

Detection and Quantification of Nocardia crassostreae, an Emerging Pathogen, in Mytilus galloprovincialis in the Mediterranean Sea Using Droplet Digital PCR

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and DNA Extraction
2.2. qPCR
2.3. ddPCR
Optimization of the ddPCR and qPCR Protocols
2.4. PCR
2.5. Sequence Analysis
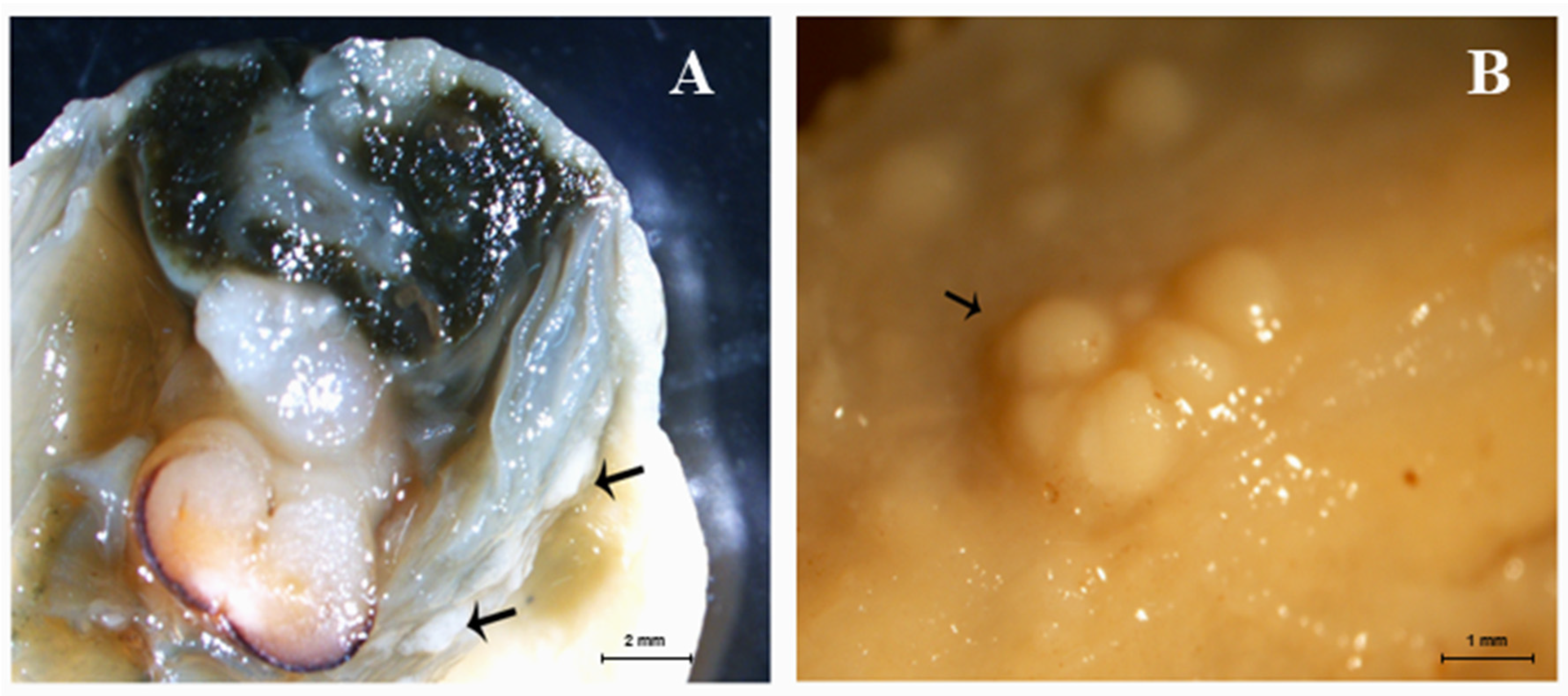
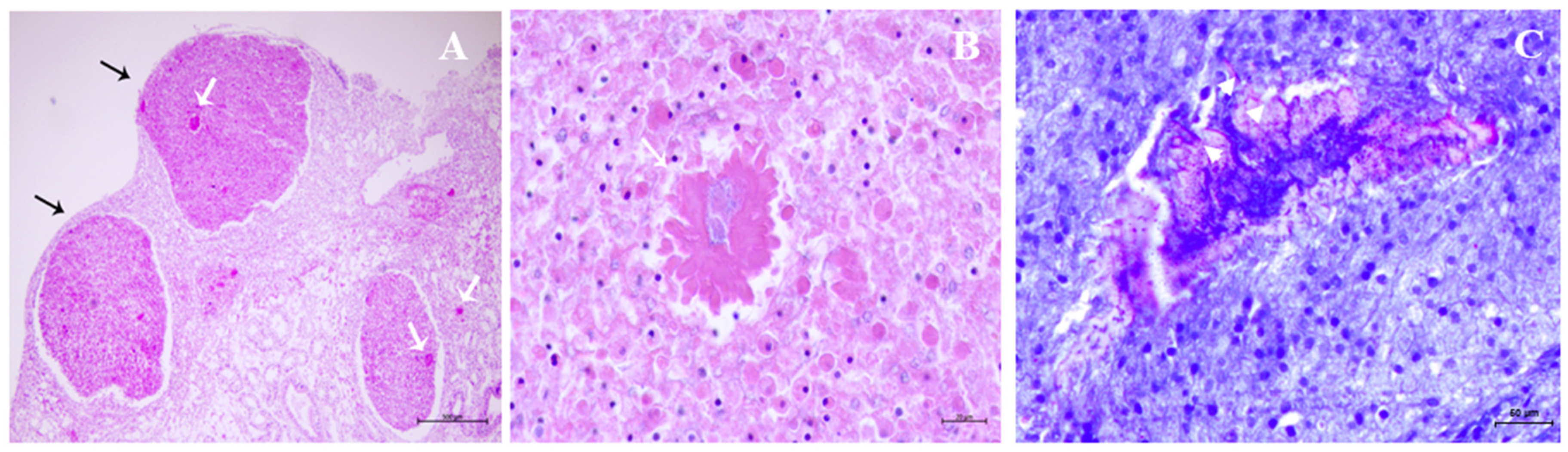
2.6. Gross and Microscopic Findings
3. Statistical Analysis
4. Results
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedman, C.S.; Beaman, B.L.; Chun, J.; Goodfellow, M.; Gee, A.; Hedrick, R.P. Nocardia crassostreae sp. nov., the causal agent of nocardiosis in Pacific oysters. Int. J. Syst. Bacteriol. 1998, 48, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Li, Z.; Huang, Y.; Han, L.; Che, Y.; Hou, X.; Li, D.; Fan, S.; Li, Z. Whole genome sequencing reveals the genomic diversity, taxonomic classification, and evolutionary relationships of the genus Nocardia. PLoS Negl. Trop. Dis. 2021, 15, e0009965. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Brown, J.M.; Conville, P.S.; Wallace, R.J. Clinical and laboratory features of the Nocardia spp. based on the current molecular taxonomy. Clin. Microbiol. Rev. 2006, 19, 259–282. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Barricarte, R. Isolated nocardiosis, an unrecognized primary immunodeficiency? Front. Immunol. 2020, 11, 590239. [Google Scholar] [CrossRef]
- Mehta, H.H.; Shamoo, Y. Pathogenic Nocardia: A diverse genus of emerging pathogens or just poorly recognized? PLoS Pathog. 2020, 16, e1008280. [Google Scholar] [CrossRef] [Green Version]
- Apostolou, A.; Bolcen, S.J.; Dave, V.; Jani, N.; Lasker, B.A.; Tan, C.G.; Montana, B.; Brown, J.M.; Genese, C.A. Nocardia cyriacigeorgica infections attributable to unlicensed cosmetic procedures—An emerging public health problem? Clin. Infect. Dis. 2012, 55, 251–253. [Google Scholar] [CrossRef] [Green Version]
- Taj-Aldeen, S.J.; Deshmukh, A.; Doiphode, S.; Wahab, A.A.; Allangawi, M.A.; AlMuzrkchi, A.; Klassen, C.H.; Meis, J.F. Molecular identification and susceptibility pattern of clinical Nocardia species: Emergence of Nocardia crassostreae as an agent of invasive nocardiosis. Can. J. Infect. Dis. Med. Microbiol. 2013, 24, e33–e38. [Google Scholar] [CrossRef] [Green Version]
- Igbaseimokumo, U.; El Shafie, S.; Al Khal, A.L. First human infection of Nocardia crassostreae in an immunocompetent patient. Chin. Med. J. 2016, 129, 114–115. [Google Scholar] [CrossRef]
- Wright, L.; Katouli, M.; Kurtböke, I. Isolation and characterization of Nocardiae associated with foaming coastal marine waters. Pathogens 2021, 10, 579. [Google Scholar] [CrossRef]
- Engelsma, M.Y.; Roozenburg, I.; Joly, J.P. First isolation of Nocardia crassostreae from Pacific oyster Crassostrea gigas in Europe. Dis. Aquat. Organ. 2008, 80, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Carella, F.; Carrasco, N.; Andree, K.B.; Lacuesta, B.; Furones, D.; De Vico, G. Nocardiosis in Mediterranean bivalves: First detection of Nocardia crassostreae in a new host Mytilus galloprovincialis and in Ostrea edulis from the Gulf of Naples (Italy). J. Invert. Pathol. 2013, 114, 324–328. [Google Scholar] [CrossRef]
- Li, H.; Bai, R.; Zhao, Z.; Tao, L.; Ma, M.; Ji, Z.; Jian, M.; Ding, Z.; Dai, X.; Bao, F.; et al. Application of droplet digital PCR to detect the pathogens of infectious diseases. Biosci. Rep. 2018, 38, BSR2018.1170. [Google Scholar] [CrossRef] [Green Version]
- Hindson, C.M.; Chevillet, J.R.; Briggs, H.A.; Gallichotte, E.N.; Ruf, I.K.; Hindson, B.J.; Vessella, R.L.; Tewari, M. Absolute quantification by droplet digital PCR versus analog real-time PCR. Nat. Methods 2013, 10, 1003–1005. [Google Scholar] [CrossRef]
- Carrasco, N.; Roozenburg, I.; Voorbergen-Laarman, M.; Itoh, N.; Engelsma, M.Y. Development of real-time PCR for detection of the oyster pathogen Nocardia crassostreae based on its homogeneous 16S-23S rRNA intergenic spacer region. J. Invert. Pathol. 2013, 114, 120–127. [Google Scholar] [CrossRef]
- Roperto, S.; Cutarelli, A.; Corrado, F.; De Falco, F.; Buonavoglia, C. Detection and quantification of bovine papillomavirus DNA by digital droplet PCR in sheep blood. Sci. Rep. 2021, 11, 10292. [Google Scholar] [CrossRef]
- De Falco, F.; Cutarelli, A.; Cuccaro, B.; Catoi, C.; De Carlo, E.; Roperto, S. Evidence of a novel cross-species transmission by ovine papillomavirus. Transbound. Emerg. Dis. 2022, 69, 3850–3857. [Google Scholar] [CrossRef]
- Tastanova, A.; Stoffel, C.I.; Dzung, A.; Cheng, P.F.; Bellini, E.; Johansen, P.; Duda, A.; Nobbe, S.; Lienhard, R.; Bosshard, P.P.; et al. A comparative study of real-time RT-PCR –Based SARS-CoV-2 detection methods and its application to human-derived and surface swabbed material. J. Mol. Diagn. 2021, 23, 796–804. [Google Scholar] [CrossRef]
- Luna, L.G. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology; McGrawHill Book Company: New York, NY, USA, 1968; p. 258. [Google Scholar]
- Fernández Robledo, J.A.; Yadavalli, R.; Allam, B.; Pales-Espinosa, E.; Gerdol, M.; Greco, S.; Stevick, R.J.; Gómez-Chiarri, M.; Zhang, Y.; Zhang, Y.; et al. From the raw bar to the bench: Bivalves as models for human health. Dev. Comp. Immunol. 2019, 92, 260–282. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020; Sustainability in Action; FAO: Rome, Italy, 2020; pp. 6–7. [Google Scholar] [CrossRef]
- Jeong, S.H.; Kwon, J.Y.; Shin, S.B.; Choi, W.S.; Lee, J.H.; Kim, S.J.; Ha, K.S. Antibiotic resistance in shellfish and major inland pollution sources in the drainage basin of Kamak Bay, Republic of Korea. Environ. Monit. Assess. 2021, 193, 471. [Google Scholar] [CrossRef]
- Bagi, A.; Kogerbø, G. Tracking bacterial pollution at a marine wastewater outfall site—A case study from Norway. Sci. Total Environ. 2022, 829, 154257. [Google Scholar] [CrossRef]
- Egan, S.; Gardiner, M. Microbial dysbiosis: Rethinking disease in marine ecosystems. Front. Microbiol. 2016, 7, 991. [Google Scholar] [CrossRef]
- Maekawa, S.; Yoshida, T.; Wang, P.C.; Chen, S.C. Current knowledge of nocardiosis in teleost fish. J. Fish Dis. 2018, 41, 413–419. [Google Scholar] [CrossRef]
- Han, H.J.; Kwak, M.J.; Ha, S.M.; Yang, S.J.; Kim, J.D.; Cho, K.H.; Kim, T.W.; Cho, M.Y.; Kim, B.Y.; Jung, S.H.; et al. Genomic characterization of Nocardia seriolae strains isolated from diseased fish. Microbiologyopen 2019, 8, e656. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Santana, P.; Fernández, A.; Díaz-Delgado, J.; Vela, A.I.; Domínguez, L.; Suárez-Santana, C.; Puig-Lozano, R.; Fernández-Maldonado, C.; Sierra, E.; Arbelo, M. Nocardiosis in free-ranging cetaceans from the central-eastern Atlantic Ocean and contiguous Mediterranean Sea. Animals 2022, 12, 434. [Google Scholar] [CrossRef]
- Britt, A.; Bernini, M.; McSweeney, B.; Dalapati, S.; Duchin, S.; Cavanna, K.; Santos, N.; Donovan, G.L.; O’Byrne, K.; Noyes, S.; et al. The effects of atrazine on the microbiome to the eastern oyster: Crassostrea virginica. Sci. Rep. 2020, 10, 11088. [Google Scholar] [CrossRef]
- De Vico, G.; Carella, F. Nocardiosis and mycobacteriosis of bivalves: “Yet-to-emerge” zoonoses of public concern? Zoonoses Public Health 2019, 66, 559–561. [Google Scholar] [CrossRef]
- Lattos, A.; Chaligiannis, I.; Papadopoulos, D.; Giantsis, I.A.; Petridou, E.; Vafeas, G.; Staikou, A.; Michaelidis, B. How safe to eat are raw bivalves? Host pathogenic and public health concern microbes within mussels, oysters, and clams in Greek markets. Foods 2021, 10, 2793. [Google Scholar] [CrossRef]
- Kandi, V. Human Nocardia infections: A review of pulmonary nocardiosis. Cureus 2015, 7, e304. [Google Scholar] [CrossRef] [Green Version]
- Stentiford, G.D.; Bateman, I.J.; Hinchliffe, S.J.; Bass, D.; Hartnell, R.; Santos, E.M.; Devlin, M.J.; Feist, S.W.; Taylor, G.H.; Verner-Jeffreys, D.W.; et al. Suitable aquaculture though the One Health lens. Nat. Food 2020, 1, 468–474. [Google Scholar] [CrossRef]
- Bizzarro, G.; Vatland, A.K.; Pampanin, D.M. The One-Health approach in seaweed production. Environ. Int. 2022, 158, 106948. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sample Set | N | ddPCR | qPCR |
|---|---|---|---|
| Pooled tissues | 185 | 26% | 3% |
| Individual mussels | 149 | 15% | 4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cutarelli, A.; Carella, F.; De Falco, F.; Cuccaro, B.; Di Nocera, F.; Nava, D.; De Vico, G.; Roperto, S. Detection and Quantification of Nocardia crassostreae, an Emerging Pathogen, in Mytilus galloprovincialis in the Mediterranean Sea Using Droplet Digital PCR. Pathogens 2023, 12, 994. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12080994
Cutarelli A, Carella F, De Falco F, Cuccaro B, Di Nocera F, Nava D, De Vico G, Roperto S. Detection and Quantification of Nocardia crassostreae, an Emerging Pathogen, in Mytilus galloprovincialis in the Mediterranean Sea Using Droplet Digital PCR. Pathogens. 2023; 12(8):994. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12080994
Chicago/Turabian StyleCutarelli, Anna, Francesca Carella, Francesca De Falco, Bianca Cuccaro, Fabio Di Nocera, Donatella Nava, Gionata De Vico, and Sante Roperto. 2023. "Detection and Quantification of Nocardia crassostreae, an Emerging Pathogen, in Mytilus galloprovincialis in the Mediterranean Sea Using Droplet Digital PCR" Pathogens 12, no. 8: 994. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12080994