
Determinants of Virus Variation, Evolution, and Host Adaptation


1
Nebraska Center for Virology, University of Nebraska-Lincoln, Lincoln, NE 68583, USA
2
Department of Plant Pathology, University of Nebraska-Lincoln, Lincoln, NE 68583, USA
3
Complex Biosystems Interdisciplinary Life Sciences Program, University of Nebraska-Lincoln, Lincoln, NE 68583, USA
*
Author to whom correspondence should be addressed.
Pathogens 2022, 11(9), 1039; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11091039
Submission received: 8 July 2022
/
Revised: 6 September 2022
/
Accepted: 9 September 2022
/
Published: 13 September 2022
(This article belongs to the Special Issue Evolution of Plant Viruses)
Abstract
:Virus evolution is the change in the genetic structure of a viral population over time and results in the emergence of new viral variants, strains, and species with novel biological properties, including adaptation to new hosts. There are host, vector, environmental, and viral factors that contribute to virus evolution. To achieve or fine tune compatibility and successfully establish infection, viruses adapt to a particular host species or to a group of species. However, some viruses are better able to adapt to diverse hosts, vectors, and environments. Viruses generate genetic diversity through mutation, reassortment, and recombination. Plant viruses are exposed to genetic drift and selection pressures by host and vector factors, and random variants or those with a competitive advantage are fixed in the population and mediate the emergence of new viral strains or species with novel biological properties. This process creates a footprint in the virus genome evident as the preferential accumulation of substitutions, insertions, or deletions in areas of the genome that function as determinants of host adaptation. Here, with respect to plant viruses, we review the current understanding of the sources of variation, the effect of selection, and its role in virus evolution and host adaptation.
1. Introduction
As the global population approaches 9 billion people, current food production is threatened by climate change and decreasing land availability, making food security an increasing concern [1]. One approach to addressing these concerns is to reduce crop loss caused by plant pathogens, including viruses. Plant viruses affect economically important staple, cash, non-cash, and secondary-staple crops, resulting in over USD 30 billion in crop loss annually [2,3]. Further, plant viruses make up almost 50% of the emerging and reemerging infectious diseases [2,4]. There is a rise in virus outbreaks in part because climate change increases the geographic areas where viruses and vectors overlap [5,6,7]. This problem is exacerbated by viruses spreading into new areas through global trade and agricultural expansion [2,8].
Plant viruses are obligate parasites that use host processes and resources to replicate and spread. They consist of single- or double-stranded DNA or RNA contained within a virion. To enter the host, plant viruses need to bypass the cuticle and cell wall [9]. This can take place either through a virus vector feeding on the plant or through mechanical means. Successful infection only occurs when there is a compatible interaction between a host and a virus [6,10]. Insects are the most numerous vectors, with aphids, whiteflies, and leafhoppers being the most common [11]. However, mites, fungi, and nematodes are important vectors for some viruses [11]. Plant viruses can also be spread through grafting, seed, and vegetative cuttings [6]. Once inside the cell, viruses uncoat, and their genetic information, for positive strand RNA viruses is translated by the host machinery, leading to virus replication and virion formation [9]. In a continuous cycle, infection begins at a single-cell level, spreads cell to cell through the plasmodesmata, and eventually leads to long distance movement throughout the host [12].
Plants defend against viruses through multiple mechanisms, including gene silencing [13], autophagy [14], and Resistance (R) genes [15]. R genes interact with pathogen avirulence genes (avr) that activate host defenses, including programmed cell death [15]. Dominant and recessive R genes have been identified in wild relatives and introduced through plant breeding into commercial cultivars of important crops such as tomato, soybean, and potato [16,17,18]. However, new virus variants and species can quickly break host resistance, especially when virus resistance is dependent on a single gene [19,20]. This phenomenon occurs when in key areas of the virus genome, mutations accumulate that provide a selective advantage [19,20,21].
The establishment of infection involves a compatible interaction between a virus and a host in a favorable environment (Figure 1). During replication, viruses may recombine, generate mutations, and, for RNA viruses, exist as a cloud of genetic variants (quasispecies) [22,23,24,25]. There is natural genetic diversity in host plants and vectors and variation in environmental conditions [20,26]. Accordingly, viruses are forced to interact with and adapt to genetically diverse host and vector factors in diverse environments. Thus, host, vector, and environmental factors, through genetic drift and selection, drive virus evolution (Figure 1). Virus evolution is established as changes in the genomic structure of a viral population, causing the emergence of new viral variants or species with novel biological properties.
This review provides a broad view of plant virus evolution and host adaptation through the lenses of genetic variation and selection. It is focused on the contributions of mutation, reassortment, and recombination to the generation of genetic diversity and on selection constraints imposed by vectors, hosts, and environmental factors, as driving forces for the emergence of new variants and new viral species with novel biological properties. Contributions of mutation, reassortment, and recombination to genetic variation and the balancing effects on selection and bottlenecks on reducing genetic variation have been reviewed [27].
1.1. Plant Virus Genome Organization
Plant viruses are a diverse group of species with a broad array of genome organization and gene expression strategies classified into taxonomic families, each with several genera and species. Whether measured by family, genus, or species, the number of RNA viruses is greater than the number of DNA viruses, with most plant viruses being (+) sense ssRNA viruses [28]. Virions might be enveloped or not enveloped, with diverse morphologies, genome sizes, and organization [29]. These characteristics are most similar between viruses in the same taxonomic family or genus but do vary across families. The most common genome structure is a segmented genome (Figure 2). The families Potyviridae, Closteroviridae, Tombusviridae, and Geminiviridae contain both monopartite and bipartite species (Figure 2). Single-stranded genomes and segmented genomes have been linked to a wider host range [30]. Segmented genomes have shown to support higher mutation rates, particle stability, and genetic exchange through reassortment [31]. These genome structures are the most common across plant viruses, suggesting there is an evolutionary benefit.
1.2. Virus-Plant Co-Evolution
Plant viruses rely on host factors to replicate, move cell to cell, and travel systemically, but there is natural variation in those host factors [20,32]. Accordingly, plant viruses co-evolve with their host [33,34] and establish a balance between genomic functionality and the bottlenecks imposed by host genetic diversity, host compatibility, antiviral defense, and the fitness cost to the virus associated with host–range expansion or host specialization [10,30,35,36]. The coevolution of viruses with their hosts is tightly linked to an arm race with the host defense system and the availability of host factors for pro-viral functions in multilayered cooperation [35]. The arms race of coevolution includes multiple layers of defense and counter-defense in which the hosts constantly evolve new means of defense that viruses constantly evolve to evade or suppress [13]. In a clear indication of cooperation, viruses co-op host genes for counter-defense, replication, and movement, and host cells recruit viral genes for diverse roles [10,35].
Viruses evolve in a host-specific manner, accumulating mutations that can increase or decrease fitness, virulence, or infectivity in a certain host [36,37,38]. Interestingly, not all areas of the virus genome accumulate mutations at the same rate. Instead, mutations preferentially accumulate in parts of the genome that are determinants of host adaptation [20,39,40,41].
When a virus is spread to a plant, infection can only occur if the interaction is compatible. This can occur when a host lacks factors, such as R genes, that would otherwise prevent infection from occurring. For example, mutating the BRI1-associated kinase 1 (BAK1) results in increased virus susceptibility [42]. Compatibility can also be mediated by the availability of susceptibility host factors essential for virus replication and by the balance between antiviral defense and the suppression of the evasion of antiviral defense [13]. Mutations in host-susceptibility genes may break the interaction with a critical viral component, reducing or eliminating virus replication or movement [43,44]. In addition, hosts can accumulate mutations in genes that mediate antiviral defense. For example, the dominant-resistance gene Ry(o)phu in Solanum tuberosum groups Phureja and Tuberosum provides protection to potato virus Y (PVY) by blocking viral replication and movement [44]. Similarly, melon accession PI 164323 is resistant to cucumber vein yellowing virus. However, a single amino acid change in the VPg coding region restored compatibility and broke resistance [45].
Often, viruses have a host range limited to a single taxonomic plant family and are not able to infect across multiple plant families [30]. This is explained by fitness optimization to a specific host (niche-filling model) [46]. However, infection across families and even across kingdoms might occur [47], forcing viruses to face strong adaptive selection pressure to maximize their fit to a new niche [46]. When viruses are exposed to genetically uniform hosts, local adaptation occurs [48]. Viruses become specialized to certain hosts due to adaptive tradeoffs where increased fitness in one host leads to decreased fitness in other hosts due to either epistatic interactions or antagonistic pleiotropy [49,50,51]. When tobacco etch potyvirus was sequentially passaged from Nicotiana tabacum into pepper, the virus accumulated genetic changes that increased virulence and coat protein accumulation in pepper at the cost of lesser virulence and accumulation in N. tabacum [33]. When not well-adapted to a host, a virus is less able to hijack the host resources for replication and movement [52]. Bridge hosts may allow the virus to accumulate mutations that support host range expansion without adaptive tradeoffs [53]. Plum pox virus accumulated VPg mutations as it moved from an N. clevelandii-adapted isolate to the bridge-host Arabidopsis thaliana, which allowed partial adaptation to Chenopodium foetidum [53].
Host response can modulate infection, as illustrated by the proteomic comparison between cucumber cultivars resistant to or susceptible to cucumber mosaic virus (CMV). The results showed that the cultivars differentially express photosynthetic, development, stress, and defense-related proteins during infection [54].
Host plants evolve and mutate, forcing viruses to adapt to genetically heterogenous host populations. In plants, ARGONAUTE 2 (AGO2) is an important component of antiviral gene silencing [55]. In A. thaliana, AGO2 shows a high degree of polymorphism that correlates with changes in susceptibility to potato virus X (PVX) [56]. In ecotype Colombia, AGO2 confers resistance to PVX, imposing selection pressure on the virus. In contrast, in ecotype C24, AGO2 confers susceptibility to PVX, potentially imposing selection pressure on the host [56]. Although AGO2 diversity is not the result of co-evolution with viruses, this example illustrates diversity in a host gene, with antiviral activity imposing selection pressure on a virus. While viruses can evolve and adapt rapidly, their hosts may ultimately shape their longer-term evolution [46,57].
1.3. Virus-Vector Co-Evolution
Plant viruses are vectored from plant to plant by a wide variety of insects, nematodes, mites, and fungi [58]. Viruses can be transmitted in a circulative or non-circulative manner with non-persistent, semi-persistent, or persistent subcategories [59]. Virus and vector interactions are highly specialized and under strong selection pressure, in part, as result of co-evolution [11,60,61,62]. For geminiviruses, capsid phylogenies reflect the phylogeny of their vectors rather than that their host plant species, supporting the model that specificity of virus–vector interactions is more stringent than the specificity of virus–plant specificity [63].
Plant viruses often have a narrow vector range: ~60% are vectored by a single species [58]. Poor compatibility between vector and virus can result in the low incidence or elimination of a virus [59]. In southern California, lettuce infectious yellows virus incidence dropped significantly after biotype A of whitefly Bemisia tabaci was replaced by biotype B, a poor vector for the virus [64]. Additionally, mutations in viral proteins that interact with vector factors can eliminate or alter transmission, creating strong selection pressure [65,66]. For example, a single amino acid in the coat protein of squash leaf curl China virus is correlated with increased transmission by Bemisia tabaci Asia-1 compared with Asia-II-1 [67].
Plant viruses can directly or indirectly alter vector behavior and physiology by inducing transcriptional changes in metabolism-related genes in their vectors [68], which alters virus transmission [69]. Viruses transmitted in a nonpersistent manner indirectly mediated positive fitness effects on their vector through changes in the host plant [70]. Vector Aphis gossypii feeds more frequently on virus-infected plants (single or mixed infections), and both Frankliniella occidentalis and A. gossypii showed higher fitness after feeding on virus-infected plants [70,71]. Tomato yellow leaf curl virus (TYLCV) directly affects Bemisia tabaci settling, probing, and feeding behavior, leading to increased transmission efficiency and spread [72]. Tomato spotted wilt tospovirus infection alters host gene expression, resulting in higher total free amino acid content, making it nutritionally advantageous and better suited for thrips vector colonization [71]. Illustrating co-evolution, the specialist aphid Lipaphis erysimi has greater population growth in turnip mosaic virus (TuMV)-infected plants compared with noninfected plants. In contrast, the population growth of a generalist aphid vector Myzus persicae was similar in TuMV-infected and healthy hosts [73]. However, some viruses trick vectors into feeding on poor-quality hosts through volatile emissions [61].
1.4. The Environment and Virus Evolution
Environmental conditions directly impact virus infection, prevalence, evolution, and host interactions. These include abiotic factors like temperature, water stress, CO2 levels, ecology, community makeup, and population heterogeneity [5,26,74,75]. Viruses, hosts, and vectors exist within a given ecosystem, which can impact the spread, disease risk, and symptoms of virus infections. Accordingly, virus evolution is also impacted by ecological factors such as host abundance and species richness, which are affected by climate change and land use [74,76]. In some cases, a loss of biodiversity in host communities is correlated with increased disease [75]. However, adding genetic diversity can amplify disease incidence if hosts are susceptible or host density increases [77]. Additionally, human management and changes in land use result in a higher disease risk due to lowered biodiversity, reduced host diversity, and increased host density [74,78]. In coordination, some virus families are found most prevalently in cultivated areas [79].
While plant viruses are studied primarily in agricultural systems, they are highly prevalent in wild plant populations and often lead to asymptomatic infections, creating possible reservoirs that contribute to virus evolution [76]. Virus infection can be beneficial rather than harmful to hosts experiencing cold damage [80] or drought [81], and environmental conditions alone can shift viruses from being antagonistic to being mutualistic to the host [82]. The environment is likely to be increasingly important for plant virus infections as climate change continues to alter vector, host, and virus properties [5,7].
1.5. Variation, Bottlenecks, Genetic Drift, and Selection Pressure in Virus Populations
Viruses exist as a cloud of related sequences, called a quasispecies, rather than a single consensus [23,24,25]. This genetic variation is the basis for the evolution and adaptation of viruses to hosts, vectors, or their environment [83]. During normal replication, genetic variation is generated through mutation, reassortment, and recombination (Figure 3). These processes underlie the ability of plant viruses to generate and maintain genetic diversity, key to circumventing and adapting to host and vector processes [83]. Mutations can occur through substitutions, insertions, or deletions introduced during the replication by RNA-dependent RNA polymerases that lack proofreading activity [84]. Reassortment occurs between segmented viruses where entire genetic segments are exchanged between related viruses [85]. Recombination involves the transfer of genetic material between parental genomes, often by template switching during genome replication, which leads to new sequence combinations [86,87].
Host and vector factors select for or against variants produced through mutation, recombination, or reassortment [23,83]. Unfit variants are removed and variants with a competitive advantage are fixed in the population (Figure 3). Characteristics of an RNA virus population are determined by interactions between co-infecting viruses in mixed infections, variants in a quasispecies, and selection acting on individual variants, leading to population-level changes [24,25]. Illustrating this, quasispecies cloud sizes change during host shifts, and quasispecies diversity varies across related viruses [88,89]. Accordingly, quasispecies increase the probability a virus can adapt to new hosts, vectors, environments, and challenges during infection [24].
Genetic drift is the change in the frequency of variants in a virus quasispecies due to random chance [90]; some variants may disappear, while some initially rare variants may become more frequent or fixed in the population [90,91]. Similar to selection, genetic drift can decrease variation within a given population while simultaneously increasing the genetic distance between viral populations. Unlike selection, genetic drift results in variants moving onward in the population through random effects rather than a fitness bonus [90,91,92].
Genetic bottlenecks are evolutionary events that cause a sharp reduction in the size and genetic diversity of a virus population in a stochastic manner [90]. The fittest variants may fail to be transmitted due to the small population size generated by these bottleneck events [90]. In this way, bottleneck events counterbalance the genetic variation generated by mutation, reassortment, and recombination [90]. For viruses, vector transmission, the environment, and availability of susceptible hosts may impose genetic bottlenecks that decrease virus genetic diversity [93]. For example, horizontal transmission by aphids caused a significant bottleneck of an artificial population of CMV where variants were consistently lost during transmission [93]. Additionally, bottleneck events occur as a virus moves between hosts and systemically within a host [90,93,94]. These bottlenecks are often extremely narrow, resulting in a small number of variants establishing a new population and potentially generating new strains [88,90]. This founder effect is a form of genetic drift and results in a randomized group of virus sequences moving forward in the population. Founder effects have the potential to eliminate advantageous variants or propagate less advantageous variants [90]. Thus, selection and drift can have opposite effects on virus evolution [91,94]. However, founder effects produced by genetic bottlenecks can result in new species, making bottlenecks an important part of virus evolution [90]. Both processes are important to consider, but distinguishing the effects of the two processes is difficult. Regardless, the generation of variation and maintenance of a diverse quasispecies cloud provides a larger genetic pool for evolutionary forces to act upon irrespective of the fitness benefits or costs to any specific variant.
After selection or bottleneck events, variants become established in the population and emerge as new virus strains or species with novel biological properties, such as pathogenicity or host range expansion [24,35,89]. Therefore, the continuous emergence of new species and strains with novel properties is, in part, explained by natural variation generated through mutation, reassortment, and recombination.
1.6. Host Adaptation
Host adaptation is the development of higher fitness in a host, mediated by the accumulation of genetic changes in key parts of the viral genome. New species, strains, or genetic variants may be better suited to a particular host and thus be selected for in the population [21,45,48]. Over time, the virus will evolve towards an optimal balance with the host: replicating and moving without killing the host before transmission [63]. Similarly, on the host side, tolerance can occur when virus and host fitness are balanced [95]. When related viruses have differences in features such as host range, symptoms, or resistance, one approach to studying viral determinants of host adaptation is to identify the changes in the genome that mediate the contrasting differences. This approach first identifies the novel property and then identifies the genetic changes. For example, a narrow host range strain of CMV was identified, which suggests lack of variation in a viral determinant of the host range [96]. Pathogenicity and host range were mapped to the 2b protein by swapping the gene between different strains [96,97]. A consequence of this approach is that results are often placed within the context of a particular virus in a specific spatial or temporal environment [20].
2. Mutation
Mutations occur through nucleotide insertions, deletions, or substitutions in viral genomes. Mutations initially result in small-scale changes in the genome sequence. However, accumulations of mutations result in large-scale changes, generating novel genomes evident as new strains with novel properties. RNA viruses have a substitution rate of 10−6 to 10−4 substitutions per nucleotide per cell infection, compared with 10−8 to 10−6 substitutions per nucleotide per cell infection for DNA viruses [98]. That is, RNA viruses mutate at a rate that is 100-fold faster than DNA viruses. This is explained by the low fidelity and lack of proofreading activity of RNA-dependent RNA polymerases that replicate viral RNA genomes compared with DNA polymerases [84]. However, confounding factors such as the transmission method and cell tropism can alter substitution rates [99,100]. Single-stranded genomes and viruses with small genomes also show higher mutation rates, blurring the evolutionary advantages of RNA vs DNA viruses [98]. As a virus replicates, unique mutations accumulate in each of the progeny genomes, creating genomes distinct from the parent sequence [101]. Strong intracellular selection and competition occur as these unique sequences replicate and move within the host [101].
Correct virus orthology is identified by nucleotide or amino acid similarity cutoffs [102]. Within a species, mutations have been directly implicated in the generation of new biological properties. For example, a single amino acid mutation allows plum pox virus C and PVY to host jump [103,104]. A single mutation cannot result in a new species [105]. In contrast, a single mutation may result in a new strain if it changes the biological properties of the virus [91,106]. Specific mutations have also allowed viruses to break resistance in important crops such as soybean, melon, and pepper [19,107,108,109]. Further, mutations can alter symptoms in a virus-infected host [110,111], alter the function of virus factors [112], or impact vector transmission [65]. Biologically, mutations in the virus genome are necessary to accommodate the variation present in host proteins [56,113]. Indeed, the ability of a virus genome to tolerate mutations (mutational robustness) is considered an evolutionary advantage in enhancing the establishment of infection in genetically diverse hosts and vectors [36,83].
Experimental mutational analyses can identify the relationships between specific amino acids, protein function, and biological properties. Targeting these conserved amino acids or general areas can result in loss of function, gain of function, or altered symptomatology or virus orthography [110,111,112]. For example, a single amino acid mutation in papaya ringspot virus identified using mutational analyses alters host specificity [114]. Another approach to studying the role of mutation in virus evolution is to identify variation between strains that exhibit different characteristics [115]. However, a negative aspect of this approach is that it is unlikely to identify single mutations that can be traced to a specific biological role.
3. Reassortment
Reassortment, formally called pseudorecombination, occurs when two related viruses co-infect a cell and entire RNA or DNA segments are swapped [116] (Figure 3). Reassortment only occurs in viruses with a segmented genome [91]. Within plant virus families, segmented genomes are the most common genome structure (Figure 2). Reassortment has been documented in almost every plant virus family with a segmented genome and has been responsible for the emergence of new strains with novel properties, including severe symptoms, pathotype differentiation, and resistance breaking (Table 1).
Mixed infections are more likely to occur when viruses and vector have overlapping temporal and spatial ecological niches [132,133]. Transmission mode, virus species, virus titer, host type, and original genetic variation of the virus population can impact the success of these mixed infections [134]. Reassortment can result in viruses containing novel combinations of segments with contrasting phylogenetic and evolutionary history and is especially important for generating genetic diversity in segmented (-) ssRNA viruses [116,135]. The fact that segmented viruses consistently evolve within and across families suggests that reassortment benefits viral fitness and evolution.
Segmented genomes may allow viruses to differentially express genes. During infection, virus segments gravitate towards the “setpoint genome formula” rather than being translated at an equal ratio [116]. This ratio suggests that segmented genomes in part evolved as a way to control the expression of viral proteins [136,137]. Further, this ratio changes from host to host. This indicates that host factors may also play a role in the regulation of the setpoint genome formula and that this process may be specifically advantageous when expanding into new hosts [136,137].
Variants produced through reassortment are subject to selection, and variants with a competitive advantage continue on in the population (Figure 3). Unfit variants are selected out through exposure to host, vector, and viral factors during cell-to-cell movement, systemic movement, and host colonization under bottleneck events [138,139]. Most reassortments result in less-fit variants due to the separation of co-adapted genes [140,141]. While viral genomic stability and the loss of reassorted variants is the status quo [142,143,144], variants that do survive in the population often are key for species differentiation and the evolution of novel characteristics (Table 1).
Reassortment has led to the emergence of new viral strains with novel characteristics and biological properties and to the formation of specific clades in the population [85,145,146,147,148]. Reassortant viruses might be genetically diverse enough to break resistance [85,127], cause novel symptoms [149,150,151], alter host range [152,153], and evolve into new species.
Examining reassortment in wild viral populations illustrates whether variants are emerging and remaining in the population. Measuring reassortment and recombination at the same time in a population allows for the comparison of their frequencies with fewer confounding factors (Table 2). Reassortment occurs at higher frequency than recombination for Alfalfa mosaic virus, in Bunaviridae, and in Secoviridae (Table 2). Reassortment frequency ranges from 3.33% to 55.17%, showing wide diversity across families, species, and likely specific wild populations (Table 2).
4. Recombination
Recombination is a critical source of genetic variation, contributing to the continual evolution of viruses and to the emergence of new species [22]. It occurs both in DNA and RNA viruses [22,160]. In RNA viruses, recombination occurs when the RNA polymerase switches from an initial template (the donor) to a second template (the acceptor) during RNA synthesis, resulting in the fusion of noncontiguous sections of RNAs and creating new combinations of RNA sequences [22,116]. Recombination occurs in RNA replication compartments when both a donor and an acceptor RNA are recruited into the same replication compartment [87]. Template switching occurs in all RNA virus types but is less prevalent in double-stranded and negative-stranded RNA viruses [161]. Theoretically, through recombination, and possibly involving viral or cellular RNA-dependent RNA polymerases, viruses exchange genes with their hosts, and viral genes are often recruited for cellular functions [35,162].
Recombination does not occur randomly across the genome. Instead, recombination preferentially occurs at ‘hotspots’ within a virus genome [148]. It is possible that those hotspots represent areas of the genome that are mutationally robust and have no deleterious effects on the fitness of the novel progeny [83,163]. Most recombinants are less fit and will be selected out of the population (Figure 3) [163].
Across plant virus families, recombination has been identified except in Endornaviridae, Partitiviridae, Pseudoviridae, and Aspiviridae (Table 3). This is likely due to a low recombination rate in these virus families combined with a scarcity of research on the subject with these viruses. However, for the other plant virus families, recombination is a significant evolutionary force that shapes viral species and their features (Table 3). Unlike mutation and reassortment, one recombination event can create variants that are widely divergent from their parental sequences [164,165,166]. This can affect the evolutionary history of viruses by generating new species or strains, thereby shaking up population genetics (Table 3). Consistently, recombination results in viruses epidemiologically important because the recombinants are new species, are resistance breaking, or impact the long-term evolutionary history of the virus [22].
Several new species have been identified that are recombinants of either related or phylogenetically distant viruses. In the genus Begomovirus, tomato yellow leaf curl sardinia virus, tomato yellow leaf curl Axarquia virus, sweet potato leaf curl Canary virus, tomato leaf curl Mahé virus, and tomato leaf curl Yunnan virus are all new species created through recombination [106,164,182].
Recombination frequently shapes the trajectory of a virus population over time [173,179,184]. In Sudan, chickpea chlorotic dwarf virus showed extensive intra- and inter-strain recombination that, in combination with later diversification, led to the foundation of several strains [184]. Several diseases and epidemics have also been tied to recombinants [185,186,187]. This is likely connected with the fact that recombinant viruses can be resistance breaking [171,188]. For example, the spread and origin of Cassava mosaic disease in East Africa has been tied to a recombination event between two cassava mosaic geminiviruses, resulting in increased symptom severity [189]. Recombination can also allow a virus to infect a new host or to increase infectibility [105,165]. A recombination event between lettuce chlorosis virus and bean yellow disorder virus resulted in the recombinant lettuce chlorosis virus-SP that exhibited a shift in host range from lettuce, its original host, to green beans, a new host [185]. However, some recombination events result in decreased host range or infectibility, such as losing systemic infection [190]. This is consistent with the findings that recombination frequency is host specific [191]. Along with new host expansion, recombination can also change the symptoms or virulence of the virus [192,193]. These characteristics showcase the impact of recombination events that, while rare, can severely impact crop safety.
Recombination in wild populations can result in the emergence of novel virus epidemics [86]. In studies measuring both recombination and reassortment, recombination frequency was greater for CMV, Geminiviridae, and Nanoviridae (Table 2). Recombination frequency ranged from 1.67% for AMV to 72.22% for faba bean necrotic yellows virus (FBNYV). For FBNYV, an ancestral recombination event led to all isolates from Spain and Tunisia being recombinants [157]. These observations show that viral population structure can be altered by a single recombination event.
5. Genetically Stationary and Transient Viruses
A virus may move across the continuum between two conceptual genetically evolutionary stages, i.e. stationary (near equilibrium) and transient (far from equilibrium), changing its status as it is exposed to genetically diverse or identical hosts and environments. Viruses in a transient dynamic would be characterized by rapid genetic and phenotypic changes driven by directional or diversifying selection as they adapt to new hosts, vectors or environments. Naturally, these viruses are able to evolve and adapt better and faster to new hosts, vectors, and environments [194]. In general, both potyviruses and orthotospoviruses are constantly changing, and new variants or species are continually detected [42,43,83]. In contrast, viruses in stationary dynamics are characterized by viral populations near an optimal fitness in relation to their current hosts, vectors, and environments and therefore under strong purifying selection. The model is that stable viruses have a narrow host range, and they evolve and adapt slowly to new hosts, vectors, and environments [194]. Stationary viruses, such as maize chlorotic mottle virus, are specialized to infect a narrow range of hosts and vectors [195] and are likely to remain in the ecosystem as infection may benefit or not severely impact both the plant and the virus. Intrinsic properties related to virus genome structure [30], transmission [30], and quasispecies diversity [89] have been correlated with host range. Similarly, this model suggests that genetically transient viruses are mutationally robust and their populations are more likely to contain a variant able to adapt to a variable set of possible hosts without a fitness cost. In contrast, genetically stationary viruses have low mutational robustness and quasispecies diversity, making them less likely to contain advantageous variants [23,36]. While the links between genetic variability, host range, and mutational robustness are currently uncertain, the ability of a virus to adapt to diverse hosts and new environments and to gain novel biological features is likely connected to this evolutionary continuum.
6. Discussion
Viruses evolve through a combination of genetic variation in the population, genetic drift, and selection mediated by host, vector, and environmental factors (Figure 1). Because they replicate faster than their host can reproduce, viruses will continue to change, escaping host immune responses while maintaining functionality in different hosts and vectors [10,15]. Virus variants are produced naturally during the infection cycle through mutation, reassortment, or recombination, showcasing the continuous adaptation of viruses [196]. Based on their fitness or through founder events, variants are selected out of or fixed into viral genomes and virus population structure (Figure 3) [19,41]. Through this process, resistance-breaking virus strains or species with new host ranges or presenting novel symptoms can emerge and lead to epidemics threatening global food security [127,187]. Tomato brown rugose fruit virus (ToBRFV) emerged in 2015 after a recombination event between the major parent, tobacco mosaic virus strain Ohio, and the minor parent, tomato mild mottle virus [86]. Six amino acid substitutions in the movement protein of ToBRFV allowed it to overcome the Tm-22 resistance gene, threatening global tomato production [197,198].
Virus taxonomy and species identification are important for virologists and the framing of their research [28,199]. High-throughput sequencing has revolutionized the identification of novel species in a wide range of hosts [74]. The known virome has subsequently expanded, and the need for classification has grown in coordination [200,201,202]. Currently, virus species are differentiated by sequence similarity cutoffs between related viruses [102]. However, reassortment and recombination often result in new virus species or strains with proteins exhibiting distinct evolutionary histories (Table 1 and Table 3) [105]. Reassortment and recombination are fundamental contributors to the evolution and differentiation of viruses and have high importance for the classification of new species and genera. When identifying species and strains, the genetic variation between and within species, the frequency of reassortment and recombination, and the phylogenetic history of viral proteins should be taken into account [102,105]. The identification of new viruses can take place in coordination with mutation, reassortment, and recombination analyses to identify the evolutionary history of the sequence and provide possible explanations for novel biological properties. Moving forward, more attention should be paid to investigating the sources of variation rather than simply showing it is present.
Measuring reassortment and recombination in wild populations remains difficult. However, occasional variants in wild populations emerge with significant economic and evolutionary effects (Table 1 and Table 3) [127]. Additionally, reassortment and recombination are commonly seen in phylogenetic and evolutionary studies of virus species over time [85,184] that show that these processes are occurring and shaping the genetic structures of wild virus populations. Further, wild population studies are more likely to reflect true recombination and reassortment frequencies because they incorporate all host, vector, and viral interactions. To support this, studies often see very different frequency rates between wild and experimental populations [203]. The frequency at which these events occur, while statistically rare [142], still results in epidemiologically important viruses and thus cannot be ignored. By examining wild population studies that measure both reassortment and recombination in the same environment, we can better understand the importance and frequency of these processes (Table 2). Further, wild and asymptomatic populations can further expand our knowledge of how these processes occur and the lasting impacts of these variants [24]. While reassortment and recombination show clear variation across families, between DNA and RNA viruses, and within families, they are still occurring within the natural population and thus should not be discredited as major evolutionary processes.
Author Contributions
Conceptualization, K.L. and H.G.-R.; methodology, K.L. and H.G.-R.; validation, K.L. and H.G.-R.; formal analysis, K.L.; investigation, K.L.; resources, H.G.-R.; data curation, K.L.; writing—original draft preparation, K.L. and H.G.-R.; writing—review and editing, K.L. and H.G.-R.; visualization, K.L. and H.G.-R.; supervision, H.G.-R.; project administration, H.G.-R.; funding acquisition, H.G.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NIH, grant number R01GM120108, and by the Nebraska Agricultural Experiment Station with funding from the Hatch Act (Accession Number 1007272) through the USDA National Institute of Food and Agriculture. Open access costs were provided by the same grant.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the Holland Computing Center for technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Godfray, H.C.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Sastry, K.S.; Zitter, T.A. Management of Virus and Viroid Diseases of Crops in the Tropics. In Plant Virus and Viroid Diseases in the Tropics: Volume 2: Epidemiology and Management; Sastry, K.S., A. Zitter, T., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 149–480. [Google Scholar]
- Jones, R.A.C.; Naidu, R.A. Global Dimensions of Plant Virus Diseases: Current Status and Future Perspectives. Annu. Rev. Virol. 2019, 6, 387–409. [Google Scholar] [CrossRef]
- Jones, R.A.C. Chapter Three—Future Scenarios for Plant Virus Pathogens as Climate Change Progresses. In Advances in Virus Research; Kielian, M., Maramorosch, K., Mettenleiter, T.C., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 95, pp. 87–147. [Google Scholar]
- Cooper, J.I.; Kuehne, T.; Polischuk, V.P. Virus Diseases and Crop Biosecurity: NATO Sponsered Workshop, Held in Kiev, Ukraine during 4–7 May 2005; Cooper, J.I., Ed.; Springer: Dordrecht, The Netherlands, 2006; p. 148. [Google Scholar]
- Trebicki, P. Climate change and plant virus epidemiology. Virus. Res. 2020, 286, 198059. [Google Scholar] [CrossRef]
- Elena, S.F.; Fraile, A.; García-Arenal, F. Chapter Three—Evolution and Emergence of Plant Viruses. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 88, pp. 161–191. [Google Scholar]
- Makinen, K.; Hafren, A. Intracellular coordination of potyviral RNA functions in infection. Front. Plant Sci. 2014, 5, 110. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Susceptibility Genes to Plant Viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef]
- Fereres, A.; Raccah, B. Plant Virus Transmission by Insects. In eLS; John, W., Sons, L., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–12. [Google Scholar]
- Heinlein, M. Plant virus replication and movement. Virology 2015, 479–480, 657–671. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef]
- Hafren, A.; Ustun, S.; Hochmuth, A.; Svenning, S.; Johansen, T.; Hofius, D. Turnip Mosaic Virus Counteracts Selective Autophagy of the Viral Silencing Suppressor HCpro. Plant Physiol. 2018, 176, 649–662. [Google Scholar] [CrossRef]
- Palukaitis, P.; Yoon, J.Y. R gene mediated defense against viruses. Curr. Opin. Virol. 2020, 45, 1–7. [Google Scholar] [CrossRef]
- Bendahmane, A.; Kanyuka, K.; Baulcombe, D.C. The Rx gene from potato controls separate virus resistance and cell death responses. Plant Cell 1999, 11, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Brommonschenkel, S.H.; Frary, A.; Frary, A.; Tanksley, S.D. The broad-spectrum tospovirus resistance gene Sw-5 of tomato is a homolog of the root-knot nematode resistance gene Mi. Mol. Plant Microbe Interact. 2000, 13, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Hajimorad, M.R.; Hill, J.H. Rsv1-mediated resistance against soybean mosaic virus-N is hypersensitive response-independent at inoculation site, but has the potential to initiate a hypersensitive response-like mechanism. Mol. Plant Microbe Interact. 2001, 14, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Heo, K.J.; Kwon, S.J.; Kim, M.K.; Kwak, H.R.; Han, S.J.; Kwon, M.J.; Rao, A.L.N.; Seo, J.K. Newly emerged resistance-breaking variants of cucumber mosaic virus represent ongoing host-interactive evolution of an RNA virus. Virus Evol. 2020, 6, veaa070. [Google Scholar] [CrossRef] [PubMed]
- Desbiez, C.; Domingo-Calap, M.L.; Pitrat, M.; Wipf-Scheibel, C.; Girardot, G.; Ferriol, I.; Lopez-Moya, J.J.; Lecoq, H. Specificity of Resistance and Tolerance to Cucumber Vein Yellowing Virus in Melon Accessions and Resistance Breaking with a Single Mutation in VPg. Phytopathology 2022, 112, 1185–1191. [Google Scholar] [CrossRef]
- Rousseau, E.; Bonneault, M.; Fabre, F.; Moury, B.; Mailleret, L.; Grognard, F. Virus epidemics, plant-controlled population bottlenecks and the durability of plant resistance. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180263. [Google Scholar] [CrossRef]
- Crespo-Bellido, A.; Hoyer, J.S.; Dubey, D.; Jeannot, R.B.; Duffy, S. Interspecies Recombination Has Driven the Macroevolution of Cassava Mosaic Begomoviruses. J. Virol. 2021, 95, e0054121. [Google Scholar] [CrossRef]
- Domingo, E.; Garcia-Crespo, C.; Perales, C. Historical Perspective on the Discovery of the Quasispecies Concept. Annu. Rev. Virol. 2021, 8, 51–72. [Google Scholar] [CrossRef]
- Domingo, E. Quasispecies: Concept and Implications for Virology, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Ali, A.; Roossinck, M.J. Analysis of quasispecies variation in single and mixed viral infection. Virus Evol. 2017, 3, vex037. [Google Scholar] [CrossRef]
- Alcaide, C.; Sardanyes, J.; Elena, S.F.; Gomez, P. Increasing temperature alters the within-host competition of viral strains and influences virus genetic variability. Virus Evol. 2021, 7, veab017. [Google Scholar] [CrossRef]
- Roossinck, M.; Ali, A. Mechanisms of plant virus evolution and identification of genetic bottlenecks: Impact on diseasemanagement. Biotechnol. Plant Dis. Manag. 2007, 109–124. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Davison, A.J.; Dempsey, D.M.; Dutilh, B.E.; Garcia, M.L.; et al. Changes to virus taxonomy and to the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2021). Arch. Virol. 2021, 166, 2633–2648. [Google Scholar] [CrossRef] [PubMed]
- Hull, R. Matthews’ Plant Virology; Academic Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Moury, B.; Fabre, F.; Hebrard, E.; Froissart, R. Determinants of host species range in plant viruses. J. Gen. Virol. 2017, 98, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Sicard, A.; Michalakis, Y.; Gutierrez, S.; Blanc, S. The Strange Lifestyle of Multipartite Viruses. PLoS Pathog. 2016, 12, e1005819. [Google Scholar] [CrossRef] [PubMed]
- Traore, O.; Pinel-Galzi, A.; Issaka, S.; Poulicard, N.; Aribi, J.; Ake, S.; Ghesquiere, A.; Sere, Y.; Konate, G.; Hebrard, E.; et al. The adaptation of Rice yellow mottle virus to the eIF(iso)4G-mediated rice resistance. Virology 2010, 408, 103–108. [Google Scholar] [CrossRef]
- Agudelo-Romero, P.; de la Iglesia, F.; Elena, S.F. The pleiotropic cost of host-specialization in Tobacco etch potyvirus. Infect. Genet. Evol. 2008, 8, 806–814. [Google Scholar] [CrossRef]
- Gong, Y.N.; Tang, R.Q.; Zhang, Y.; Peng, J.; Xian, O.; Zhang, Z.H.; Zhang, S.B.; Zhang, D.Y.; Liu, H.; Luo, X.W.; et al. The NIa-Protease Protein Encoded by the Pepper Mottle Virus Is a Pathogenicity Determinant and Releases DNA Methylation of Nicotiana benthamiana. Front. Microbiol. 2020, 11, 102. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V. A virocentric perspective on the evolution of life. Curr. Opin. Virol. 2013, 3, 546–557. [Google Scholar] [CrossRef]
- Wilke, C.O.; Wang, J.L.; Ofria, C.; Lenski, R.E.; Adami, C. Evolution of digital organisms at high mutation rates leads to survival of the flattest. Nature 2001, 412, 331–333. [Google Scholar] [CrossRef]
- Hillung, J.; Cuevas, J.M.; Valverde, S.; Elena, S.F. Experimental evolution of an emerging plant virus in host genotypes that differ in their susceptibility to infection. Evolution 2014, 68, 2467–2480. [Google Scholar] [CrossRef] [Green Version]
- Wallis, C.M.; Stone, A.L.; Sherman, D.J.; Damsteegt, V.D.; Gildow, F.E.; Schneider, W.L. Adaptation of plum pox virus to a herbaceous host (Pisum sativum) following serial passages. J. Gen. Virol. 2007, 88, 2839–2845. [Google Scholar] [CrossRef] [PubMed]
- Obenauer, J.C.; Denson, J.; Mehta, P.K.; Su, X.; Mukatira, S.; Finkelstein, D.B.; Xu, X.; Wang, J.; Ma, J.; Fan, Y.; et al. Large-scale sequence analysis of avian influenza isolates. Science 2006, 311, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Nigam, D.; Garcia-Ruiz, H. Variation Profile of the Orthotospovirus Genome. Pathogens 2020, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Nigam, D.; LaTourrette, K.; Souza, P.F.N.; Garcia-Ruiz, H. Genome-Wide Variation in Potyviruses. Front. Plant Sci. 2019, 10, 1439. [Google Scholar] [CrossRef] [PubMed]
- Korner, C.J.; Klauser, D.; Niehl, A.; Dominguez-Ferreras, A.; Chinchilla, D.; Boller, T.; Heinlein, M.; Hann, D.R. The immunity regulator BAK1 contributes to resistance against diverse RNA viruses. Mol. Plant Microbe Interact. 2013, 26, 1271–1280. [Google Scholar] [CrossRef]
- Hebrard, E.; Poulicard, N.; Gerard, C.; Traore, O.; Wu, H.C.; Albar, L.; Fargette, D.; Bessin, Y.; Vignols, F. Direct interaction between the Rice yellow mottle virus (RYMV) VPg and the central domain of the rice eIF(iso)4G1 factor correlates with rice susceptibility and RYMV virulence. Mol. Plant Microbe Interact. 2010, 23, 1506–1513. [Google Scholar] [CrossRef]
- Torrance, L.; Cowan, G.H.; McLean, K.; MacFarlane, S.; Al-Abedy, A.N.; Armstrong, M.; Lim, T.Y.; Hein, I.; Bryan, G.J. Natural resistance to Potato virus Y in Solanum tuberosum Group Phureja. Theor. Appl. Genet. 2020, 133, 967–980. [Google Scholar] [CrossRef]
- Jridi, C.; Martin, J.F.; Marie-Jeanne, V.; Labonne, G.; Blanc, S. Distinct viral populations differentiate and evolve independently in a single perennial host plant. J. Virol. 2006, 80, 2349–2357. [Google Scholar] [CrossRef]
- Simmonds, P.; Aiewsakun, P.; Katzourakis, A. Prisoners of war-host adaptation and its constraints on virus evolution. Nat. Rev. Microbiol. 2019, 17, 321–328. [Google Scholar] [CrossRef]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [Green Version]
- Elena, S.F. Local adaptation of plant viruses: Lessons from experimental evolution. Mol. Ecol. 2017, 26, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Bedhomme, S.; Lafforgue, G.; Elena, S.F. Multihost experimental evolution of a plant RNA virus reveals local adaptation and host-specific mutations. Mol. Biol. Evol. 2012, 29, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- McLeish, M.J.; Fraile, A.; Garcia-Arenal, F. Evolution of plant-virus interactions: Host range and virus emergence. Curr. Opin. Virol. 2019, 34, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Perez, M.G.; Pagan, I.; Aragon-Caballero, L.; Caceres, F.; Fraile, A.; Garcia-Arenal, F. Ecological and genetic determinants of Pepino Mosaic Virus emergence. J. Virol. 2014, 88, 3359–3368. [Google Scholar] [CrossRef]
- Correa, R.L.; Sanz-Carbonell, A.; Kogej, Z.; Muller, S.Y.; Ambros, S.; Lopez-Gomollon, S.; Gomez, G.; Baulcombe, D.C.; Elena, S.F. Viral Fitness Determines the Magnitude of Transcriptomic and Epigenomic Reprograming of Defense Responses in Plants. Mol. Biol. Evol. 2020, 37, 1866–1881. [Google Scholar] [CrossRef]
- Martinez-Turino, S.; Calvo, M.; Bedoya, L.C.; Zhao, M.; Garcia, J.A. Virus Host Jumping Can Be Boosted by Adaptation to a Bridge Plant Species. Microorganisms 2021, 9, 805. [Google Scholar] [CrossRef]
- Novakova, S.; Subr, Z.; Kovac, A.; Fialova, I.; Beke, G.; Danchenko, M. Cucumber mosaic virus resistance: Comparative proteomics of contrasting Cucumis sativus cultivars after long-term infection. J. Proteom. 2020, 214, 103626. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H.; Carbonell, A.; Hoyer, J.S.; Fahlgren, N.; Gilbert, K.B.; Takeda, A.; Giampetruzzi, A.; Garcia Ruiz, M.T.; McGinn, M.G.; Lowery, N.; et al. Roles and programming of Arabidopsis ARGONAUTE proteins during Turnip mosaic virus infection. PLoS Pathog. 2015, 11, e1004755. [Google Scholar] [CrossRef]
- Brosseau, C.; Bolaji, A.; Roussin-Leveillee, C.; Zhao, Z.; Biga, S.; Moffett, P. Natural variation in the Arabidopsis AGO2 gene is associated with susceptibility to potato virus X. New Phytol. 2020, 226, 866–878. [Google Scholar] [CrossRef]
- Dolja, V.V.; Krupovic, M.; Koonin, E.V. Deep Roots and Splendid Boughs of the Global Plant Virome. Annu. Rev. Phytopathol. 2020, 58, 23–53. [Google Scholar] [CrossRef]
- Power, A.G.; Flecker, A.S. 17. Virus Specificity in Disease Systems: Are Species Redundant? In The Importance of Species; Kareiva, P., Levin, S.A., Eds.; Princeton University Press: Princeton, NJ, USA, 2003; pp. 330–346. [Google Scholar]
- Blanc, S.; Michalakis, Y. Manipulation of hosts and vectors by plant viruses and impact of the environment. Curr. Opin. Insect. Sci. 2016, 16, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Power, A.G. Community Ecology of Plant Viruses. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 15–26. [Google Scholar]
- Mauck, K.E.; Chesnais, Q.; Shapiro, L.R. Chapter Seven—Evolutionary Determinants of Host and Vector Manipulation by Plant Viruses. In Advances in Virus Research; Malmstrom, C.M., Ed.; Environmental Virology and Virus Ecology; Academic Press: Cambridge, MA, USA, 2018; Volume 101, pp. 189–250. [Google Scholar]
- Lu, G.; Li, S.; Zhou, C.; Qian, X.; Xiang, Q.; Yang, T.; Wu, J.; Zhou, X.; Zhou, Y.; Ding, X.S.; et al. Tenuivirus utilizes its glycoprotein as a helper component to overcome insect midgut barriers for its circulative and propagative transmission. PLoS Pathog. 2019, 15, e1007655. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, P.; Martin, D.P.; Elena, S.F.; Shepherd, D.N.; Roumagnac, P.; Varsani, A. Evolution and ecology of plant viruses. Nat. Rev. Microbiol. 2019, 17, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Wisler, G.C.; Duffus, J.E.; Liu, H.Y.; Li, R.H. Ecology and Epidemiology of Whitefly-Transmitted Closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef]
- Peter, K.A.; Liang, D.; Palukaitis, P.; Gray, S.M. Small deletions in the potato leafroll virus readthrough protein affect particle morphology, aphid transmission, virus movement and accumulation. J. Gen. Virol. 2008, 89, 2037–2045. [Google Scholar] [CrossRef]
- Sin, S.H.; McNulty, B.C.; Kennedy, G.G.; Moyer, J.W. Viral genetic determinants for thrips transmission of Tomato spotted wilt virus. Proc. Natl. Acad. Sci. USA 2005, 102, 5168–5173. [Google Scholar] [CrossRef]
- Pan, L.L.; Chi, Y.; Liu, C.; Fan, Y.Y.; Liu, S.S. Mutations in the coat protein of a begomovirus result in altered transmission by different species of whitefly vectors. Virus Evol. 2020, 6, veaa014. [Google Scholar] [CrossRef]
- Ding, T.B.; Li, J.; Chen, E.H.; Niu, J.Z.; Chu, D. Transcriptome Profiling of the Whitefly Bemisia tabaci MED in Response to Single Infection of Tomato yellow leaf curl virus, Tomato chlorosis virus, and Their Co-infection. Front. Physiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- Wang, Q.; Li, J.; Dang, C.; Chang, X.; Fang, Q.; Stanley, D.; Ye, G. Rice dwarf virus infection alters green rice leafhopper host preference and feeding behavior. PLoS ONE 2018, 13, e0203364. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Dutta, B.; Coolong, T.; Srinivasan, R. A non-persistent aphid-transmitted Potyvirus differentially alters the vector and non-vector biology through host plant quality manipulation. Sci. Rep. 2019, 9, 2503. [Google Scholar] [CrossRef] [Green Version]
- Nachappa, P.; Challacombe, J.; Margolies, D.C.; Nechols, J.R.; Whitfield, A.E.; Rotenberg, D. Tomato Spotted Wilt Virus Benefits Its Thrips Vector by Modulating Metabolic and Plant Defense Pathways in Tomato. Front. Plant Sci. 2020, 11, 575564. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Delafuente, A.; Garzo, E.; Moreno, A.; Fereres, A. A plant virus manipulates the behavior of its whitefly vector to enhance its transmission efficiency and spread. PLoS ONE 2013, 8, e61543. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Honma, T.; Yasaka, R.; Ohshima, K.; Tokuda, M. Effects of infection by Turnip mosaic virus on the population growth of generalist and specialist aphid vectors on turnip plants. PLoS ONE 2018, 13, e0200784. [Google Scholar] [CrossRef] [PubMed]
- Fetters, A.M.; Cantalupo, P.G.; Wei, N.; Robles, M.T.S.; Stanley, A.; Stephens, J.D.; Pipas, J.M.; Ashman, T.L. The pollen virome of wild plants and its association with variation in floral traits and land use. Nat. Commun. 2022, 13, 523. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Nevado, C.; Gavilán, R.G.; Pagan, I. Host Abundance and Identity Determine the Epidemiology and Evolution of a Generalist Plant Virus in a Wild Ecosystem. Phytopathology 2020, 110, 94–105. [Google Scholar] [CrossRef]
- Lacroix, C.; Jolles, A.; Seabloom, E.W.; Power, A.G.; Mitchell, C.E.; Borer, E.T. Non-random biodiversity loss underlies predictable increases in viral disease prevalence. J. R. Soc. Interface 2014, 11, 20130947. [Google Scholar] [CrossRef]
- Liu, X.; Chen, L.F.; Liu, M.; Garcia-Guzman, G.; Gilbert, G.S.; Zhou, S.R. Dilution effect of plant diversity on infectious diseases: Latitudinal trend and biological context dependence. Oikos 2020, 129, 457–465. [Google Scholar] [CrossRef]
- Pagan, I.; Gonzalez-Jara, P.; Moreno-Letelier, A.; Rodelo-Urrego, M.; Fraile, A.; Pinero, D.; Garcia-Arenal, F. Effect of biodiversity changes in disease risk: Exploring disease emergence in a plant-virus system. PLoS Pathog. 2012, 8, e1002796. [Google Scholar] [CrossRef]
- Bernardo, P.; Charles-Dominique, T.; Barakat, M.; Ortet, P.; Fernandez, E.; Filloux, D.; Hartnady, P.; Rebelo, T.A.; Cousins, S.R.; Mesleard, F.; et al. Geometagenomics illuminates the impact of agriculture on the distribution and prevalence of plant viruses at the ecosystem scale. ISME J 2018, 12, 173–184. [Google Scholar] [CrossRef]
- Roossinck, M.J. The good viruses: Viral mutualistic symbioses. Nat. Rev. Microbiol. 2011, 9, 99–108. [Google Scholar] [CrossRef]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Hily, J.M.; Poulicard, N.; Mora, M.A.; Pagan, I.; Garcia-Arenal, F. Environment and host genotype determine the outcome of a plant-virus interaction: From antagonism to mutualism. New Phytol. 2016, 209, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Merleau, N.S.C.; Penisson, S.; Gerrish, P.J.; Elena, S.F.; Smerlak, M. Why are viral genomes so fragile? The bottleneck hypothesis. PLoS Comput Biol 2021, 17, e1009128. [Google Scholar] [CrossRef]
- Steinhauer, D.A.; Domingo, E.; Holland, J.J. Lack of evidence for proofreading mechanisms associated with an RNA virus polymerase. Gene 1992, 122, 281–288. [Google Scholar] [CrossRef]
- Tentchev, D.; Verdin, E.; Marchal, C.; Jacquet, M.; Aguilar, J.M.; Moury, B. Evolution and structure of Tomato spotted wilt virus populations: Evidence of extensive reassortment and insights into emergence processes. J. Gen. Virol. 2011, 92, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Mansour, A.; Ciuffo, M.; Falk, B.W.; Turina, M. A new tobamovirus infecting tomato crops in Jordan. Arch. Virol. 2016, 161, 503–506. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H.; Diaz, A.; Ahlquist, P. Intermolecular RNA Recombination Occurs at Different Frequencies in Alternate Forms of Brome Mosaic Virus RNA Replication Compartments. Viruses 2018, 10, 131. [Google Scholar] [CrossRef]
- Schneider, W.L.; Roossinck, M.J. Genetic diversity in RNA virus quasispecies is controlled by host-virus interactions. J Virol 2001, 75, 6566–6571. [Google Scholar] [CrossRef]
- Schneider, W.L.; Roossinck, M.J. Evolutionarily related Sindbis-like plant viruses maintain different levels of population diversity in a common host. J Virol 2000, 74, 3130–3134. [Google Scholar] [CrossRef]
- Ali, A.; Roossinck, M.J. Genetic Bottlenecks. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 123–131. [Google Scholar]
- Garcia-Arenal, F.; Fraile, A.; Malpica, J.M. Variability and genetic structure of plant virus populations. Annu. Rev. Phytopathol. 2001, 39, 157–186. [Google Scholar] [CrossRef]
- Sun, B.J.; Sun, L.Y.; Tugume, A.K.; Adams, M.J.; Yang, J.; Xie, L.H.; Chen, J.P. Selection pressure and founder effects constrain genetic variation in differentiated populations of soilborne bymovirus Wheat yellow mosaic virus (Potyviridae) in China. Phytopathology 2013, 103, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Li, H.; Schneider, W.L.; Sherman, D.J.; Gray, S.; Smith, D.; Roossinck, M.J. Analysis of genetic bottlenecks during horizontal transmission of Cucumber mosaic virus. J. Virol. 2006, 80, 8345–8350. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Roossinck, M.J. Genetic bottlenecks reduce population variation in an experimental RNA virus population. J. Virol. 2004, 78, 10582–10587. [Google Scholar] [CrossRef] [PubMed]
- Paudel, D.B.; Sanfacon, H. Exploring the Diversity of Mechanisms Associated With Plant Tolerance to Virus Infection. Front. Plant Sci. 2018, 9, 1575. [Google Scholar] [CrossRef] [PubMed]
- Tepfer, M.; Girardot, G.; Feneant, L.; Ben Tamarzizt, H.; Verdin, E.; Moury, B.; Jacquemond, M. A genetically novel, narrow-host-range isolate of cucumber mosaic virus (CMV) from rosemary. Arch. Virol. 2016, 161, 2013–2017. [Google Scholar] [CrossRef]
- Shi, B.J.; Palukaitis, P.; Symons, R.H. Differential virulence by strains of Cucumber mosaic virus is mediated by the 2b gene. Mol. Plant Microbe Interact. 2002, 15, 947–955. [Google Scholar] [CrossRef]
- Sanjuan, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef]
- Duffy, S.; Shackelton, L.A.; Holmes, E.C. Rates of evolutionary change in viruses: Patterns and determinants. Nat. Rev. Genet. 2008, 9, 267–276. [Google Scholar] [CrossRef]
- Hicks, A.L.; Duffy, S. Cell tropism predicts long-term nucleotide substitution rates of mammalian RNA viruses. PLoS Pathog. 2014, 10, e1003838. [Google Scholar] [CrossRef]
- Schulte, M.B.; Draghi, J.A.; Plotkin, J.B.; Andino, R. Experimentally guided models reveal replication principles that shape the mutation distribution of RNA viruses. Elife 2015, 4, e03753. [Google Scholar] [CrossRef]
- Duffy, S.; Seah, Y.M. 98% identical, 100% wrong: Per cent nucleotide identity can lead plant virus epidemiology astray. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1891–1897. [Google Scholar] [CrossRef]
- Calvo, M.; Malinowski, T.; Garcia, J.A. Single amino acid changes in the 6K1-CI region can promote the alternative adaptation of Prunus- and Nicotiana-propagated Plum pox virus C isolates to either host. Mol. Plant Microbe Interact. 2014, 27, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Vassilakos, N.; Simon, V.; Tzima, A.; Johansen, E.; Moury, B. Genetic Determinism and Evolutionary Reconstruction of a Host Jump in a Plant Virus. Mol. Biol. Evol. 2016, 33, 541–553. [Google Scholar] [CrossRef]
- Simmonds, P. Methods for virus classification and the challenge of incorporating metagenomic sequence data. J. Gen. Virol. 2015, 96, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pendon, J.A.; Sanchez-Campos, S.; Fortes, I.M.; Moriones, E. Tomato Yellow Leaf Curl Sardinia Virus, a Begomovirus Species Evolving by Mutation and Recombination: A Challenge for Virus Control. Viruses 2019, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Eggenberger, A.L.; Hajimorad, M.R.; Hill, J.H. Gain of virulence on Rsv1-genotype soybean by an avirulent Soybean mosaic virus requires concurrent mutations in both P3 and HC-Pro. Mol. Plant Microbe Interact. 2008, 21, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Khatabi, B.; Fajolu, O.L.; Wen, R.H.; Hajimorad, M.R. Evaluation of North American isolates of Soybean mosaic virus for gain of virulence on Rsv-genotype soybeans with special emphasis on resistance-breaking determinants on Rsv4. Mol. Plant Pathol. 2012, 13, 1077–1088. [Google Scholar] [CrossRef]
- Guiu-Aragones, C.; Diaz-Pendon, J.A.; Martin-Hernandez, A.M. Four sequence positions of the movement protein of Cucumber mosaic virus determine the virulence against cmv1-mediated resistance in melon. Mol. Plant Pathol. 2015, 16, 675–684. [Google Scholar] [CrossRef]
- Wieczorek, P.; Obrepalska-Steplowska, A. A single amino acid substitution in movement protein of tomato torrado virus influences ToTV infectivity in Solanum lycopersicum. Virus. Res. 2016, 213, 32–36. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Shi, L.; Zhou, Y.; Li, D.; Han, C.; Zhang, Z.; Yu, J. Two distinct sites are essential for virulent infection and support of variant satellite RNA replication in spontaneous beet black scorch virus variants. J. Gen. Virol. 2012, 93, 2718–2728. [Google Scholar] [CrossRef] [Green Version]
- Bertens, P.; Wellink, J.; Goldbach, R.; van Kammen, A. Mutational analysis of the cowpea mosaic virus movement protein. Virology 2000, 267, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Jiggins, F.M.; Halligan, D.L.; Little, T.J. Natural selection drives extremely rapid evolution in antiviral RNAi genes. Curr. Biol. 2006, 16, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Chiang, C.H.; Raja, J.A.; Liu, F.L.; Tai, C.H.; Yeh, S.D. A single amino acid of niapro of papaya ringspot virus determines host specificity for infection of papaya. Mol. Plant Microbe Interact. 2008, 21, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Azzam, O.; Arboleda, M.; Umadhay, K.M.; de los Reyes, J.B.; Cruz, F.S.; Mackenzie, A.; McNally, K.L. Genetic composition and complexity of virus populations at tungro-endemic and outbreak rice sites. Arch. Virol. 2000, 145, 2643–2657. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef]
- Yuksel Ozmen, C.; Khabbazi, S.D.; Khabbazi, A.D.; Gurel, S.; Kaya, R.; Oguz, M.C.; Turan, F.; Rezaei, F.; Kibar, U.; Gurel, E.; et al. Genome composition analysis of multipartite BNYVV reveals the occurrence of genetic re-assortment in the isolates of Asia Minor and Thrace. Sci. Rep. 2020, 10, 4129. [Google Scholar] [CrossRef]
- Codoner, F.M.; Elena, S.F. The promiscuous evolutionary history of the family Bromoviridae. J. Gen. Virol. 2008, 89, 1739–1747. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kil, E.J.; Kwak, H.R.; Kim, M.; Seo, J.K.; Lee, S.; Choi, H.S. Phylogenetic Characterization of Tomato chlorosis virus Population in Korea: Evidence of Reassortment between Isolates from Different Origins. Plant Pathol. J. 2018, 34, 199–207. [Google Scholar] [CrossRef]
- Ohki, T.; Sasaya, T.; Maoka, T. Cylindrical Inclusion Protein of Wheat Yellow Mosaic Virus Is Involved in Differential Infection of Wheat Cultivars. Phytopathology 2019, 109, 1475–1480. [Google Scholar] [CrossRef]
- Gu, H.; Clark, A.J.; de Sa, P.B.; Pfeiffer, T.W.; Tolin, S.; Ghabrial, S.A. Diversity Among Isolates of Bean pod mottle virus. Phytopathology 2002, 92, 446–452. [Google Scholar] [CrossRef]
- Kalyandurg, P.; Gil, J.F.; Lukhovitskaya, N.I.; Flores, B.; Muller, G.; Chuquillanqui, C.; Palomino, L.; Monjane, A.; Barker, I.; Kreuze, J.; et al. Molecular and pathobiological characterization of 61 Potato mop-top virus full-length cDNAs reveals great variability of the virus in the centre of potato domestication, novel genotypes and evidence for recombination. Mol. Plant Pathol. 2017, 18, 864–877. [Google Scholar] [CrossRef] [PubMed]
- Thekke-Veetil, T.; Polashock, J.J.; Marn, M.V.; Plesko, I.M.; Schilder, A.C.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. Population structure of blueberry mosaic associated virus: Evidence of reassortment in geographically distinct isolates. Virus. Res. 2015, 201, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Dangwal, M.; Mishra, R. Variability of Emaravirus Species Associated with Sterility Mosaic Disease of Pigeonpea in India Provides Evidence of Segment Reassortment. Viruses 2017, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Bolus, S.; Braithwaite, K.S.; Grinstead, S.C.; Fuentes-Bueno, I.; Beiriger, R.; Falk, B.W.; Mollov, D. Completion of Maize Stripe Virus Genome Sequence and Analysis of Diverse Isolates. Front. Microbiol. 2021, 12, 684599. [Google Scholar] [CrossRef]
- Roy, A.; Stone, A.L.; Otero-Colina, G.; Wei, G.; Brlansky, R.H.; Ochoa, R.; Bauchan, G.; Schneider, W.L.; Nakhla, M.K.; Hartung, J.S. Reassortment of Genome Segments Creates Stable Lineages Among Strains of Orchid Fleck Virus Infecting Citrus in Mexico. Phytopathology 2020, 110, 106–120. [Google Scholar] [CrossRef]
- Kwon, S.J.; Cho, Y.E.; Kwon, O.H.; Kang, H.G.; Seo, J.K. Resistance-Breaking Tomato Spotted Wilt Virus Variant that Recently Occurred in Pepper in South Korea is a Genetic Reassortant. Plant Dis. 2021, 105, 2771–2775. [Google Scholar] [CrossRef]
- Petrzik, K. Evolutionary forces at work in partitiviruses. Virus Genes 2019, 55, 563–573. [Google Scholar] [CrossRef]
- Yin, X.; Zheng, F.Q.; Tang, W.; Zhu, Q.Q.; Li, X.D.; Zhang, G.M.; Liu, H.T.; Liu, B.S. Genetic structure of rice black-streaked dwarf virus populations in China. Arch. Virol. 2013, 158, 2505–2515. [Google Scholar] [CrossRef]
- Savory, F.R.; Ramakrishnan, U. Asymmetric patterns of reassortment and concerted evolution in Cardamom bushy dwarf virus. Infect. Genet. Evol. 2014, 24, 15–24. [Google Scholar] [CrossRef]
- Chen, L.F.; Rojas, M.; Kon, T.; Gamby, K.; Xoconostle-Cazares, B.; Gilbertson, R.L. A severe symptom phenotype in tomato in Mali is caused by a reassortant between a novel recombinant begomovirus (Tomato yellow leaf curl Mali virus) and a betasatellite. Mol. Plant Pathol. 2009, 10, 415–430. [Google Scholar] [CrossRef]
- Briese, T.; Calisher, C.H.; Higgs, S. Viruses of the family Bunyaviridae: Are all available isolates reassortants? Virology 2013, 446, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Rosa, C. Thrips as the Transmission Bottleneck for Mixed Infection of Two Orthotospoviruses. Plants 2020, 9, 509. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, S.; Michalakis, Y.; Blanc, S. Virus population bottlenecks during within-host progression and host-to-host transmission. Curr. Opin. Virol. 2012, 2, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Chao, L. Levels of selection, evolution of sex in RNA viruses, and the origin of life. J. Theor. Biol. 1991, 153, 229–246. [Google Scholar] [CrossRef]
- Sicard, A.; Yvon, M.; Timchenko, T.; Gronenborn, B.; Michalakis, Y.; Gutierrez, S.; Blanc, S. Gene copy number is differentially regulated in a multipartite virus. Nat. Commun. 2013, 4, 2248. [Google Scholar] [CrossRef]
- Wu, B.; Zwart, M.P.; Sanchez-Navarro, J.A.; Elena, S.F. Within-host Evolution of Segments Ratio for the Tripartite Genome of Alfalfa Mosaic Virus. Sci. Rep. 2017, 7, 5004. [Google Scholar] [CrossRef]
- Lowen, A.C. It’s in the mix: Reassortment of segmented viral genomes. PLoS Pathog. 2018, 14, e1007200. [Google Scholar] [CrossRef]
- Miyashita, S.; Kishino, H. Estimation of the size of genetic bottlenecks in cell-to-cell movement of soil-borne wheat mosaic virus and the possible role of the bottlenecks in speeding up selection of variations in trans-acting genes or elements. J. Virol. 2010, 84, 1828–1837. [Google Scholar] [CrossRef]
- Escriu, F.; Fraile, A.; Garcia-Arenal, F. Constraints to genetic exchange support gene coadaptation in a tripartite RNA virus. PLoS Pathog. 2007, 3, e8. [Google Scholar] [CrossRef]
- White, M.C.; Lowen, A.C. Implications of segment mismatch for influenza A virus evolution. J. Gen. Virol. 2018, 99, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Fraile, A.; Alonso-Prados, J.L.; Aranda, M.A.; Bernal, J.J.; Malpica, J.M.; Garcia-Arenal, F. Genetic exchange by recombination or reassortment is infrequent in natural populations of a tripartite RNA plant virus. J. Virol. 1997, 71, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Arenal, F.; Fraile, A.; Malpica, J.M. Variation and evolution of plant virus populations. Int. Microbiol. 2003, 6, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Arenal, F.; McDonald, B.A. An analysis of the durability of resistance to plant viruses. Phytopathology 2003, 93, 941–952. [Google Scholar] [CrossRef]
- Adegbola, R.O.; Marshall, S.H.; Batuman, O.; Ullman, D.E.; Gilbertson, R.L.; Adkins, S.; Naidu, R.A. Sequence analysis of the medium and small RNAs of impatiens necrotic spot virus reveals segment reassortment but not recombination. Arch. Virol. 2019, 164, 2829–2836. [Google Scholar] [CrossRef]
- Roossinck, M.J. Evolutionary history of Cucumber mosaic virus deduced by phylogenetic analyses. J. Virol. 2002, 76, 3382–3387. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Hirota, K.; Maruyama, K.; Andika, I.B.; Suzuki, N. A possible occurrence of genome reassortment among bipartite rhabdoviruses. Virology 2017, 508, 18–25. [Google Scholar] [CrossRef]
- Grigoras, I.; Ginzo, A.; Martin, D.P.; Varsani, A.; Romero, J.; Mammadov, A.C.; Huseynova, I.M.; Aliyev, J.A.; Kheyr-Pour, A.; Huss, H.; et al. Genome diversity and evidence of recombination and reassortment in nanoviruses from Europe. J. Gen. Virol. 2014, 95, 1178–1191. [Google Scholar] [CrossRef]
- Gu, H.; Zhang, C.; Ghabrial, S.A. Novel Naturally Occurring Bean pod mottle virus Reassortants with Mixed Heterologous RNA1 Genomes. Phytopathology 2007, 97, 79–86. [Google Scholar] [CrossRef]
- Pita, J.S.; Fondong, V.N.; Sangare, A.; Otim-Nape, G.W.; Ogwal, S.; Fauquet, C.M. Recombination, pseudorecombination and synergism of geminiviruses are determinant keys to the epidemic of severe cassava mosaic disease in Uganda. J. Gen. Virol. 2001, 82, 655–665. [Google Scholar] [CrossRef]
- Kim, Y.I.; Si, Y.J.; Kwon, H.I.; Kim, E.H.; Park, S.J.; Robles, N.J.; Nguyen, H.D.; Yu, M.A.; Yu, K.M.; Lee, Y.J.; et al. Pathogenicity and genetic characterisation of a novel reassortant, highly pathogenic avian influenza (HPAI) H5N6 virus isolated in Korea, 2017. Euro Surveill. 2018, 23, 18–00045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höfer, P.; Engel, M.; Jeske, H.; Frischmuth, T. Host Range Limitation of a Pseudorecombinant Virus Produced by Two Distinct Bipartite Geminiviruses. Mol. Plant Microbe Interact. 1997, 10, 1019–1022. [Google Scholar] [CrossRef]
- Idris, A.M.; Mills-Lujan, K.; Martin, K.; Brown, J.K. Melon chlorotic leaf curl virus: Characterization and differential reassortment with closest relatives reveal adaptive virulence in the squash leaf curl virus clade and host shifting by the host-restricted bean calico mosaic virus. J. Virol. 2008, 82, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Bergua, M.; Luis-Arteaga, M.; Escriu, F. Genetic Diversity, Reassortment, and Recombination in Alfalfa mosaic virus Population in Spain. Phytopathology 2014, 104, 1241–1250. [Google Scholar] [CrossRef]
- Lian, S.; Lee, J.S.; Cho, W.K.; Yu, J.; Kim, M.K.; Choi, H.S.; Kim, K.H. Phylogenetic and recombination analysis of tomato spotted wilt virus. PLoS ONE 2013, 8, e63380. [Google Scholar] [CrossRef]
- Rodelo-Urrego, M.; Garcia-Arenal, F.; Pagan, I. The effect of ecosystem biodiversity on virus genetic diversity depends on virus species: A study of chiltepin-infecting begomoviruses in Mexico. Virus Evol. 2015, 1, vev004. [Google Scholar] [CrossRef]
- Kraberger, S.; Kumari, S.G.; Najar, A.; Stainton, D.; Martin, D.P.; Varsani, A. Molecular characterization of faba bean necrotic yellows viruses in Tunisia. Arch. Virol. 2018, 163, 687–694. [Google Scholar] [CrossRef]
- Ferriol, I.; Ferrer, R.M.; Luis-Arteaga, M.; Guerri, J.; Moreno, P.; Rubio, L. Genetic variability and evolution of broad bean wilt virus 1: Role of recombination, selection and gene flow. Arch. Virol. 2014, 159, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.R.; Kim, M.K.; Lee, Y.J.; Seo, J.K.; Kim, J.S.; Kim, K.H.; Cha, B.; Choi, H.S. Molecular Characterization and Variation of the Broad bean wilt virus 2 Isolates Based on Analyses of Complete Genome Sequences. Plant Pathol. J. 2013, 29, 397–409. [Google Scholar] [CrossRef]
- Galvino-Costa, S.B.F.; Figueira, A.D.; Camargos, V.V.; Geraldino, P.S.; Hu, X.J.; Nikolaeva, O.V.; Kerlan, C.; Karasev, A.V. A novel type of Potato virus Y recombinant genome, determined for the genetic strain PVYE. Plant Pathol. 2012, 61, 388–398. [Google Scholar] [CrossRef]
- Chare, E.R.; Holmes, E.C. A phylogenetic survey of recombination frequency in plant RNA viruses. Arch. Virol. 2006, 151, 933–946. [Google Scholar] [CrossRef]
- Nigam, D.; LaTourrette, K.; Garcia-Ruiz, H. Mutations in virus-derived small RNAs. Sci. Rep. 2020, 10, 9540. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Ahlquist, P. Inducible yeast system for Viral RNA recombination reveals requirement for an RNA replication signal on both parental RNAs. J. Virol. 2006, 80, 8316–8328. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Andres, S.; Monci, F.; Navas-Castillo, J.; Moriones, E. Begomovirus genetic diversity in the native plant reservoir Solanum nigrum: Evidence for the presence of a new virus species of recombinant nature. Virology 2006, 350, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, P.; Moriones, E. Recombination as a motor of host switches and virus emergence: Geminiviruses as case studies. Curr. Opin. Virol. 2015, 10, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Heinze, C.; Wobbe, V.; Lesemann, D.E.; Zhang, D.Y.; Willingmann, P.; Adam, G. Pelargonium necrotic spot virus: A new member of the genus Tombusvirus. Arch. Virol. 2004, 149, 1527–1539. [Google Scholar] [CrossRef]
- Hasiów-Jaroszewska, B.; Kuzniar, A.; Peters, S.A.; Leunissen, J.A.M.; Pospieszny, H. Evidence for RNA recombination between distinct isolates of Pepino mosaic virus. Acta Biochim. Pol. 2010, 57, 385–388. [Google Scholar] [CrossRef]
- Bagayoko, I.; Celli, M.G.; Romay, G.; Poulicard, N.; Pinel-Galzi, A.; Julian, C.; Filloux, D.; Roumagnac, P.; Sereme, D.; Bragard, C.; et al. Genetic Diversity of Rice stripe necrosis virus and New Insights into Evolution of the Genus Benyvirus. Viruses 2021, 13, 737. [Google Scholar] [CrossRef]
- Veerakone, S.; Liefting, L.W.; Khan, S.; Pal, C.; Tang, J.; Ward, L.I. Partial biological and molecular characterization of a novel citrivirus from Nandina domestica. Arch. Virol. 2021, 166, 1395–1399. [Google Scholar] [CrossRef]
- Jacquemond, M. Chapter 13—Cucumber Mosaic Virus. In Advances in Virus Research; Loebenstein, G., Lecoq, H., Eds.; Viruses and Virus Diseases of Vegetables in the Mediterranean Basin; Academic Press: Cambridge, MA, USA, 2012; Volume 84, pp. 439–504. [Google Scholar]
- Harper, S.J.; Dawson, T.E.; Pearson, M.N. Isolates of Citrus tristeza virus that overcome Poncirus trifoliata resistance comprise a novel strain. Arch. Virol. 2010, 155, 471–480. [Google Scholar] [CrossRef]
- Desbiez, C.; Wipf-Scheibel, C.; Millot, P.; Verdin, E.; Dafalla, G.; Lecoq, H. New species in the papaya ringspot virus cluster: Insights into the evolution of the PRSV lineage. Virus. Res. 2017, 241, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.E.; Vigne, E.; Fuchs, M. Genetic structure and molecular variability of Grapevine fanleaf virus populations. Virus. Res. 2010, 152, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Moonan, F.; Molina, J.; Mirkov, T.E. Sugarcane yellow leaf virus: An emerging virus that has evolved by recombination between luteoviral and poleroviral ancestors. Virology 2000, 269, 156–171. [Google Scholar] [CrossRef] [PubMed]
- Tzanetakis, I.E.; Tsai, C.H.; Martin, R.R.; Dreher, T.W. A tymovirus with an atypical 3′-UTR illuminates the possibilities for 3′-UTR evolution. Virology 2009, 392, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Chavan, R.R.; Pearson, M.N. Molecular characterisation of a novel recombinant Ribgrass mosaic virus strain FSHS. Virol. J. 2016, 13, 29. [Google Scholar] [CrossRef]
- He, M.; Guan, S.Y.; He, C.Q. Evolution of rice stripe virus. Mol. Phylogenet. Evol. 2017, 109, 343–350. [Google Scholar] [CrossRef]
- Pappi, P.G.; Maliogka, V.I.; Amoutzias, G.D.; Katis, N.I. Genetic variation of eggplant mottled dwarf virus from annual and perennial plant hosts. Arch. Virol. 2016, 161, 631–639. [Google Scholar] [CrossRef]
- Iftikhar, R.; Ramesh, S.V.; Bag, S.; Ashfaq, M.; Pappu, H.R. Global analysis of population structure, spatial and temporal dynamics of genetic diversity, and evolutionary lineages of Iris yellow spot virus (Tospovirus: Bunyaviridae). Gene 2014, 547, 111–118. [Google Scholar] [CrossRef]
- Li, Y.; Xia, Z.; Peng, J.; Zhou, T.; Fan, Z. Evidence of recombination and genetic diversity in southern rice black-streaked dwarf virus. Arch. Virol. 2013, 158, 2147–2151. [Google Scholar] [CrossRef]
- Hyder, M.Z.; Shah, S.H.; Hameed, S.; Naqvi, S.M. Evidence of recombination in the Banana bunchy top virus genome. Infect. Genet. Evol. 2011, 11, 1293–1300. [Google Scholar] [CrossRef]
- Scussel, S.; Claverie, S.; Hoareau, M.; Moustache, R.; Delatte, H.; Lefeuvre, P.; Lett, J.M. Tomato leaf curl Mahe virus: A novel tomato-infecting monopartite begomovirus from the Seychelles. Arch. Virol. 2018, 163, 3451–3453. [Google Scholar] [CrossRef]
- Yasaka, R.; Nguyen, H.D.; Ho, S.Y.; Duchene, S.; Korkmaz, S.; Katis, N.; Takahashi, H.; Gibbs, A.J.; Ohshima, K. The temporal evolution and global spread of Cauliflower mosaic virus, a plant pararetrovirus. PLoS ONE 2014, 9, e85641. [Google Scholar] [CrossRef] [PubMed]
- Kraberger, S.; Kumari, S.G.; Hamed, A.A.; Gronenborn, B.; Thomas, J.E.; Sharman, M.; Harkins, G.W.; Muhire, B.M.; Martin, D.P.; Varsani, A. Molecular diversity of Chickpea chlorotic dwarf virus in Sudan: High rates of intra-species recombination—A driving force in the emergence of new strains. Infect. Genet. Evol. 2015, 29, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Simon, A.; Garcia, C.; Velasco, L.; Janssen, D. First natural crossover recombination between two distinct species of the family Closteroviridae leads to the emergence of a new disease. PLoS ONE 2018, 13, e0198228. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, J.; Zheng, G.; Zhang, M.; Zhi, H. Recombinant soybean mosaic virus is prevalent in Chinese soybean fields. Arch. Virol. 2014, 159, 1793–1796. [Google Scholar] [CrossRef]
- Silva, T.F.; Correa, R.L.; Castilho, Y.; Silvie, P.; Belot, J.L.; Vaslin, M.F. Widespread distribution and a new recombinant species of Brazilian virus associated with cotton blue disease. Virol. J. 2008, 5, 123. [Google Scholar] [CrossRef]
- Miras, M.; Sempere, R.N.; Kraft, J.J.; Miller, W.A.; Aranda, M.A.; Truniger, V. Interfamilial recombination between viruses led to acquisition of a novel translation-enhancing RNA element that allows resistance breaking. New Phytol. 2014, 202, 233–246. [Google Scholar] [CrossRef]
- Legg, J.P.; Thresh, J.M. Cassava mosaic virus disease in East Africa: A dynamic disease in a changing environment. Virus. Res. 2000, 71, 135–149. [Google Scholar] [CrossRef]
- Green, K.J.; Funke, C.N.; Chojnacky, J.; Alvarez-Quinto, R.A.; Ochoa, J.B.; Quito-Avila, D.F.; Karasev, A.V. Potato Virus Y (PVY) Isolates from Solanum betaceum Represent Three Novel Recombinants Within the PVY(N) Strain Group and Are Unable to Systemically Spread in Potato. Phytopathology 2020, 110, 1588–1596. [Google Scholar] [CrossRef]
- Ouedraogo, R.S.; Pita, J.S.; Somda, I.P.; Traore, O.; Roossinck, M.J. Impact of Cultivated Hosts on the Recombination of Cucumber Mosaic Virus. J. Virol. 2019, 93, e01770-18. [Google Scholar] [CrossRef]
- Hou, Y.M.; Gilbertson, R.L. Increased pathogenicity in a pseudorecombinant bipartite geminivirus correlates with intermolecular recombination. J. Virol. 1996, 70, 5430–5436. [Google Scholar] [CrossRef] [Green Version]
- Monci, F.; Sanchez-Campos, S.; Navas-Castillo, J.; Moriones, E. A natural recombinant between the geminiviruses Tomato yellow leaf curl Sardinia virus and Tomato yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in Spanish populations. Virology 2002, 303, 317–326. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.S.; Melo, F.L.; Inoue-Nagata, A.K.; Nagata, T.; Kitajima, E.W.; Resende, R.O. Characterization of bean necrotic mosaic virus: A member of a novel evolutionary lineage within the Genus Tospovirus. PLoS ONE 2012, 7, e38634. [Google Scholar] [CrossRef] [PubMed]
- Braidwood, L.; Quito-Avila, D.F.; Cabanas, D.; Bressan, A.; Wangai, A.; Baulcombe, D.C. Maize chlorotic mottle virus exhibits low divergence between differentiated regional sub-populations. Sci. Rep. 2018, 8, 1173. [Google Scholar] [CrossRef]
- Komorowska, B.; Hasiow-Jaroszewska, B.; Elena, S.F. Evolving by deleting: Patterns of molecular evolution of Apple stem pitting virus isolates from Poland. J. Gen. Virol. 2019, 100, 1442–1456. [Google Scholar] [CrossRef]
- Hak, H.; Spiegelman, Z. The Tomato Brown Rugose Fruit Virus Movement Protein Overcomes Tm-2(2) Resistance in Tomato While Attenuating Viral Transport. Mol. Plant Microbe Interact. 2021, 34, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.Y.; Ma, H.Y.; Wang, L.; Tettey, C.; Zhao, M.S.; Geng, C.; Tian, Y.P.; Li, X.D. Identification of genetic determinants of tomato brown rugose fruit virus that enable infection of plants harbouring the Tm-2(2) resistance gene. Mol. Plant Pathol. 2021, 22, 1347–1357. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Siddell, S.G. Recognizing species as a new focus of virus research. PLoS Pathog. 2021, 17, e1009318. [Google Scholar] [CrossRef]
- Dutilh, B.E.; Varsani, A.; Tong, Y.; Simmonds, P.; Sabanadzovic, S.; Rubino, L.; Roux, S.; Munoz, A.R.; Lood, C.; Lefkowitz, E.J.; et al. Perspective on taxonomic classification of uncultivated viruses. Curr. Opin. Virol. 2021, 51, 207–215. [Google Scholar] [CrossRef]
- Rumbou, A.; Vainio, E.J.; Buttner, C. Towards the Forest Virome: High-Throughput Sequencing Drastically Expands Our Understanding on Virosphere in Temperate Forest Ecosystems. Microorganisms 2021, 9, 1730. [Google Scholar] [CrossRef]
- Maclot, F.; Candresse, T.; Filloux, D.; Malmstrom, C.M.; Roumagnac, P.; van der Vlugt, R.; Massart, S. Illuminating an Ecological Blackbox: Using High Throughput Sequencing to Characterize the Plant Virome Across Scales. Front. Microbiol. 2020, 11, 578064. [Google Scholar] [CrossRef]
- Desbiez, C.; Joannon, B.; Wipf-Scheibel, C.; Chandeysson, C.; Lecoq, H. Recombination in natural populations of watermelon mosaic virus: New agronomic threat or damp squib? J. Gen. Virol. 2011, 92, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A model of virus evolution driven by the interactions between a virus, host, and vector. These interactions, combined with genetic drift and selective pressure, generate new variants. Contributing variables are indicated for viruses, vectors, and host plants.
Figure 1.
A model of virus evolution driven by the interactions between a virus, host, and vector. These interactions, combined with genetic drift and selective pressure, generate new variants. Contributing variables are indicated for viruses, vectors, and host plants.

Figure 2.
Frequency of plant virus families with multiple and single genomic segments. Some families contain species in both categories. Families are color coded.
Figure 2.
Frequency of plant virus families with multiple and single genomic segments. Some families contain species in both categories. Families are color coded.

Figure 3.
Generation of genetic variants through mutation, reassortment, and recombination. After initial infection, diverse genomes within the same host exist and may mutate, reassort, or recombine. These variants are exposed to genetic drift and selection pressure by host and vector factors. Random or competitive variants are passed on and may become fixed in the population and emerge as new strains or species with novel biological properties.
Figure 3.
Generation of genetic variants through mutation, reassortment, and recombination. After initial infection, diverse genomes within the same host exist and may mutate, reassort, or recombine. These variants are exposed to genetic drift and selection pressure by host and vector factors. Random or competitive variants are passed on and may become fixed in the population and emerge as new strains or species with novel biological properties.


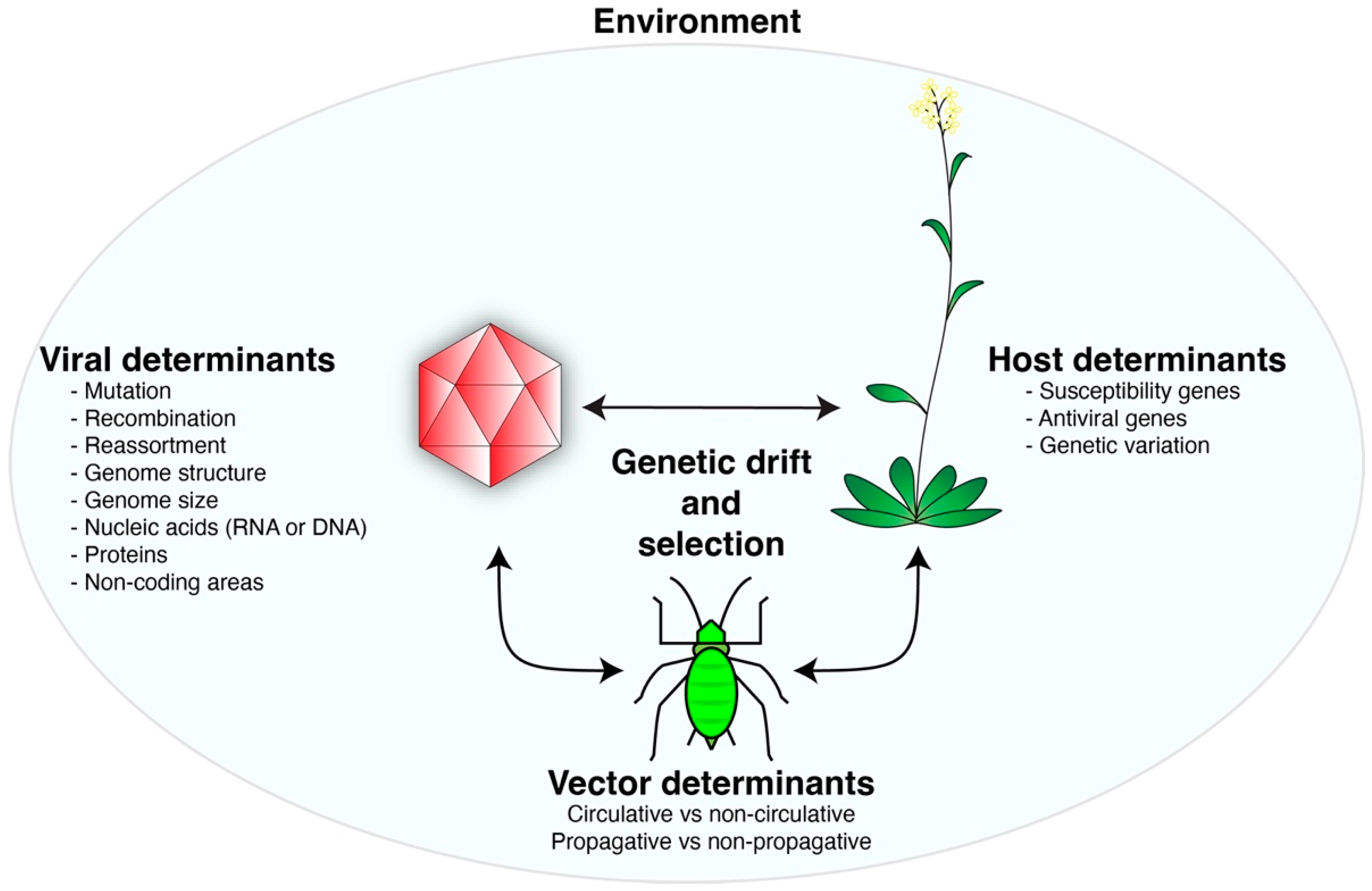{kind=link}
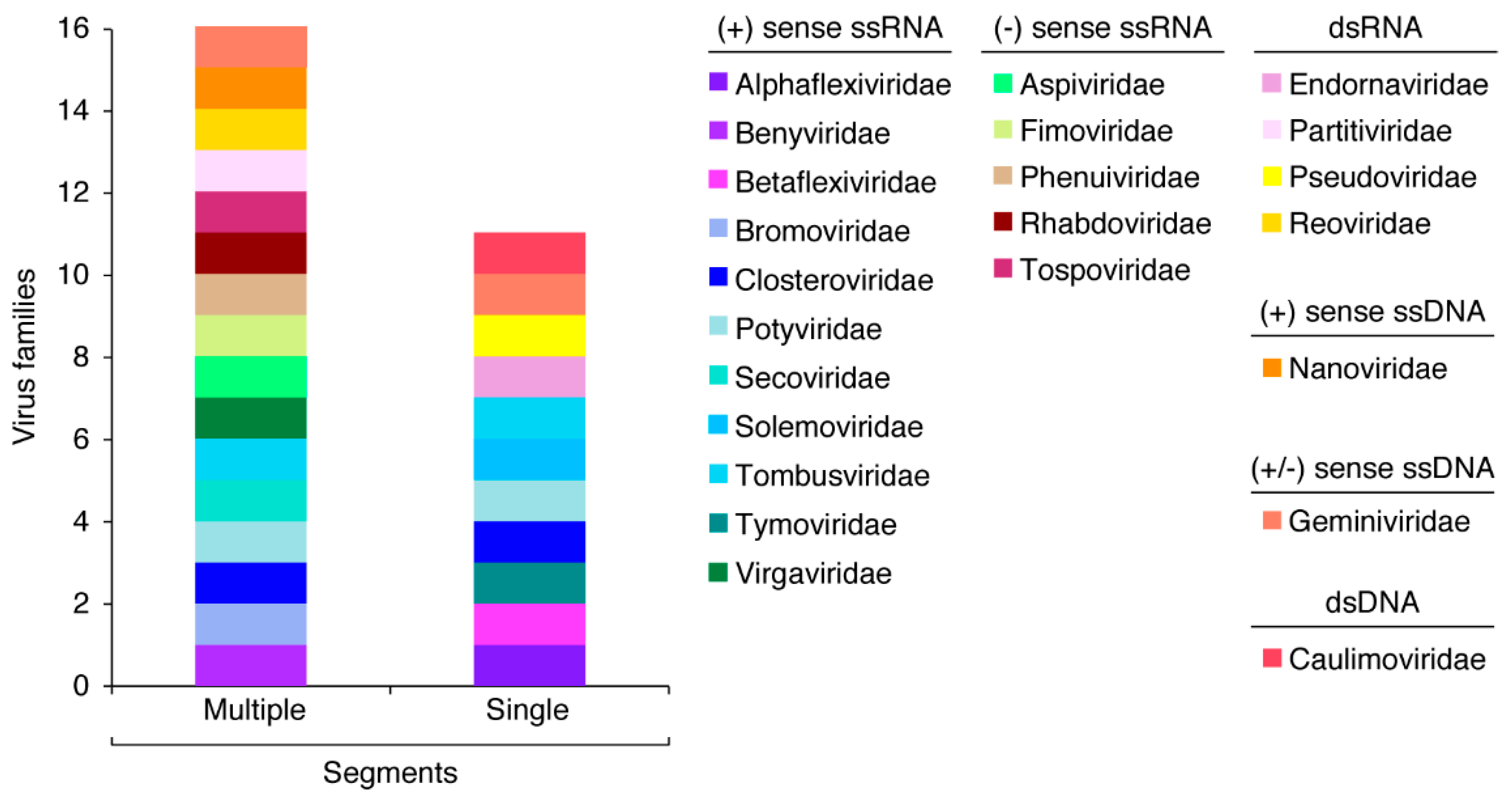{kind=link}
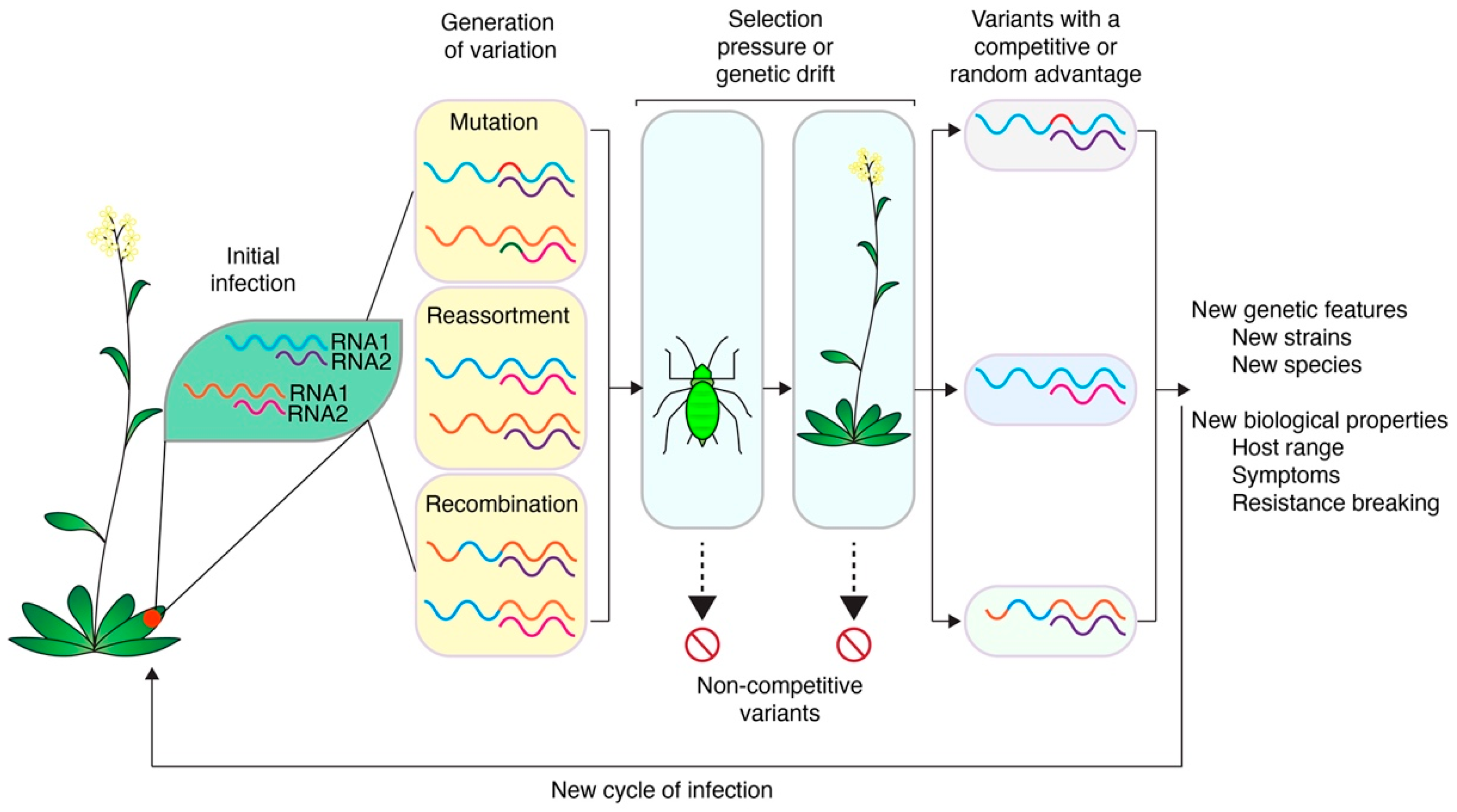{kind=link}
Table 1.
Representative effects of reassortment organized by plant virus family.
| Family | Genus | Species | Reassortment Effect | Reference |
|---|---|---|---|---|
| Benyviridae | Benyvirus | Beet necrotic yellow vein virus | Impact phylogenetic history | [117] |
| Bromoviridae | Bromovirus, Cucumovirus, Ilarvirus, and Oleavirus | 18 species | Impact phylogenetic history | [118] |
| Closteroviridae | Crinivirus | Tomato chlorosis virus | Impact phylogenetic history | [119] |
| Potyviridae | Bymovirus | Wheat yellow mosaic virus | Pathotype differentiation | [120] |
| Secoviridae | Comovirus | Bean pod mottle virus | Severe symptoms | [121] |
| Virgaviridae | Pomovirus | Potato mop-top virus | Impact phylogenetic history | [122] |
| Aspiviridae | Ophiovirus | Blueberry mosaic associated ophiovirus* | Impact phylogenetic history | [123] |
| Fimoviridae | Emaravirus | Pigeonpea sterility mosaic emaravirus 1* and Pigeonpea sterility mosaic emaravirus 2* | Impact phylogenetic history | [124] |
| Phenuiviridae | Tenuivirus | Maize stripe virus | Impact phylogenetic history | [125] |
| Rhabdoviridae | Dichorhavirus | Orchid fleck dichorhavirus* | New strain | [126] |
| Tospoviridae | Orthotospovirus | Tomato spotted wilt orthotospovirus* | Resistance-breaking | [127] |
| Partitiviridae | Alphapartitivirus, Betapartitivirus, Cryspovirus Deltapartitivirus, and Gammapartitivirus | 12 species | Impact phylogenetic history | [128] |
| Reoviridae | Fijivirus | Rice black-streaked dwarf virus | Impact phylogenetic history | [129] |
| Nanoviridae | Babuvirus | Cardamom bushy dwarf virus | Impact phylogenetic history | [130] |
| Geminiviridae | Begomovirus | Tomato yellow leaf curl Mali virus and Cotton leaf curl Gezira betasatellite | Severe symptoms | [131] |
* Name is changed to reflect current ICTV taxonomy.
Table 2.
Reassortment and recombination frequencies in wild viral populations. Frequencies of reassortment and recombination were calculated from papers studying both processes simultaneously in wild virus populations. Frequency was determined by the number of reassorted or recombined sequences identified divided by the total number of virus variants in the sampled population.
Table 2.
Reassortment and recombination frequencies in wild viral populations. Frequencies of reassortment and recombination were calculated from papers studying both processes simultaneously in wild virus populations. Frequency was determined by the number of reassorted or recombined sequences identified divided by the total number of virus variants in the sampled population.
| Family | Species | Reassortment Frequency | Recombination Frequency | Reference |
|---|---|---|---|---|
| Bromoviridae | Alfalfa mosaic virus | 2/60 = 3.33% | 1/60 = 1.67% | [154] |
| Cucumber mosaic virus | 4% | 7% | [142] | |
| Tospoviridae | Tomato spotted wilt orthotospovirus * | 3/13 = 23.08% | 2/13 = 15.38% | [155] |
| Impatiens necrotic spot orthotospovirus * | 6/18 = 33.33% | 0/18 = 0% | [145] | |
| Geminiviridae | Pepper golden mosaic virus | 20/47 = 42.55% | 27/47 = 57.45% | [156] |
| Pepper huasteco yellow vein virus | 11/42 = 26.19% | 13/42= 31.00% | ||
| Nanoviridae | Faba bean necrotic yellows virus | 6/18 = 33.33% | 13/18 = 72.22% | [157] |
| Secoviridae | Broad bean wilt virus 1 | 6/37 = 16.22% | 2/37 = 5.41% | [158] |
| Broad bean wilt virus 2 | 16/29 = 55.17% | 2/29 = 6.70% | [159] |
* Name is changed to reflect current ICTV taxonomy.
Table 3.
Representative effects of recombination organized by plant virus family.
| Family | Genus | Species | Recombination Effect | Reference |
|---|---|---|---|---|
| Alphaflexiviridae | Potexvirus | Pepino mosaic virus | Impact phylogenetic history | [167] |
| Benyviridae | Benyvirus | Rice stripe necrosis virus | Impact phylogenetic history | [168] |
| Betaflexiviridae | Citrivirus | Citrus leaf blotch virus-Rec | New species | [169] |
| Bromoviridae | Cucumovirus | Cucumber mosaic virus | Impact phylogenetic history | [170] |
| Closteroviridae | Closterovirus | Citrus tristeza virus-RB | Resistance-breaking | [171] |
| Potyviridae | Potyvirus | Sudan watermelon mosaic virus | New species | [172] |
| Secoviridae | Nepovirus | Grapevine fanleaf virus | Impact phylogenetic history | [173] |
| Solemoviridae | Polerovirus | Sugarcane yellow leaf virus | New species | [174] |
| Tombusviridae | Tombusvirus | Pelargonium necrotic spot virus | New species | [166] |
| Tymoviridae | Tymovirus | Dulcamara mottle virus | New species | [175] |
| Virgaviridae | Tobamovirus | Ribgrass mosaic virus strain FSHS | New strain | [176] |
| Aspiviridae | NA | NA | NA | NA |
| Fimoviridae | Emaravirus | Pigeonpea sterility mosaic emaravirus 1 and Pigeonpea sterility mosaic emaravirus 2 | Impact phylogenetic history | [124] |
| Phenuiviridae | Tenuivirus | Rice stripe tenuivirus * | Impact phylogenetic history | [177] |
| Rhabdoviridae | Alphanucleorhabdovirus | Eggplant mottled dwarf alphanucleorhabdovirus | Impact phylogenetic history | [178] |
| Tospoviridae | Orthotospovirus | Iris yellow spot orthotospovirus * | Genotype separation | [179] |
| Endornaviridae | NA | NA | NA | NA |
| Partitiviridae | NA | NA | NA | NA |
| Pseudoviridae | NA | NA | NA | NA |
| Reoviridae | Fijivirus | Southern rice black-streaked dwarf virus | Impact phylogenetic history | [180] |
| Nanoviridae | Babuvirus | Banana bunchy top virus | Impact phylogenetic history | [181] |
| Geminiviridae | Begomovirus | Tomato leaf curl Mahé virus | New species | [182] |
| Caulimoviridae | Caulimovirus | Cauliflower mosaic virus | Impact phylogenetic history | [183] |
* Name is changed to reflect current ICTV taxonomy.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
LaTourrette, K.; Garcia-Ruiz, H. Determinants of Virus Variation, Evolution, and Host Adaptation. Pathogens 2022, 11, 1039. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11091039
AMA Style
LaTourrette K, Garcia-Ruiz H. Determinants of Virus Variation, Evolution, and Host Adaptation. Pathogens. 2022; 11(9):1039. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11091039
Chicago/Turabian StyleLaTourrette, Katherine, and Hernan Garcia-Ruiz. 2022. "Determinants of Virus Variation, Evolution, and Host Adaptation" Pathogens 11, no. 9: 1039. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11091039
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.