R.J.; Fragoso, G.; Sciutto, E.; et al. The genomes of four tapeworm species reveal adaptations to parasitism. Nature 2013, 496, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, P. Hox genes and the parasitic flatworms: New opportunities, challenges and lessons from the free-living. Parasitol. Int. 2008, 57, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Markoski, M.M.; Bizarro, C.; Farias, S.; Espinoza, I.; Galanti, N.; Zaha, A.; Ferreira, H. In Vitro Segmentation Induction of Mesocestoides Corti (Cestoda) Tetrathyridia. J. Parasitol. 2003, 89, 27–34. [Google Scholar] [CrossRef] [PubMed]
- de Lima, J.C.; Monteiro, K.M.; Cabrera, T.N.B.; Paludo, G.P.; Moura, H.; Barr, J.R.; Zaha, A.; Ferreira, H.B. Comparative proteomics of the larval and adult stages of the model cestode parasite Mesocestoides corti. J. Proteom. 2018, 175, 127–135. [Google Scholar] [CrossRef]
- McKAY, D.M. The immune response to and immunomodulation by Hymenolepis diminuta. Parasitology 2009, 137, 385–394. [Google Scholar] [CrossRef]
- Rozario, T.; Newmark, P.A. A confocal microscopy-based atlas of tissue architecture in the tapeworm Hymenolepis diminuta. Exp. Parasitol. 2015, 158, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Woolsey, I.D.; Fredensborg, B.L.; Jensen, P.M.; Kapel, C.M.O.; Meyling, N.V. An insect–tapeworm model as a proxy for anthelminthic effects in the mammalian host. Parasitol. Res. 2015, 114, 2777–2780. [Google Scholar] [CrossRef] [Green Version]
- Mansur, F.; Luoga, W.; Buttle, D.; Duce, I.; Lowe, A.; Behnke, J. The anthelmintic efficacy of natural plant cysteine proteinases against two rodent cestodes Hymenolepis diminuta and Hymenolepis microstoma in vitro. Vet. Parasitol. 2014, 201, 48–58. [Google Scholar] [CrossRef]
- Saghir, N.; Conde, P.J.; Brophy, P.M.; Barrett, J. A new diagnostic tool for neurocysticercosis is a member of a cestode specific hydrophobic ligand binding protein family. FEBS Lett. 2000, 487, 181–184. [Google Scholar] [CrossRef] [PubMed]
- von Nickisch-Rosenegk, M.; Brown, W.M.; Boore, J.L. Complete Sequence of the Mitochondrial Genome of the Tapeworm Hymenolepis diminuta: Gene Arrangements Indicate that Platyhelminths Are Eutrochozoans. Mol. Biol. Evol. 2001, 18, 721–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, R.M.; Jastrzębski, J.P.; Kuśmirek, W.; Sałamatin, R.; Rydzanicz, M.; Sobczyk-Kopcioł, A.; Sulima-Celińska, A.; Paukszto, Ł.; Makowczenko, K.G.; Płoski, R.; et al. Hybrid de novo whole-genome assembly and annotation of the model tapeworm Hymenolepis diminuta. Sci. Data 2020, 6, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohajer-Maghari, B.; Amini-Bavil-Olyaee, S.; Webb, R.A.; Coe, I.R. Molecular cloning and characterization of Hymenolepis diminuta alpha-tubulin gene. DNA Seq. 2007, 18, 80–83. [Google Scholar] [CrossRef]
- Řežábková, L.; Brabec, J.; Jirků, M.; Dellerba, M.; Kuchta, R.; Modrý, D.; Parker, W.; Pomajbíková, K.J. Genetic diversity of the potentially therapeutic tapeworm Hymenolepis diminuta (Cestoda: Cyclophyllidea). Parasitol. Int. 2019, 71, 121–125. [Google Scholar] [CrossRef]
- Sharma, S.; Lyngdoh, D.; Roy, B.; Tandon, V. Differential diagnosis and molecular characterization of Hymenolepis nana and Hymenolepis diminuta (Cestoda: Cyclophyllidea: Hymenolepididae) based on nuclear rDNA ITS2 gene marker. Parasitol. Res. 2016, 115, 4293–4298. [Google Scholar] [CrossRef]
- Hoque, T.; Boghal, M.; Webb, R.A. Validation of internal controls for gene expression analysis in the intestine of rats infected with Hymenolepis diminuta. Parasitol. Int. 2007, 56, 325–329. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.I.; Wojtkowiak-Giera, A.; Kolasa, A.; Baranowska-Bosiacka, I.; Lanocha, N.; Wandurska-Nowak, E.; Izabela, G.; Salamatin, R.; Jagodzinski, P.P. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes and protein (TLR3 and TLR9) in the small and large intestines of rats. Exp. Parasitol. 2014, 145, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Salamatin, R.; Jagodzinski, P.; Wandurska-Nowak, E. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes (TLR2 and TLR4) in the small and large intestines of rats. Exp. Parasitol. 2012, 130, 261–266. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.; Wojtkowiak-Giera, A.; Kolasa, A.; Czernomysy-Furowicz, D.; Lanocha-Arendarczyk, N.; Wandurska-Nowak, E.; Salamatin, R.; Jagodzinski, P. Hymenolepis diminuta: Analysis of the expression of Toll-like receptor genes (TLR2 and TLR4) in the small and large intestines of rats. Part II. Exp. Parasitol. 2013, 135, 437–445. [Google Scholar] [CrossRef]
- Kosik-Bogacka, D.I.; Baranowska-Bosiacka, I.; Kolasa-Wołosiuk, A.; Lanocha-Arendarczyk, N.; Gutowska, I.; Korbecki, J.; Namięta, H.; Rotter, I. The inflammatory effect of infection with Hymenolepis diminuta via the increased expression and activity of COX-1 and COX-2 in the rat jejunum and colon. Exp. Parasitol. 2016, 169, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Wang, M.-H.; Pai, A.; Yan, G. Transcription profiling of immune genes during parasite infection in susceptible and resistant strains of the flour beetles (Tribolium castaneum). Exp. Parasitol. 2013, 134, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warr, E.; Meredith, J.M.; Nimmo, D.D.; Basu, S.; Hurd, H.; Eggleston, P. A tapeworm molecule manipulates vitellogenin expression in the beetle Tenebrio molitor. Insect Mol. Biol. 2006, 15, 497–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikopoulos, G.J.; Hoque, T.; Webb, R.A. Infection with the cestode Hymenolepis diminuta induces changes in acetylcholine metabolism and muscarinic receptor mRNA expression in the rat jejunum. Parasitol. Res. 2006, 99, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Pai, A.; Yan, G. Quantitative Trait Loci for Susceptibility to Tapeworm Infection in the Red Flour Beetle. Genetics 2003, 165, 1307–1315. [Google Scholar] [CrossRef]
- Zhong, D.; Pai, A.; Yan, G. Costly resistance to parasitism: Evidence from simultaneous quantitative trait loci mapping for resistance and fitness in Tribolium castaneum. Genetics 2005, 169, 2127–2135. [Google Scholar] [CrossRef] [Green Version]
- Warr, E.; Eggleston, P.; Hurd, H. Apoptosis in the fat body tissue of the beetle Tenebrio molitor parasitised by Hymenolepis diminuta. J. Insect Physiol. 2004, 50, 1037–1043. [Google Scholar] [CrossRef]
- Saghir, N.; Conde, P.J.; Brophy, P.M.; Barrett, J. Biochemical characterisation of a hydrophobic ligand binding protein from the tapeworm Hymenolepis diminuta. Int. J. Parasitol. 2001, 31, 653–660. [Google Scholar] [CrossRef]
- Onufriev, M.V.; Gulyaeva, N.V.; Terenina, N.B.; Tolstenkov, O.O.; Gustafsson, M.K.S. The effect of a nitric oxide donor on the synthesis of cGMP in Hymenolepis diminuta: A radiometric study. Parasitol. Res. 2004, 95, 22–24. [Google Scholar] [CrossRef]
- Park, J.P.; Fioravanti, C.F. Catalysis of NADH→NADP+ transhydrogenation by adult Hymenolepis diminuta mitochondria. Parasitol. Res. 2005, 98, 200–206. [Google Scholar] [CrossRef]
- Mercer-Haines, N.; Fioravanti, C.F. Hymenolepis diminuta: Mitochondrial transhydrogenase as an additional site for anaerobic phosphorylation. Exp. Parasitol. 2008, 119, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.-C.; Lai, S.-C. Induction of cysteine proteinase in the encapsulation of Hymenolepis diminuta eggs in the American cockroach, Periplaneta americana. J. Invertebr. Pathol. 2006, 92, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Skrzycki, M.; Majewska, M.; Podsiad, M.; Czeczot, H.; Salamatin, R.; Twarowska, J.; Grytner-Zięcina, B. Hymenolepis diminuta: Experimental studies on the antioxidant system with short and long term infection periods in the rats. Exp. Parasitol. 2011, 129, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Czeczot, H.; Skrzycki, M.; Majewska-Wierzbicka, M.; Podsiad, M.; Salamatin, R.; Grytner-Zięcina, B. The antioxidant defence mechanisms of parasite and host after chronic Hymenolepis diminuta infestation of the rat. Pol. J. Vet. Sci. 2013, 16, 121–123. [Google Scholar] [CrossRef] [Green Version]
- Kosik-Bogacka, D.I.; Baranowska-Bosiacka, I.; Noceń, I.; Jakubowska, K.; Chlubek, D. Hymenolepis diminuta: Activity of anti-oxidant enzymes in different parts of rat gastrointestinal tract. Exp. Parasitol. 2011, 128, 265–271. [Google Scholar] [CrossRef]
- Vokřál, I.; Jirásko, R.; Jedličková, V.; Bártíková, H.; Skálová, L.; Lamka, J.; Holčapek, M.; Szotáková, B. The inability of tapeworm Hymenolepis diminuta and fluke Dicrocoelium dendriticum to metabolize praziquantel. Vet. Parasitol. 2012, 185, 168–174. [Google Scholar] [CrossRef]
- Bártíková, H.; Vokřál, I.; Skálová, L.; Kubíček, V.; Firbasová, J.; Briestenský, D.; Lamka, J.; Szotáková, B. The activity of drug-metabolizing enzymes and the biotransformation of selected anthelmintics in the model tapeworm Hymenolepis diminuta. Parasitology 2012, 139, 809–818. [Google Scholar] [CrossRef]
- Kreshchenko, N.; Terenina, N.; Ermakov, A. Serotonin Signalling in Flatworms: An Immunocytochemical Localisation of 5-HT7 Type of Serotonin Receptors in Opisthorchis felineus and Hymenolepis diminuta. Biomolecules 2021, 11, 1212. [Google Scholar] [CrossRef]
- Wegener Parfrey, L.; Jirků, M.; Šíma, R.; Jalovecká, M.; Sak, B.; Grigore, K.; Jirků Pomajbíková, K. A benign helminth alters the host immune system and the gut microbiota in a rat model system. PLoS ONE 2017, 12, e0182205. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.; Maizels, R.M. Diversity and dialogue in immunity to helminths. Nat. Rev. Immunol. 2011, 11, 375–388. [Google Scholar] [CrossRef]
- Webb, R.A.; Hoque, T.; Dimas, S. Expulsion of the gastrointestinal cestode, Hymenolepis diminuta by tolerant rats: Evidence for mediation by a Th2 type immune enhanced goblet cell hyperplasia, increased mucin production and secretion. Parasite Immunol. 2007, 29, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; McKAY, D.M. Immune modulation by a high molecular weight fraction from the rat tapeworm Hymenolepis diminuta. Parasitology 1999, 130, 575–585. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.M.; Khan, W.I. STAT-6 Is an Absolute Requirement for Murine Rejection of Hymenolepis diminuta. J. Parasitol. 2003, 89, 188–189. [Google Scholar] [CrossRef] [PubMed]
- Ishih, A.; Uchikawa, R. Immunoglobulin E and mast cell responses are related to worm biomass but not expulsion of Hymenolepis diminuta during low dose infection in rats. Parasite Immunol. 2000, 22, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T.; Kai, T.; Miyasaka, Y.; Maruyama, H.; Ishih, A.; Kino, H. Intestinal immunity suppresses carrying capacity of rats for the model tapeworm, Hymenolepis diminuta. Parasitol. Int. 2018, 67, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.-L.R.; Leung, G.; McKay, D.M. Cestode regulation of inflammation and inflammatory diseases. Int. J. Parasitol. 2013, 43, 233–243. [Google Scholar] [CrossRef]
- Lopes, F.; Reyes, J.L.; Wang, A.; Leung, G.; McKay, D.M. Enteric epithelial cells support growth of Hymenolepis diminuta in vitro and trigger TH2-promoting events in a species-specific manner. Int. J. Parasitol. 2015, 45, 691–696. [Google Scholar] [CrossRef]
- Starke-Buzetti, W.A.; Oaks, J.A. Increased glial-derived neurotrophic factor in the small intestine of rats infected with the tapeworm, Hymenolepis diminuta. Int. J. Exp. Pathol. 2008, 89, 458–465. [Google Scholar] [CrossRef]
- Bień, J.; Sałamatin, R.; Sulima, A.; Savijoki, K.; Bruce Conn, D.; Näreaho, A.; Młocicki, D. Mass spectrometry analysis of the excretory-secretory (E-S) products of the model cestode Hymenolepis diminuta reveals their immunogenic properties and the presence of new E-S proteins in cestodes. Acta Parasitol. 2016, 61, 429–442. [Google Scholar] [CrossRef]
- Zawistowska-Deniziak, A.; Basałaj, K.; Strojny, B.; Młocicki, D. New Data on Human Macrophages Polarization by Hymenolepis diminuta Tapeworm—An In Vitro Study. Front. Immunol. 2017, 8, 148. [Google Scholar] [CrossRef]
- Johnston, M.J.; Wang, A.; Catarino, M.E.; Ball, L.; Phan, V.C.; MacDonald, J.A.; McKay, D.M. Extracts of the rat tapeworm, Hymenolepis diminuta, suppress macrophage activation in vitro and alleviate chemically induced colitis in mice. Infect. Immun. 2010, 78, 1364–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aira, N.; Andersson, A.-M.; Singh, S.K.; McKay, D.M.; Blomgran, R. Species dependent impact of helminth-derived antigens on human macrophages infected with Mycobacterium tuberculosis: Direct effect on the innate anti-mycobacterial response. PLoS Negl. Trop. Dis. 2017, 11, e0005390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, N.; Venu, V.K.P.; Yipp, B.; Petri, B.; Hirota, S.; Gilleard, J.; McKay, D.M.; Lopes, F. A Trypsin-Sensitive Proteoglycan from the Tapeworm Hymenolepis diminuta Inhibits Murine Neutrophil Chemotaxis in vitro by Suppressing p38 MAP Kinase Activation. J. Innate Immun. 2018, 11, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.M.; Wang, A.; Hirota, C.L.; McKay, D.M. Neutralizing Anti-IL-10 Antibody Blocks the Protective Effect of Tapeworm Infection in a Murine Model of Chemically Induced Colitis. J. Immunol. 2005, 174, 7368–7375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, M.M.; Wang, A.; Parhar, K.K.S.; Johnston, M.J.; Van Rooijen, N.; Beck, P.L.; McKay, D.M. In Vitro-Derived Alternatively Activated Macrophages Reduce Colonic Inflammation in Mice. Gastroenterology 2010, 138, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Melon, A.; Wang, A.; Phan, V.; McKay, D.M. Infection with Hymenolepis diminuta Is More Effective than Daily Corticosteroids in Blocking Chemically Induced Colitis in Mice. J. Biomed. Biotechnol. 2009, 2010, 384523. [Google Scholar] [CrossRef] [Green Version]
- Persaud, R.; Wang, A.; Reardon, C.; McKay, D.M. Characterization of the immuno-regulatory response to the tapeworm Hymenolepis diminuta in the non-permissive mouse host. Int. J. Parasitol. 2007, 37, 393–403. [Google Scholar] [CrossRef]
- Matisz, C.E.; Leung, G.; Reyes, J.L.; Wang, A.; Sharkey, K.A.; McKay, D.M. Adoptive transfer of helminth antigen-pulsed dendritic cells protects against the development of experimental colitis in mice. Eur. J. Immunol. 2015, 45, 3126–3139. [Google Scholar] [CrossRef] [Green Version]
- Matisz, C.E.; Faz-López, B.; Thomson, E.; Al Rajabi, A.; Lopes, F.; Terrazas, L.I.; Wang, A.; Sharkey, K.A.; McKay, D.M. Suppression of colitis by adoptive transfer of helminth antigen-treated dendritic cells requires interleukin-4 receptor-α signaling. Sci. Rep. 2017, 7, 40631. [Google Scholar] [CrossRef]
- Matisz, C.E.; Geuking, M.B.; Lopes, F.; Petri, B.; Wang, A.; Sharkey, K.A.; McKay, D.M. Helminth Antigen-Conditioned Dendritic Cells Generate Anti-Inflammatory CD4 T Cells Independent of Antigen Presentation via MHC II. Am. J. Pathol. 2018, 188, 2589–2604. [Google Scholar] [CrossRef]
- Reyes, J.L.; Fernando, M.R.; Lopes, F.; Leung, G.; Mancini, N.L.; Matisz, C.E.; Wang, A.; McKay, D.M. IL-22 Restrains Tapeworm-Mediated Protection against Experimental Colitis via Regulation of IL-25 Expression. PLoS Pathog. 2016, 12, e1005481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Rajeev, S.; Wang, A.; McKay, D.M. Infection with Hymenolepis diminuta Blocks Colitis and Hastens Recovery While Colitis Has Minimal Impact on Expulsion of the Cestode from the Mouse Host. Pathogens 2021, 10, 994. [Google Scholar] [CrossRef] [PubMed]
- Pomajbíková, K.J.; Jirků, M.; Levá, J.; Sobotková, K.; Morien, E.; Parfrey, L.W. The benign helminth Hymenolepis diminuta ameliorates chemically induced colitis in a rat model system. Parasitology 2018, 145, 1324–1335. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Arai, T.; Campbell, A.; Reyes, J.L.; Lopes, F.; McKay, D.M. Triggering immunological memory against the tapeworm Hymenolepis diminuta to protect against colitis. Parasite Immunol. 2017, 39, e12490. [Google Scholar] [CrossRef]
- Reyes, J.L.; Wang, A.; Fernando, M.R.; Graepel, R.; Leung, G.; van Rooijen, N.; Sigvardsson, M.; McKay, D.M. Splenic B Cells from Hymenolepis diminuta—Infected Mice Ameliorate Colitis Independent of T Cells and via Cooperation with Macrophages. J. Immunol. 2014, 194, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Reardon, C.; Sanchez, A.; Hogaboam, C.M.; McKay, D.M. Tapeworm Infection Reduces Epithelial Ion Transport Abnormalities in Murine Dextran Sulfate Sodium-Induced Colitis. Infect. Immun. 2001, 69, 4417–4423. [Google Scholar] [CrossRef] [Green Version]
- Reyes, J.L.; Lopes, F.; Leung, G.; Mancini, N.L.; Matisz, C.E.; Wang, A.; Thomson, E.A.; Graves, N.; Gilleard, J.; McKay, D.M. Treatment with Cestode Parasite Antigens Results in Recruitment of CCR2+ Myeloid Cells, the Adoptive Transfer of Which Ameliorates Colitis. Infect. Immun. 2016, 84, 3471–3483. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Wang, A.; Prescott, D.; Waterhouse, C.C.; Zhang, S.; McDougall, J.J.; Sharkey, K.A.; McKay, D.M. Infection with an intestinal helminth parasite reduces Freund’s complete adjuvant-induced monoarthritis in mice. Arthritis Rheum. 2011, 63, 434–444. [Google Scholar] [CrossRef]
- Graepel, R.; Leung, G.; Wang, A.; Villemaire, M.; Jirik, F.R.; Sharkey, K.A.; McDougall, J.J.; McKay, D.M. Murine autoimmune arthritis is exaggerated by infection with the rat tapeworm, Hymenolepis diminuta. Int. J. Parasitol. 2013, 43, 593–601. [Google Scholar] [CrossRef]
- McKay, D.M.; Wallace, J.L. Acetic Acid Induced Ulceration in Rats Is Not Affected by Infection with Hymenolepis diminuta. J. Parasitol. 2009, 95, 481–482. [Google Scholar] [CrossRef]
- Fan, P.-C.; Chung, W.-C.; Ito, A. Immunization of Rodents Against Hymenolepis Infections using Non-Viable Homologous Oncospheres. Kaohsiung J. Med. Sci. 2004, 20, 575–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, T.; Lopes, F.; Shute, A.; Wang, A.; McKay, D.M. Young mice expel the tapeworm Hymenolepis diminuta and are protected from colitis by triggering a memory response with worm antigen. Am. J. Physiol. Liver Physiol. 2018, 314, G461–G470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, K.; Morton, C.; Mathew, A.; Karuturi, S.; Haley, C.; Zhang, M.; Holzknecht, Z.E.; Swanson, C.; Lin, S.S.; Parker, W. Production and Use of Hymenolepis diminuta Cysticercoids as Anti-Inflammatory Therapeutics. J. Clin. Med. 2017, 6, 98. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.M.; Wang, A.; Mckay, D.M. Helminth Infection Enhances Disease in a Murine TH2 Model of Colitis. Gastroenterology 2007, 132, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Fernando, M.; Leung, G.; Phan, V.; Smyth, D.; McKay, D.M. Exacerbation of Oxazolone Colitis by Infection with the Helminth Hymenolepis diminuta: Involvement of IL-5 and Eosinophils. Am. J. Pathol. 2010, 177, 2850–2859. [Google Scholar] [CrossRef]
- Sauer, S.; Beinart, D.; Finn, S.M.B.; Kumar, S.L.; Cheng, Q.; Hwang, S.E.; Parker, W.; Devi, G.R. Hymenolepis diminuta-based helminth therapy in C3(1)-TAg mice does not alter breast tumor onset or progression. Evol. Med. Public Health 2021, 9, 131–138. [Google Scholar] [CrossRef]
- Sloup, V.; Jankovská, I.; Štolcová, M.; Magdálek, J.; Karešová, V.; Lanková, S.; Langrová, I. Effects of excessive dietary zinc or zinc/cadmium and tapeworm infection on the biochemical parameters in rats. J. Anim. Physiol. Anim. Nutr. 2021, 105, 989–995. [Google Scholar] [CrossRef]
- Colinet, H.; Salin, C.; Boivin, G.; Hance, T. Host age and fitness-related traits in a koinobiont aphid parasitoid. Ecol. Èntomol. 2005, 30, 473–479. [Google Scholar] [CrossRef]
- Blaser, M.; Schmid-Hempel, P. Determinants of virulence for the parasite Nosema whitei in its host Tribolium castaneum. J. Invertebr. Pathol. 2005, 89, 251–257. [Google Scholar] [CrossRef]
- Shostak, A.W. Effect of Age of the Intermediate Host Tribolium confusum (Coleoptera) on Infection by Hymenolepis diminuta (Cestoda). J. Parasitol. 2008, 94, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, S.; Buss, S.M.; Cassidy, E.J.; Meyling, N.V.; Fredensborg, B.L. Establishment Success of the Beetle Tapeworm Hymenolepis diminuta Depends on Dose and Host Body Condition. Insects 2018, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitchen, S.J.; Shostak, A.W.; Belosevic, M. Hymenolepis diminuta (Cestoda) induces changes in expression of select genes of Tribolium confusum (Coleoptera). Parasitol. Res. 2009, 105, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Shea, J.F. The effect of Hymenolepis diminuta (Cestoda) cysticercoids on the weight change, frass production, and food intake of the intermediate host, Tenebrio molitor (Coleoptera). Parasitol. Res. 2005, 98, 1. [Google Scholar] [CrossRef] [PubMed]
- Shea, J.F. Sex differences in frass production and weight change in Tenebrio molitor (Coleoptera) infected with cysticercoids of the tapeworm Hymenolepis diminuta (Cestoda). J. Insect Sci. 2005, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Shea, J.F. Lack of preference for infective faeces in Hymenolepis diminuta-infected beetles (Tenebrio molitor). J. Helminthol. 2007, 81, 293–299. [Google Scholar] [CrossRef]
- Ponton, F.; Lalubin, F.; Fromont, C.; Wilson, K.; Behm, C.; Simpson, S.J. Hosts use altered macronutrient intake to circumvent parasite-induced reduction in fecundity. Int. J. Parasitol. 2011, 41, 43–50. [Google Scholar] [CrossRef]
- DeMuth, J.P.; Naidu, A.; Mydlarz, L.D. Sex, War, and Disease: The Role of Parasite Infection on Weapon Development and Mating Success in a Horned Beetle (Gnatocerus cornutus). PLoS ONE 2012, 7, e28690. [Google Scholar] [CrossRef]
- Hurd, H.; Warr, E.; Polwart, A. A parasite that increases host lifespan. Proc. R. Soc. B Boil. Sci. 2001, 268, 1749–1753. [Google Scholar] [CrossRef] [Green Version]
- Hurd, H.; Arme, C. Hymenolepis diminuta: Effect of metacestodes on production and viability of eggs in the intermediate host, Tenebrio molitor. J. Invertebr. Pathol. 1986, 47, 225–230. [Google Scholar] [CrossRef]
- Shostak, A.W. Tapeworm (Hymenolepis diminuta) infection in flour beetles (Tribolium confusum): Does it cause a trade-off between host fecundity and egg size? Can. J. Zool. 2009, 87, 1087–1095. [Google Scholar] [CrossRef]
- Pai, A.; Yan, G. Effects of Tapeworm Infection on Male Reproductive Success and Mating Vigor in The Red Flour Beetle, Tribolium Castaneum. J. Parasitol. 2003, 89, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Hurd, H.; Ardin, R. Infection increases the value of nuptial gifts, and hence male reproductive success, in the Hymenolepis diminuta-Tenebrio molitor association. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270 (Suppl. 2), S172–S174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, T.; Eggleston, P.; Hurd, H. Juvenile hormone titre and egg production in Tenebrio molitor infected by Hymenolepis diminuta: Effect of male and/or female infection, male age and mating. J. Insect Physiol. 2003, 49, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Worden, B.D.; Parker, P.G.; Pappas, P.W. Parasites reduce attractiveness and reproductive success in male grain beetles. Anim. Behav. 2000, 59, 543–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, C.; Poulin, R. Preference of female rats for the odours of non-parasitised males: The smell of good genes? Folia Parasitol. 2000, 47, 6–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worden, B.D.; Parker, P.G. Females prefer noninfected males as mates in the grain beetle Tenebrio molitor: Evidence in pre- and postcopulatory behaviours. Anim. Behav. 2005, 70, 1047–1053. [Google Scholar] [CrossRef]
- Sheiman, I.M.; Shkutin, M.F.; Terenina, N.B.; Gustafsson, M.K.S. A behavioral study of the beetle Tenebrio molitor infected with cysticercoids of the rat tapeworm Hymenolepis diminuta. Naturwissenschaften 2006, 93, 305–308. [Google Scholar] [CrossRef]
- Webster, J.P.; Gowtage-Sequeira, S.; Berdoy, M.; Hurd, H. Predation of beetles (Tenebrio molitor) infected with tapeworms (Hymenolepis diminuta): A note of caution for the Manipulation Hypothesis. Parasitology 2000, 120, 313–318. [Google Scholar] [CrossRef]
- Blecharz-Klin, K.; Świerczyńska, M.; Piechal, A.; Wawer, A.; Joniec-Maciejak, I.; Pyrzanowska, J.; Wojnar, E.; Zawistowska-Deniziak, A.; Sulima-Celińska, A.; Młocicki, D.; et al. Infection with intestinal helminth (Hymenolepis diminuta) impacts exploratory behavior and cognitive processes in rats by changing the central level of neurotransmitters. PLoS Pathog. 2022, 18, e1010330. [Google Scholar] [CrossRef]
- Goswami, R.; Singh, S.M.; Kataria, M.; Somvanshi, R. Clinicopathological studies on spontaneous Hymenolepis diminuta infection in wild and laboratory rats. Braz. J. Vet. Pathol. 2011, 4, 103–111. [Google Scholar]
- Kosik-Bogacka, D.I.; Kolasa, A. Histopathological changes in small and large intestines during hymenolepidosis in rats. Folia Biol. 2012, 60, 195–198. [Google Scholar] [CrossRef]
- Ahmad, A.K.; Abdel-Hafeez, E.H.; Kamal, A.M. Some studies on spontaneous Hymenolepis diminuta infection in laboratory rats. J. Egypt. Soc. Parasitol. 2015, 45, 115–124. [Google Scholar] [PubMed]
- Kosik-Bogacka, D.I.; Baranowska-Bosiacka, I.; Salamatin, R. Hymenolepis diminuta: Effect of infection on ion transport in colon and blood picture of rats. Exp. Parasitol. 2010, 124, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Kosik-Bogacka, D.I.; Kolasa, A.; Baranowska-Bosiacka, I.; Marchlewicz, M. Hymenolepis diminuta: The effects of infection on transepithelial ion transport and tight junctions in rat intestines. Exp. Parasitol. 2011, 127, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, N.P.; Brownfield, M.S.; Devente, J.; Bass, P.; Oaks, J.A. cGMP Secreted From the Tapeworm Hymenolepis diminuta Is a Signal Molecule to the Host Intestine. J. Parasitol. 2008, 94, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Kapczuk, P.; Kosik-Bogacka, D.; Kupnicka, P.; Kopytko, P.; Tarnowski, M.; Kolasa, A.; Chlubek, D.; Baranowska-Bosiacka, I. Hymenolepis diminuta Infection Affects Apoptosis in the Small and Large Intestine. Int. J. Environ. Res. Public Health 2022, 19, 9753. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, T.P.; Rathnayaka, Y.; Perera, P.K.; Peachey, L.E.; Nolan, M.J.; Krause, L.; Rajakaruna, R.S.; Cantacessi, C. Infections by human gastrointestinal helminths are associated with changes in faecal microbiota diversity and composition. PLoS ONE 2017, 12, e0184719. [Google Scholar] [CrossRef]
- Zaiss, M.M.; Harris, N.L. Interactions between the intestinal microbiome and helminth parasites. Parasite Immunol. 2015, 38, 5–11. [Google Scholar] [CrossRef] [Green Version]
- McKenney, E.A.; Williamson, L.; Yoder, A.D.; Rawls, J.F.; Bilbo, S.D.; Parker, W. Alteration of the rat cecal microbiome during colonization with the helminth Hymenolepis diminuta. Gut Microbes 2015, 6, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Shute, A.; Wang, A.; Jayme, T.S.; Strous, M.; McCoy, K.D.; Buret, A.G.; McKay, D.M. Worm expulsion is independent of alterations in composition of the colonic bacteria that occur during experimental Hymenolepis diminuta-infection in mice. Gut Microbes 2020, 11, 497–510. [Google Scholar] [CrossRef]
- Shute, A.; Callejas, B.E.; Li, S.; Wang, A.; Jayme, T.S.; Ohland, C.; Lewis, I.A.; Layden, B.T.; Buret, A.G.; McKay, D.M. Cooperation between host immunity and the gut bacteria is essential for helminth-evoked suppression of colitis. Microbiome 2021, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fredensborg, B.L.; Fossdal, Í.; Kálvalíð, I.; Johannesen, T.B.; Stensvold, C.R.; Nielsen, H.V.; Kapel, C.M.O. Parasites modulate the gut-microbiome in insects: A proof-of-concept study. PLoS ONE 2020, 15, e0227561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulima, A.; Bień, J.; Savijoki, K.; Näreaho, A.; Sałamatin, R.; Conn, D.B.; Młocicki, D. Identification of immunogenic proteins of the cysticercoid of Hymenolepis diminuta. Parasites Vectors 2017, 10, 577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sulima, A.; Savijoki, K.; Bień-Kalinowska, J.; Näreaho, A.; Sałamatin, R.; Conn, D.B.; Młocicki, D. Comparative Proteomic Analysis of Hymenolepis diminuta Cysticercoid and Adult Stages. Front. Microbiol. 2018, 8, 2672. [Google Scholar] [CrossRef] [Green Version]
- Młocicki, D.; Sulima, A.; Bień, J.; Näreaho, A.; Zawistowska-Deniziak, A.; Basałaj, K.; Sałamatin, R.; Conn, D.B.; Savijoki, K. Immunoproteomics and Surfaceomics of the Adult Tapeworm Hymenolepis diminuta. Front. Immunol. 2018, 9, 2487. [Google Scholar] [CrossRef] [Green Version]
- Mazanec, H.; Koník, P.; Gardian, Z.; Kuchta, R. Extracellular vesicles secreted by model tapeworm Hymenolepis diminuta: Biogenesis, ultrastructure and protein composition. Int. J. Parasitol. 2020, 51, 327–332. [Google Scholar] [CrossRef]
- Burt, M.D.B. Aspects of the Life History and Systematics of Hymenolepis Diminuta. In Biology of the Tapeworm Hymenolepis Diminuta; Arai, P.H., Ed.; Academic Press: New York, NY, USA, 1980; pp. 1–57. [Google Scholar]
- Álvarez-Fernández, B.E.; Rodrfguez-Bataz, E.; Dfaz-Chiguer, D.L.; Márquez-Navarro, A.; Sánchez-Manzano, R.M.; Nogueda-Torres, B. Mixed Hymenolepis species infection in two family members: A case report from an urban area of Chilpancingo, Guerrero, México. Trop. Gastroenterol. 2012, 33, 83–84. [Google Scholar] [CrossRef]
- Kołodziej, P.; Rzymowska, J.; Stępień-Rukasz, H.; Lorencowicz, R.; Lucińska, M.; Dzióbek, M. Analysis of a child infected with Hymenolepis diminuta in Poland. Ann. Agric. Environ. Med. 2014, 21, 510–511. [Google Scholar] [CrossRef] [Green Version]
- Sinhabahu, V.; Perera, T.; Samarasinghe, S. A case of Hymenolepis diminuta (rat tape worm) infestation in a child. Ceylon Med. J. 2014, 59, 70–71. [Google Scholar] [CrossRef] [Green Version]
- Panti-May, J.A.; Rodríguez-Vivas, R.I.; García-Prieto, L.; Servián, A.; Costa, F. Worldwide overview of human infections with Hymenolepis diminuta. Parasitol. Res. 2020, 119, 1997–2004. [Google Scholar] [CrossRef]
- Jones, W.E. Niclosamide as a Treatment for Hymenolepis diminuta and Dipylidium Caninum Infection in Man. Am. J. Trop. Med. Hyg. 1979, 28, 300–302. [Google Scholar] [CrossRef]
- Tangpu, V.; Temjenmongla; Yadav, A.K. Anticestodal property of Strobilanthes discolor: An experimental study in Hymenolepis diminuta—Rat model. J. Ethnopharmacol. 2006, 105, 459–463. [Google Scholar] [CrossRef]
- Yadav, A.K.; Tangpu, V. Anticestodal activity of Adhatoda vasica extract against Hymenolepis diminuta infections in rats. J. Ethnopharmacol. 2008, 119, 322–324. [Google Scholar] [CrossRef]
- Yadav, A.K.; Tangpu, V. Therapeutic efficacy of Zanthoxylum rhetsa DC extract against experimental Hymenolepis diminuta (Cestoda) infections in rats. J. Parasit. Dis. 2009, 33, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.K. Temjenmongla Anticestodal activity of Houttuynia cordata leaf extract against Hymenolepis diminuta in experimentally infected rats. J. Parasit. Dis. 2011, 35, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.K.; Tangpu, V. Anthelmintic activity of ripe fruit extract of Solanum myriacanthum Dunal (Solanaceae) against experimentally induced Hymenolepis diminuta (Cestoda) infections in rats. Parasitol. Res. 2011, 110, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Kundu, S.; Roy, S.; Lyndem, L.M. Cassia alata L: Potential role as anthelmintic agent against Hymenolepis diminuta. Parasitol. Res. 2012, 111, 1187–1192. [Google Scholar] [CrossRef]
- Giri, B.R.; Bharti, R.R.; Roy, B. In vivo anthelmintic activity of Carex baccans and its active principle resveratrol against Hymenolepis diminuta. Parasitol. Res. 2015, 114, 785–788. [Google Scholar] [CrossRef]
- Deori, K.; Yadav, A.K. Anthelmintic effects of Oroxylum indicum stem bark extract on juvenile and adult stages of Hymenolepis diminuta (Cestoda), an in vitro and in vivo study. Parasitol. Res. 2015, 115, 1275–1285. [Google Scholar] [CrossRef]
- Gogoi, S.; Yadav, A.K. In vitro and in vivo anthelmintic effects of Caesalpinia bonducella (L.) Roxb. leaf extract on Hymenolepis diminuta (Cestoda) and Syphacia obvelata (Nematoda). J. Intercult. Ethnopharmacol. 2016, 5, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Nath, P.; Yadav, A.K. Anthelmintic activity of a standardized extract from the rhizomes of Acorus calamus Linn. (Acoraceae) against experimentally induced cestodiasis in rats. J. Intercult. Ethnopharmacol. 2016, 5, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Roy, S.; Nandi, S.; Ukil, B.; Lyndem, L.M. In vitro anthelmintic effects of Senna occidentalis (L.) link (Leguminosae) on rat tapeworm Hymenolepis diminuta. Int. J. Pharm. Pharm. Sci. 2015, 7, 268–271. [Google Scholar]
- Kundu, S.; Roy, S.; Nandi, S.; Ukil, B.; Lyndem, L.M. Senna alexandrina Mill. induced ultrastructural changes in Hymenolepis diminuta. J. Parasit. Dis. 2017, 41, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ukil, B.; Roy, S.; Nandi, S.; Lyndem, L.M. Senna Plant Induces Disruption on The Mitochondria of Hymenolepis Diminuta. Int. J. Pharm. Pharm. Sci. 2018, 10, 136–138. [Google Scholar] [CrossRef] [Green Version]
- Ukil, B.; Joardar, N.; Babu, S.P.S.; Lyndem, L.M. Effect of Senna plant on the mitochondrial activity of Hymenolepis diminuta. J. Parasit. Dis. 2021, 46, 139–151. [Google Scholar] [CrossRef]
- Roy, S.; Joardar, N.; Babu, S.P.S.; Lyndem, L.M. Senna plant generates reactive oxygen species (ROS) and induces apoptosis in Hymenolepis diminuta. Mol. Biochem. Parasitol. 2020, 238, 111297. [Google Scholar] [CrossRef]
- Abdel-Ghaffar, F.; Semmler, M.; Al-Rasheid, K.A.S.; Strassen, B.; Fischer, K.; Aksu, G.; Klimpel, S.; Mehlhorn, H. The effects of different plant extracts on intestinal cestodes and on trematodes. Parasitol. Res. 2010, 108, 979–984. [Google Scholar] [CrossRef]
- Sapaat, A.; Satrija, F.; Mahsol, H.H.; Ahmad, A.H. Anthelmintic activity of papaya seeds on Hymenolepis diminuta infections in rats. Trop. Biomed. 2012, 29, 508–512. [Google Scholar]
- Stepek, G.; Buttle, D.J.; Duce, I.R.; Lowe, A.; Behnke, J.M. Assessment of the anthelmintic effect of natural plant cysteine proteinases against the gastrointestinal nematode, Heligmosomoides polygyrus, in vitro. Parasitology 2004, 130, 203–211. [Google Scholar] [CrossRef]
- Dhakal, S.; Meyling, N.V.; Williams, A.R.; Mueller-Harvey, I.; Fryganas, C.; Kapel, C.M.; Fredensborg, B.L. Efficacy of condensed tannins against larval Hymenolepis diminuta (Cestoda) in vitro and in the intermediate host Tenebrio molitor (Coleoptera) in vivo. Vet. Parasitol. 2015, 207, 49–55. [Google Scholar] [CrossRef]
- Merwad, A.; Mitchell, S.; Zajac, A.; Flick, G.; Lindsay, D. Effects of high pressure processing on hatching of eggs of the zoonotic rat tapeworm Hymenolepis diminuta. Vet. Parasitol. 2010, 176, 185–188. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulima-Celińska, A.; Kalinowska, A.; Młocicki, D. The Tapeworm Hymenolepis diminuta as an Important Model Organism in the Experimental Parasitology of the 21st Century. Pathogens 2022, 11, 1439. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121439
Sulima-Celińska A, Kalinowska A, Młocicki D. The Tapeworm Hymenolepis diminuta as an Important Model Organism in the Experimental Parasitology of the 21st Century. Pathogens. 2022; 11(12):1439. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121439
Chicago/Turabian StyleSulima-Celińska, Anna, Alicja Kalinowska, and Daniel Młocicki. 2022. "The Tapeworm Hymenolepis diminuta as an Important Model Organism in the Experimental Parasitology of the 21st Century" Pathogens 11, no. 12: 1439. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11121439