
A New Sturgeon Herpesvirus from Juvenile Lake Sturgeon Acipenser fulvescens Displaying Epithelial Skin Lesions

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
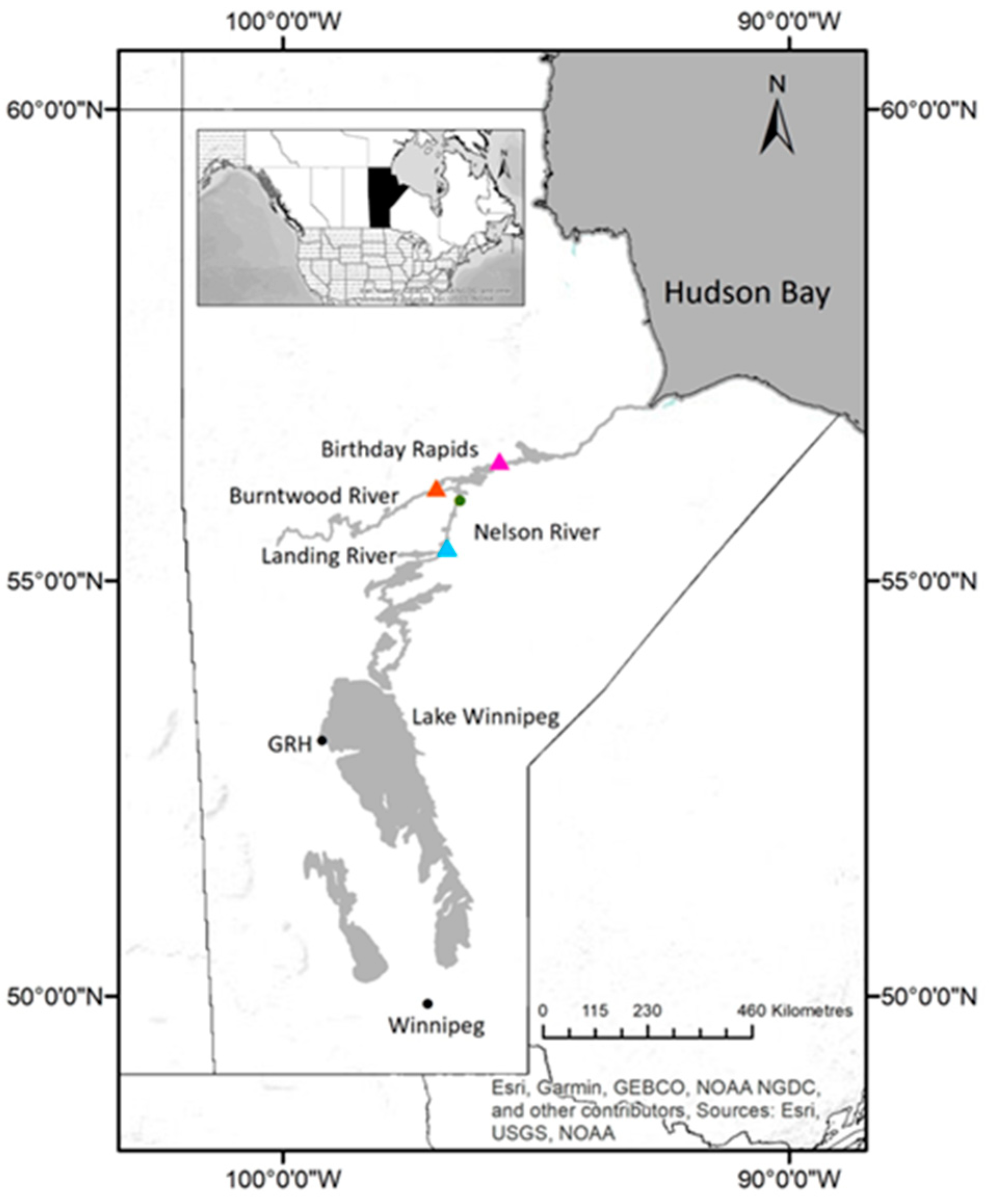
2.1. Case History—Hatchery Case Study
2.1.1. Grand Rapids Hatchery 2017
2.1.2. Grand Rapids Hatchery 2018
2.1.3. Statistical Analyses
2.2. Virology, Bacteriology, Histology and Electron Microscopy
2.2.1. Viruses
2.2.2. Virology—Hatchery Case Study
2.2.3. Bacteriology—Hatchery Case Study
2.2.4. Histology—Hatchery Case Study
2.2.5. Electron Microscopy—Hatchery Case Study
2.3. Viral Nucleic Acid
2.3.1. DNA Synthesis and Plasmid Purification
2.3.2. Nucleic Acid Extraction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Abbreviation | Accession Number(s) | Description of Use in Study d |
|---|---|---|---|
| Alloherpesviridae, Acivirus a | |||
| Acipenser herpesvirus 1 | AciHV-1 | ||
| Lake Sturgeon Black River | AciHV-1 200413-11TC | WAS29283.1, 84, 85, 87, 89 | Systematics [15] |
| Lake Sturgeon Wolf River | AciHV-1 WR | UMM52714.1, 31, 41, 45, 47 | Systematics [16] |
| White Sturgeon UC Davis | AciHV-1 UCD | UKB92883.1, 84, 85, 87, 88 | Systematics [16] |
| Acipenser herpesvirus 3 a | AciHV-3 | ||
| Atlantic Sturgeon AciHV-3 | AciHV-3 JABEPO | JABEPO010010078.1 b | Genotyping e |
| Lake Sturgeon AciHV-3 63-201:210 c | AciHV-3 UNR | OR242753, 54, 55, 56, 57 | Systematics: 2017YC GRH UNR LS |
| Lake Sturgeon AciHV-3 63-207 | AciHV-3 UNR 2017 | OR242743 | Genotyping: 2017YC GRH UNR LS |
| Lake Sturgeon AciHV-3 63-251 | AciHV-3 BWR 2017 | OR242744 | Genotyping: 2017YC GRH BWR LS |
| Lake Sturgeon AciHV-3 71-206 | AciHV-3 LNR 2018 | OR242746 | Genotyping: 2018YC GRH LNR LS |
| Lake Sturgeon AciHV-3 71-201 | AciHV-3 UNR 2018 | OR242745 | Genotyping: 2018YC GRH UNR LS |
| Lake Sturgeon AciHV-3 LS gonad cell line | AciHV-3 LSGO | OR242742 | Genotyping [29] |
| Pallid Sturgeon AciHV-3 MT 2008 | AciHV-3 MT 2008 | OR242752 | Genotyping: DNA from R. Hedrick |
| Shortnose Sturgeon AciHV-3 54-1402 | AciHV-3 NB 2015 | OR242747 | Genotyping: 2013YC SnS (NB hatchery) |
| Sterlet Sturgeon AciHV-3 Gen_M01 | AciHV-3 Gen M01 | VTUV01000924.1 b | Systematics, genotyping e |
| Sterlet Sturgeon AciHV-3 WHYD16114868_AA | AciHV-3 WHYD | SCEB01000000.1 b | Systematics, genotyping e |
| Sterlet Sturgeon AciHV-3 fAciRut3 | AciHV-3 fAciRut3 | OV754656b | Systematics, genotyping e [39] |
| White Sturgeon AciHV-3 63-1 | AciHV-3 UCR 2017 | OR242748 | Genotyping: BC UCR wild WS (2017) |
| White Sturgeon AciHV-3 89-174 | AciHV-3 VIU 2022 | OR242749 | Genotyping: VIU WS (2022) |
| White Sturgeon AciHV-3 WSS-2 cell line | AciHV-3 WSS-2 | OR242751 | Genotyping [28] |
| White Sturgeon AciHV-3 WSSK cell line | AciHV-3 WSSK | OR242750 | Genotyping [7] |
| Alloherpesviridae, Batravirus | |||
| Bufonid herpesvirus 1 FO1_2015 | BfHV-1 | YP_009552889.1, 891, 918, 933, 946 | Systematics |
| Ranid herpesvirus I McKinnell | RaHV-1 | YP_656709.1, 718, 727, 748, 750 | Systematics |
| Ranid herpesvirus 2 Rafferty | RaHV-2 | YP_656588.1, 596, 618, 637, 639 | Systematics |
| Ranid herpesvirus 3 FO1_2015 | RaHV-3 | YP_009362382.1, 385, 398, 405, 419 | Systematics |
| Alloherpesviridae, Cyvirus | |||
| Anguillid herpesvirus 1 500138 | AngHV-1 | YP_003358175.1, 176, 194, 196, 243 | Systematics |
| Cyprinid herpesvirus 1 NG-J1 | CyHV-1 | YP_007003734.1, 735, 741, 742, 755 | Systematics |
| Cyprinid herpesvirus 2 ST-J1 | CyHV-2 | YP_007003890.1, 891, 897, 898, 911 | Systematics |
| Cyprinid herpesvirus 3 F98-50 | CyHV-3 | YP_001096106.1, 107, 113, 114, 127 | Systematics |
| Alloherpesviridae, Ictavirus | |||
| Acipenser herpesvirus 2 | AciHV-2 | ||
| Shortnose Sturgeon AciHV-2 NB_2015 | AciHV-2 NB 2015 | OR242758, 59, 60, 61, 62 | Systematics |
| Snake River White Sturgeon herpesvirus | AciHV-2 SRWSHV | YP_009664566.1, 568, 569, 579, 599 | Systematics |
| Ictalurivirus herpesvirus 1 Auburn 1 | IcHV-1 | NP_041116.1, 118, 119, 130, NP_041148.2 | Systematics |
| Ictalurivirus herpesvirus 2 760/94 | IcHV-2 | YP_009447852.1, 853, 854, 864, 884 | Systematics |
| Silurid herpesvirus 1 KRB14001 | SiHV-1 | AVP72202.1, 03, 04, 14, 65 | Systematics |
| Orthoherpesviridae, Alphaherpesvirinae, Simplexvirus | |||
| Human alphaherpesvirus 1 Strain 17 | HSV-1 | YP_009137079.1, 092, 093, 100, 105 | Systematics |
| Orthoherpesviridae, Gammaherpesvirinae, Rhadinovirus | |||
| Saimiriine gammaherpesvirus 2 | SaHV-2 | NP_040211.1, 219, 227, 228, 246 | Systematics |
2.3.3. PCR and Sanger Sequencing
2.4. Next-Generation Sequencing, Assembly and Annotation
2.4.1. NGS and Assembly of the AciHV-2 Dataset
2.4.2. NGS and Assembly of the AciHV-3 Dataset
2.4.3. Annotation of the AciHV-2 and AciHV-3 Datasets
2.5. Sequence Similarity, Phylogenetics and Genotyping Analyses
2.5.1. Identity Matrix for Alloherpesvirus Major Capsid Protein Sequences
2.5.2. Phylogenetic Analyses
3. Results
3.1. Lesion Gross Pathology and Prevalence
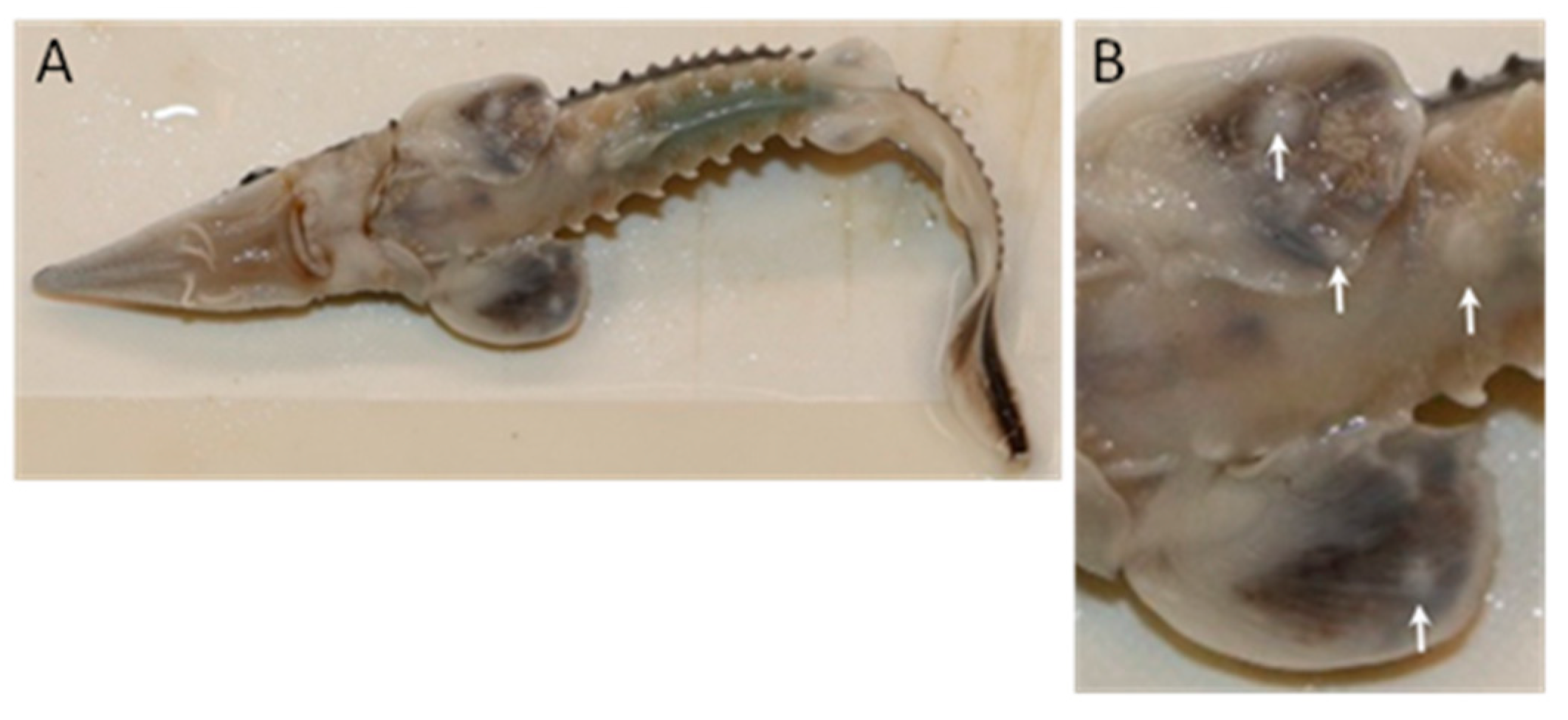
3.1.1. Gross Pathology

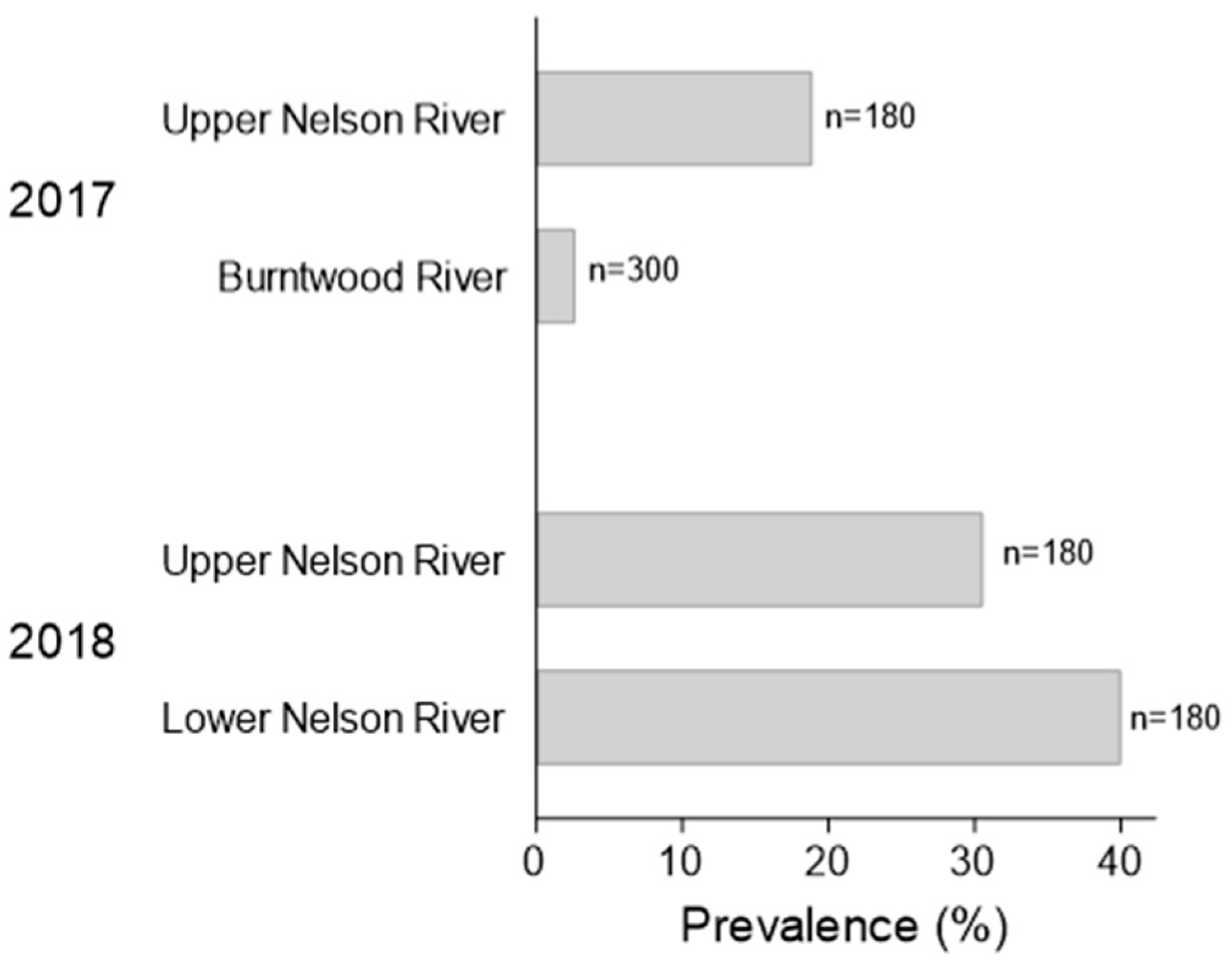
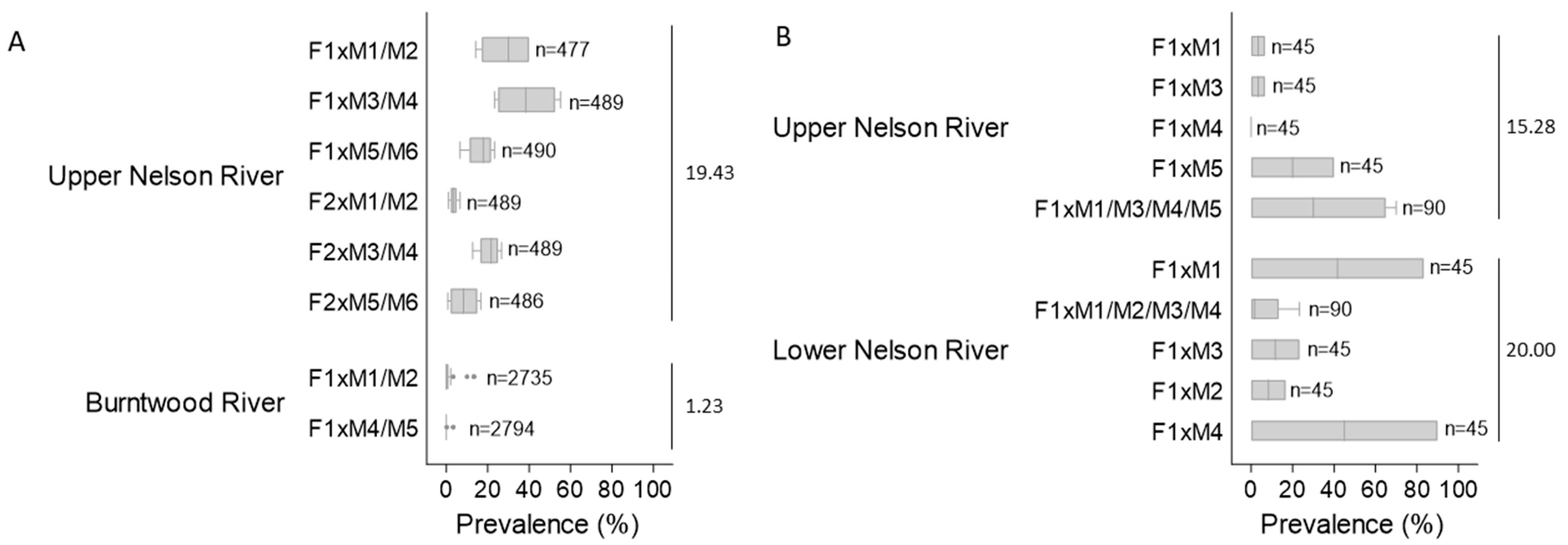
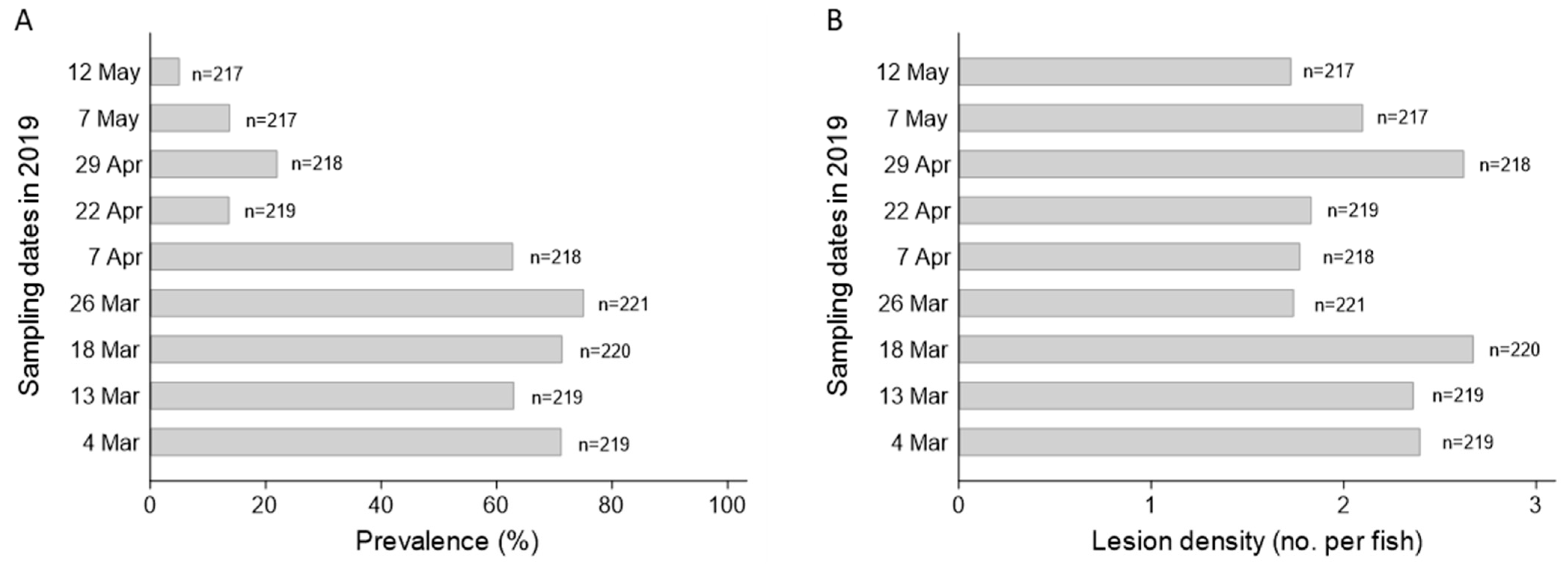
3.1.2. Lesion Prevalence and Density
3.1.3. Bacteriology and Virology
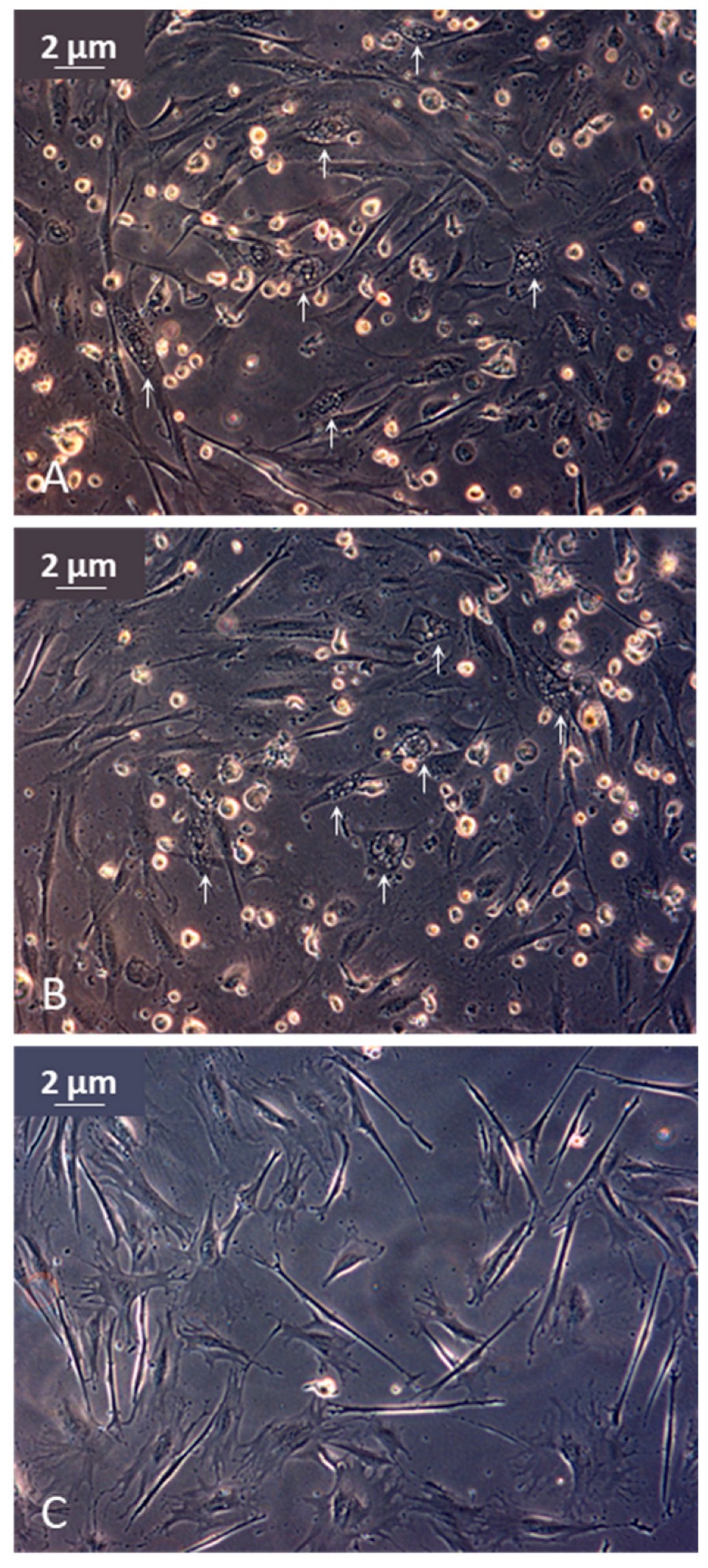
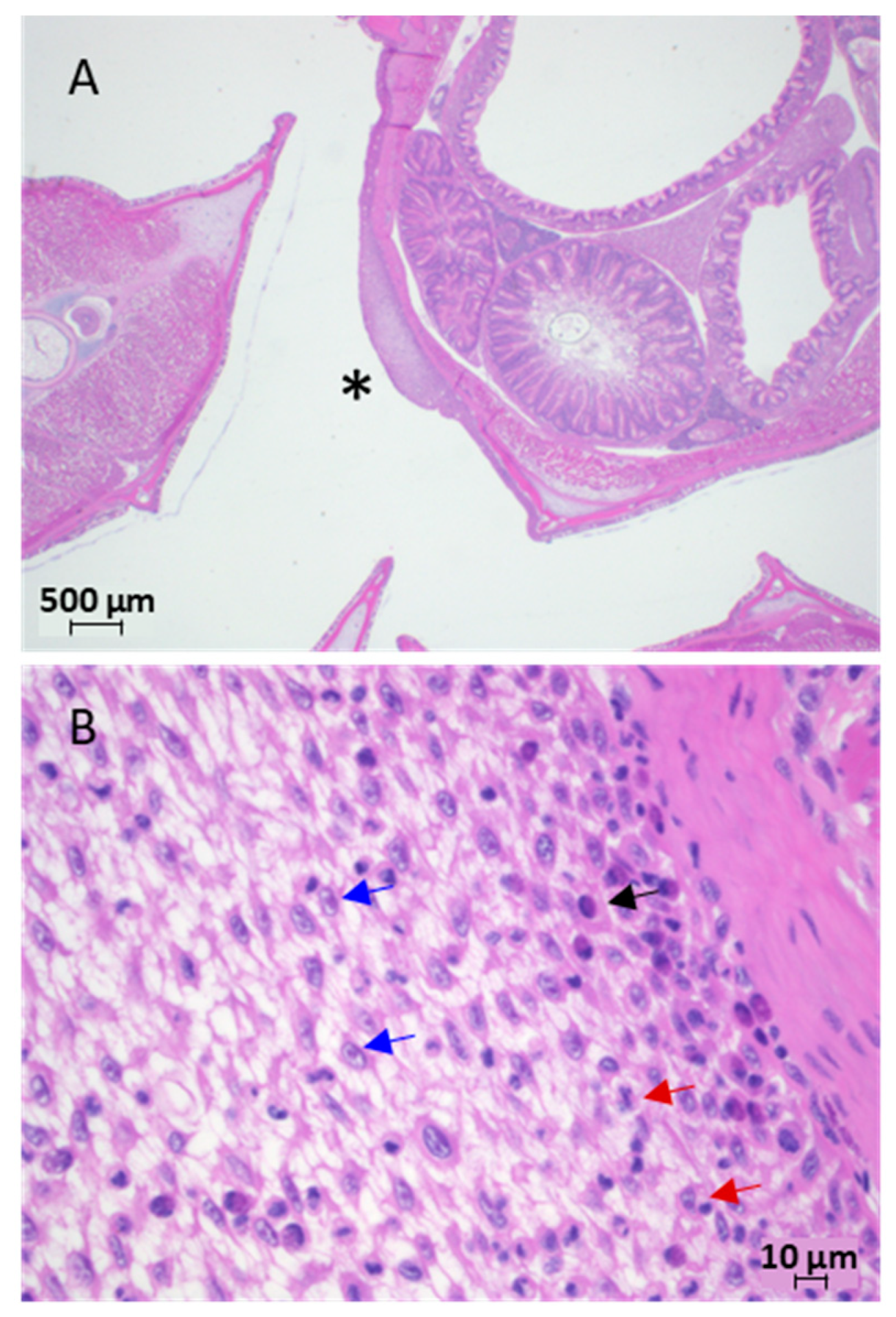
3.1.4. Histology and Electron Microscopy


3.2. AciHV Contigs Encode Alloherpesvirus Core Proteins
3.3. AciHV-3 Major Capsid Protein Sequences Are Highly Conserved
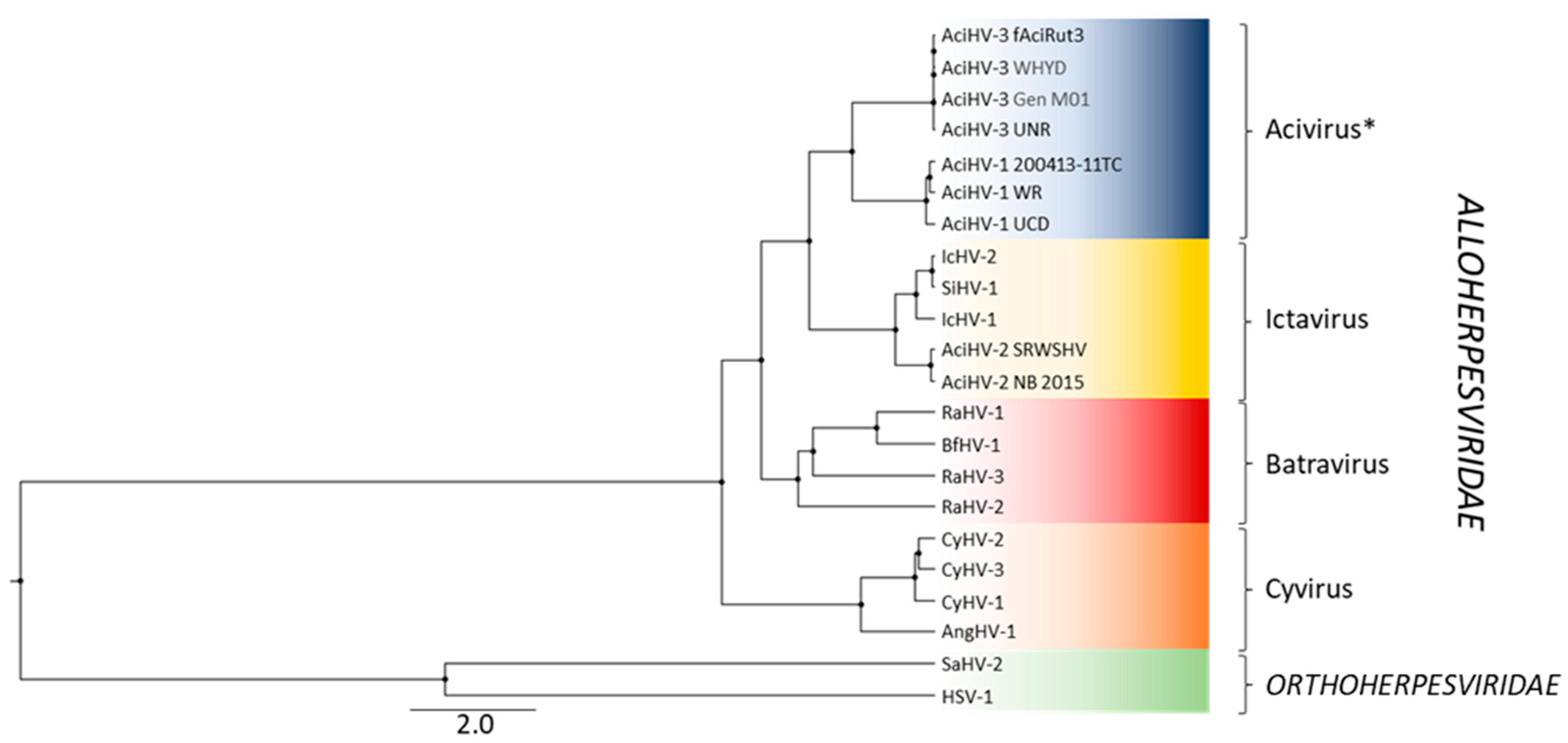
3.4. AciHV-3 Is a New Species and Member of a New Alloherpesvirus Genus
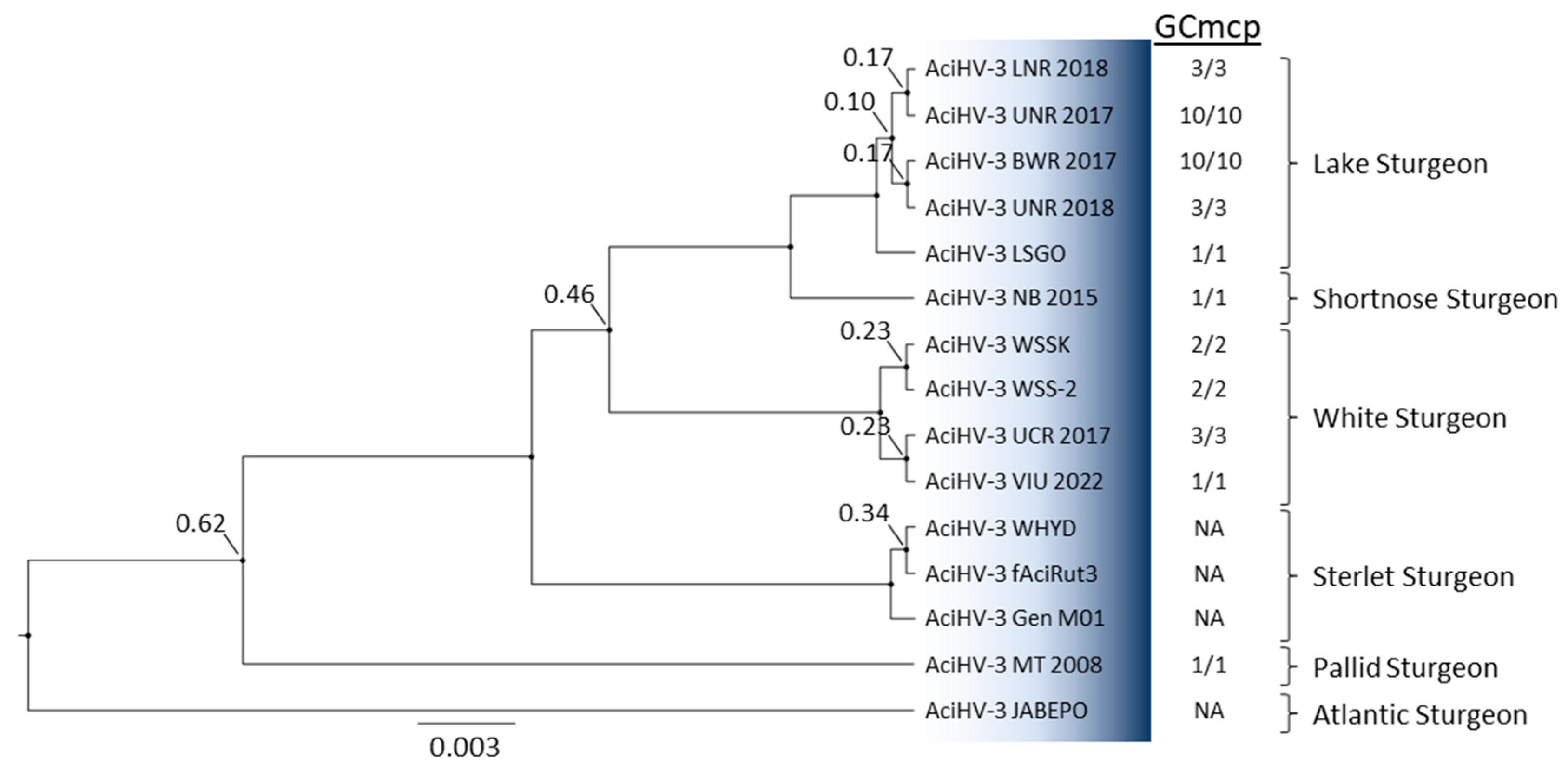
3.5. AciHV-3 Is Present in Other Sturgeon Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, D.; Li, Y.; Zhao, Q.; Zhao, L.; Ludwig, A.; Peng, Z. Highly resolved phylogenetic relationships within order Acipenseriformes according to novel nuclear markers. Genes 2019, 10, 38. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, N.; Liu, Z.; Chen, Q.; Li, Y. Phylogenetic perspective on the relationships and evolutionary history of the Acipenseriformes. Genomics 2020, 112, 3511–3517. [Google Scholar] [CrossRef]
- Shedko, S.V. Molecular dating of phylogeny of sturgeons (Acipenseridae) based on total evidence analysis. Russ. J. Genet. 2022, 58, 718–729. [Google Scholar] [CrossRef]
- Ruban, G.I. Modern concepts on the origin and distribution of Acipenserids (Acipenseridae). J. Ichthyol. 2023, 63, 206–215. [Google Scholar] [CrossRef]
- COSEWIC. COSEWIC Assessment and Status Report on the Lake Sturgeon Acipenser fulvescens, Western Hudson Bay Populations, Saskatchewan-Nelson River Populations, Southern Hudson Bay-James Bay Populations and Great Lakes-Upper St. Lawrence Populations in Canada; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2017; 153p. [Google Scholar]
- Osborne, M.J.; Dowling, T.E.; Scribner, K.T.; Turner, T.E. Wild at heart: Programs to diminish negative ecological and evolutionary effects of conservation hatcheries. Biol. Conserv. 2020, 251, 108768. [Google Scholar] [CrossRef]
- Hedrick, R.P.; Groff, J.M.; McDowell, T.S. Isolation of an epitheliotropic herpesvirus from white sturgeon (Acipenser transmontanus). Dis. Aquat. Org. 1991, 11, 49–56. [Google Scholar] [CrossRef]
- Watson, L.R.; Yun, S.C.; Groff, J.M.; Hedrick, R.P. Characteristics and pathogenicity of a novel herpesvirus isolated from adult and subadult white sturgeon Acipenser transmontanus. Dis. Aquat. Org. 1995, 22, 199–210. [Google Scholar] [CrossRef]
- Doszpoly, A.; Kovacs, E.R.; Bovo, G.; LaPatra, S.E.; Harrach, B.; Benko, M. Molecular confirmation of a new herpesvirus from catfish (Ameiurus melas) by testing the performance of a novel PCR method, designed to target the DNA polymerase gene of alloherpesviruses. Arch. Virol. 2008, 153, 2123–2127. [Google Scholar] [CrossRef]
- Kurobe, T.; Kelley, G.O.; Waltzek, T.B.; Hedrick, R.P. Revised phylogenetic relationships among herpesviruses isolated from sturgeons. J. Aquat. Anim. Health 2008, 20, 96–102. [Google Scholar] [CrossRef]
- LaPatra, S.E.; Groff, J.M.; Keith, I.; Hogans, W.E.; Groman, D. Iridoviral infection associated with disease in cultured shortnose sturgeon, Acipenser brevirostrum (L.), from the Atlantic coast of Canada. J. Fish. Dis. 2014, 37, 141–147. [Google Scholar] [CrossRef]
- Shchelkunov, I.S.; Shchelkunova, T.I.; Shchelkunov, A.I.; Kolbassova, Y.P.; Didenko, L.V.; Bykovsky, A.P. First detection of a viral agent causing disease in farmed sturgeon in Russia. Dis. Aquatic Org. 2009, 86, 193–203. [Google Scholar] [CrossRef]
- Doszpoly, A.; Shchelkunov, I.S. Partial genome analysis of Siberian Sturgeon alloherpesvirus suggests its close relation to AciHV-2–Short communication. Acta Vet. Hung. 2010, 58, 269–274. [Google Scholar] [CrossRef]
- Doszpoly, A.; Kalabekov, I.M.; Breyta, R.; Shchelkunov, I.S. Isolation and characterization of an atypical Siberian Sturgeon herpesvirus strain in Russia: Novel North American Acipenserid herpesvirus 2 strain in Europe? J. Fish. Dis. 2017, 40, 1363–1372. [Google Scholar] [CrossRef]
- Johnston, A.E.; Shavalier, M.A.; Scribner, K.T.; Soto, E.; Griffin, M.J.; Waldbieser, G.C.; Richardson, B.M.; Winters, A.D.; Yun, S.; Baker, E.A.; et al. First isolation of a herpesvirus (Family Alloherpesviridae) from Great Lakes Lake Sturgeon (Acipenser fulvescens). Animals 2022, 12, 3230. [Google Scholar] [CrossRef]
- Walker, L.; Subramaniam, K.; Viadanna, P.H.O.; Vann, J.A.; Marcquenski, S.; Godard, D.; Kieran, E.; Frasca, S., Jr.; Popov, V.L.; Kerr, K.; et al. Characterization of an alloherpesvirus from wild lake sturgeon Acipenser fulvescens in Wisconsin (USA). Dis. Aquat. Org. 2022, 14, 83–96. [Google Scholar] [CrossRef]
- Kelley, G.O.; Waltzek, T.B.; McDowell, T.S.; Yun, S.C.; LaPatra, S.E.; Hedrick, R.P. Genetic relationships among herpes-like viruses isolated from sturgeon. J. Aquat. Animal Health 2005, 17, 297–303. [Google Scholar] [CrossRef]
- Davison, A.J. Evolution of the herpesviruses. Vet. Micro. 2002, 86, 69–88. [Google Scholar] [CrossRef]
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef]
- Doszpoly, A.; Somogyi, V.; LaPatra, S.E.; Benko, M. Partial genome characterization of acipenserid herpesvirus 2: Taxonomical proposal for the demarcation of three subfamilies in Alloherpesviridae. Arch. Virol. 2011, 156, 2291–2296. [Google Scholar] [CrossRef]
- ICTV (International Committee on Taxonomy of Viruses). Available online: https://ictv.global/taxonomy (accessed on 10 June 2023).
- Clouthier, S.C.; Freshwater Institute, Winnipeg, MB, Canada. Personal communication, 2023.
- Davison, A.J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R.P.; Waltzek, T.B. Comparative genomics of carp herpesviruses. J. Virol. 2013, 87, 2908–2922. [Google Scholar] [CrossRef]
- Hanson, L.; Dishon, A.; Kotler, M. Herpesviruses that infect fish. Viruses 2011, 3, 2160–2191. [Google Scholar] [CrossRef] [PubMed]
- Hanson, L.; Doszpoly, A.; van Beurden, S.J.; Viadanna, P.; Waltzek, T. Alloherpesviruses of fish. In Aquaculture Virology; Kibenge, F.S.B., Godoy, M.G., Eds.; Academic Press: London, UK, 2016; pp. 153–172. [Google Scholar]
- Georgiadis, M.P.; Hedrick, R.P.; Johnson, W.O.; Yun, S.; Garner, I.A. Risk factors for outbreaks of disease attributable to white sturgeon iridovirus and white sturgeon herpesvirus-2 at a commercial sturgeon farm. Am. J. Vet. Res. 2000, 61, 1232–1240. [Google Scholar] [CrossRef]
- Cohen, J.I. Herpesvirus latency. J. Clin. Investig. 2020, 130, 3361–3369. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, R.P.; McDowell, T.S.; Rosemark, R.; Aronstein, D.; Lannan, C.N. Two cell lines from White Sturgeon. Trans. Am. Fish. Soc. 1991, 120, 528–534. [Google Scholar] [CrossRef]
- Clouthier, S.C.; VanWalleghem, E.; Copeland, S.; Klassen, C.; Hobbs, G.; Nielsen, O.; Anderson, E.D. A new species of nucleo-cytoplasmic large DNA virus (NCLDV) associated with mortalities in Manitoba Lake Sturgeon Acipenser fulvescens. Dis. Aquat. Org. 2013, 102, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Hanson, L.A.; Rudis, M.R.; Vasquez-Lee, M.; Montgomery, R.D. A broadly applicable method to characterize large DNA viruses and adenoviruses based on the DNA polymerase gene. Virol. J. 2006, 3, 28. [Google Scholar] [CrossRef]
- Macdonald, D. Nelson River Sturgeon Studies 1993–1997: Five Year Report to the Nelson River Sturgeon Co-Management Board; Nelson River Sturgeon Co-Management Board: Thompson, MB, Canada, 1998. [Google Scholar]
- Keeyask Hydropower Limited Partnership. Keeyask Generation Project Fisheries Offsetting and Mitigation Plan; Keeyask Hydropower Limited Partnership: Winnipeg, MB, Canada, 2015. [Google Scholar]
- Nelson, P.A.; Gosselin, T.; McDougall, C.A.; Bernatchez, L. In-stream population structuring of Lake Sturgeon in northern Manitoba, Canada. Fish. Manag. Ecol. 2023, 30, 406–422. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10.6.1; Environmental Systems Research Institute: Redlands, CA, USA, 2016. [Google Scholar]
- NRC (Government of Canada, Natural Resources Canada)–CANVec Series (Internet). Available online: https://open.canada.ca/data/en/dataset/9d96e8c9-22fe-4ad2-b5e8-94a6991b744b (accessed on 10 June 2023).
- BD Diagnostics. Difco & BBL Manual. Manual of Microbiological Culture Media, 2nd ed.; Becton, Dickinson and Company: Sparks, MD, USA, 2009. [Google Scholar]
- Gephart, P.; Murray, R.G.E.; Costilow, R.N.; Nester, E.W.; Wood, W.A.; Krieg, N.R.; Phillips, G.B. Manual of Methods for General Bacteriology; ASM Press: Washington, DC, USA, 1981. [Google Scholar]
- Yasutake, W.T.; Wales, J.H. Microscopic Anatomy of Salmonids: An Atlas; Resource Publication 150; United States Department of the Interior Fish and Wildlife Service: Washington, DC, USA, 1983.
- Du, K.; Stock, M.; Kneitz, S.; Klopp, C.; Woltering, J.M.; Adolfi, M.C.; Feron, R.; Prokopov, D.; Makunin, A.; Kichigin, I.; et al. The sterlet sturgeon genome sequence and the mechanisms of segmental rediploidization. Nat. Ecol. Evol. 2020, 4, 841–852. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bankevick, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Borodovsky, M.; Lomsadze, A. Gene identification in prokaryotic genomes, phages, metagenomes, and EST sequences with GeneMarkS suite. Curr. Protoc. Microbiol. 2014, 32, 1E.7.1–1E.7.17. [Google Scholar] [CrossRef] [PubMed]
- Tcherepanov, T.; Ehlers, A.; Upton, C. Genome Annotation Transfer Utility (GATU): Rapid annotation of viral genomes using a closely related reference genome. BMC Genom. 2006, 7, 150. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.M.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal Z version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Schwarz, G. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Yule, G.U. A mathematical theory of evolution, based on the conclusions of Dr. JC Willis. FRS. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1925, 213, 21–87. [Google Scholar]
- Gernhard, T. New analytic results for speciation times in neutral models. Bull. Math. Biol. 2008, 70, 1082–1097. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Henikoff, J.G. Amino acid substitution matrices from protein blocks. Proc. Natl. Acad. Sci. USA 1992, 89, 10915–10919. [Google Scholar] [CrossRef] [PubMed]
- Kingman, J.F.C. The coalescent. Stoch. Process Appl. 1982, 13, 235–248. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.7.1. 2018. Available online: https://github.com/beast-dev/tracer/releases/tag/v1.7.1 (accessed on 1 June 2023).
- Rambaut, A. FigTree, a Graphical Viewer of Phylogenetic Trees v1.4.4. 2018. Available online: https://github.com/rambaut/figtree/releases (accessed on 1 June 2023).
- Thiebaux, H.J.; Zwiers, F.W. The interpretation and estimation of effective sample size. J. Clim. Appl. Meteorol. 1984, 23, 800–811. [Google Scholar] [CrossRef]
- Kimura, T.; Yoshimizu, M.; Tanaka, M. Fish viruses: Tumor induction in Oncorhynchus keta by the herpesvirus. In Phyletic Approaches to Cancer; Dawe, C.J., Harshbarger, J.C., Kondo, S., Sugimura, T., Takayama, S., Eds.; Japanese Scientific Society Press: Tokyo, Japan, 1981; pp. 59–68. [Google Scholar]
- Yoshimizu, M.; Tanaka, M.; Kimura, T. Oncorhynchus masou virus (OMV): Incidence of tumor development among experimentally infected representative salmonid species. Fish Pathol. 1987, 22, 7–10. [Google Scholar] [CrossRef]
- Anders, K.; Yoshimizu, M. Role of viruses in the induction of skin tumours and tumour-like proliferations of fish. Dis. Aquat. Org. 1994, 19, 215–232. [Google Scholar] [CrossRef]
- Aswad, A.; Katzourakis, A. The first endogenous herpesvirus, identified in the tarsier genome and novel sequences from primate rhadinoviruses and lymphocryptoviruses. PLoS Genet. 2014, 10, e1004332. [Google Scholar] [CrossRef]
- Aswad, A.; Katzourakis, A. A novel viral lineage distantly related to herpesviruses discovered within fish genome sequence data. Virus Evol. 2017, 3, vex016. [Google Scholar] [CrossRef]
- Inoue, Y.; Saga, T.; Aikawa, T.; Kumagai, M.; Shimada, A.; Kawaguchi, Y.; Naruse, K.; Morishita, S.; Koga, A.; Takeda, H. Complete fusion of a transposon and herpesvirus created the Teratorn mobile element in medaka fish. Nat. Commun. 2017, 8, 551. [Google Scholar] [CrossRef]
- Kaufer, B.B.; Jarosinski, K.W.; Osterrieder, N. Herpesvirus telomeric repeats facilitate genomic integration into host telomeres and mobilization of viral DNA during reactivation. J. Exp. Med. 2011, 208, 605–615. [Google Scholar] [CrossRef]
- Delecluse, H.J.; Hammerschmidt, W. Status of Marek’s disease virus in established lymphoma cell lines: Herpesvirus integration is common. J. Virol. 1993, 67, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Delecluse, H.J.; Schuller, S.; Hammerschmidt, W. Latent Marek’s disease virus can be activated from its chromosomally integrated state in herpesvirustransformed lymphoma cells. EMBO J. 1993, 12, 3277–3286. [Google Scholar] [CrossRef]
- Arbuckle, J.H.; Medveczky, M.M.; Luka, J.; Hadley, S.H.; Luegmayr, A.; Ablashi, D.; Lund, T.C.; Tolar, J.; De Meirleir, K.; Montoya, J.G.; et al. The latent human herpesvirus-6A genome specifically integrates in telomeres of human chromosomes in vivo and in vitro. Proc. Natl. Acad. Sci. USA 2010, 107, 5563–5568. [Google Scholar] [CrossRef] [PubMed]
- Arbuckle, J.H.; Pantry, S.N.; Medveczky, M.M.; Prichett, J.; Loomis, K.S.; Ablashi, D.; Medveczky, P.G. Mapping the telomere integrated genome of human herpesvirus 6A and 6B. Virology 2013, 442, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Kelly, R.K.; Nielsen, O. Epidermal hyperplasias of Northern Pike (Esox lucius) associated with herpesvirus and C-type particles. Arch. Virol. 1983, 79, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Margenau, T.L.; Marcquenski, S.V.; Rasmussen, P.W.; Macconnell, E. Prevalence of Blue Spot Disease (Esocid Herpesvirus-1) on Northern Pike and Muskellunge in Wisconsin. J. Aquat. Anim. Health 1995, 7, 29–33. [Google Scholar] [CrossRef]
- Garver, K.A.; Leskisenoja, K.; Macrae, R.; Hawley, L.M.; Subramaniam, K.; Waltzek, T.B.; Richard, J.; Josefsson, C.; Valtonen, E.T. An alloherpesvirus infection of European perch Perca fluviatilis in Finland. Dis. Aquatic Org. 2018, 28, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Bowser, P.R.; Plumb, J.A. Growth rates of a new cell line from channel catfish ovary and channel catfish virus replication at different temperatures. Can. J. Fish. Aquat. Sci. 1980, 37, 871–873. [Google Scholar] [CrossRef]
- Wolf, K.; Darlington, R.W.; Taylor, W.G.; Quimby, M.C.; Nagabayashi, T. Herpesvirus salmonis: Characterization of a New Pathogen of Rainbow Trout. J. Virol. 1978, 27, 659–666. [Google Scholar] [CrossRef]
- Anders, K. A herpesvirus associated with an epizootic epidermal papillomatosis in European smelt (Osmerus eperlanus). In Viruses of Lower Vertebrates; Ahne, W., Kurstak, E., Eds.; Springer: Berlin, Germany, 1989; pp. 184–197. [Google Scholar]
- Sano, T.; Fukuda, H.; Furukawa, M. Herpesvirus cyprini: Biological and oncogenic properties. Fish Pathol. 1985, 20, 381–388. [Google Scholar] [CrossRef]
- Sano, T. Viral diseases of cultured fishes in Japan. Fish Pathol. 1976, 10, 221–226. [Google Scholar] [CrossRef]
- Kimura, T.; Yoshimizu, M.; Tanaka, M.; Sannohe, H. Studies on a new virus (OMV) from Oncorhynchus masou–I. Characteristics and pathogenicity. Fish Pathol. 1981, 15, 143–147. [Google Scholar] [CrossRef]
- McAllister, P.E.; Herman, R.L. Epizootic mortality in hatchery-reared lake trout Salvelinus namaycush caused by a putative virus possibly of the herpesvirus group. Dis. Aquat. Org. 1989, 6, 113–119. [Google Scholar] [CrossRef]
- Bradley, T.M.; Medina, D.J.; Chang, P.W.; McClain, J. Epizootic epitheliotropic disease of lake trout (Salvelinus namaycush): History and viral etiology. Dis. Aquat. Org. 1989, 7, 195–201. [Google Scholar] [CrossRef]
- Iida, Y.; Masumura, K.; Nakai, T.; Sorimachi, M.; Matsuda, H. A viral disease in larvae and juveniles of the Japanese Flounder Paralichthys olivaceus. J. Aquat. Anim. Health 1989, 1, 7–12. [Google Scholar] [CrossRef]
- Koonin, E.V.; Krupovic, M. The depths of virus exaptation. COVIRO 2018, 31, 1–8. [Google Scholar] [CrossRef]
- Shapira, Y.; Magen, Y.; Zak, T.; Kotler, M.; Hulata, G.; Levavi-Sivan, B. Differential resistance to koi herpes virus (KHV)/carp interstitial nephritis and gill necrosis virus (CNGV) among common carp (Cyprinus carpio L.) strains and crossbreds. Aquaculture 2005, 245, 1–11. [Google Scholar] [CrossRef]
- Dixon, P.F.; Joiner, C.L.; Way, K.; Reese, R.A.; Jeney, G.; Jeney, Z. Comparison of the resistance of selected families of common carp, Cyprinus carpio L.; to koi herpesvirus: Preliminary study. J. Fish. Dis. 2009, 32, 1035–1039. [Google Scholar] [CrossRef]
- Piackova, V.; Flajshans, M.; Pokorova, D.; Reschova, S.; Gela, D.; Cizek, A.; Vesely, T. Sensitivity of common carp, Cyprinus carpio L.; strains and crossbreeds reared in the Czech Republic to infection by cyprinid herpesvirus 3 (CyHV-3; KHV). J. Fish Dis. 2013, 36, 75–80. [Google Scholar] [CrossRef]
- Kane, M.; Golovkina, T. Common threads in persistent viral infections. J. Virol. 2010, 84, 4116–4123. [Google Scholar] [CrossRef] [PubMed]
- Rakus, K.; Ouyang, P.; Boutier, M.; Ronsmans, M.; Reschner, A.; Vancsok, C.; Jazowiecka-Rakus, J.; Vanderplasschen, A. Cyprinid Herpesvirus 3: An Interesting Virus for Applied and Fundamental Research. Vet. Res. 2013, 44, 85. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, P.; Rakus, K.; Boutier, M.; Reschner, A.; Leroy, B.; Ronsmans, M.; Fournier, G.; Scohy, S.; Costes, B.; Wattiez, R.; et al. The IL-10 homologue encoded by cyprinid herpesvirus 3 is essential neither for viral replication in vitro nor for virulence in vivo. Vet. Res. 2013, 44, 53. [Google Scholar] [CrossRef] [PubMed]
- Clouthier, S.; Caskenette, A.; VanWalleghem, E.; Schroeder, T.; Macdonald, D.; Anderson, E.D. Molecular phylogeny of sturgeon mimiviruses and Bayesian hierarchical modeling of their effect on wild Lake Sturgeon (Acipenser fulvescens) in Central Canada. Infect. Genet. Evol. 2020, 84, e104491. [Google Scholar] [CrossRef]
- McGeoch, D.J.; Rixon, F.J.; Davison, A.J. Topics in herpesvirus genomics and evolution. Virus Res. 2006, 117, 90–104. [Google Scholar] [CrossRef]
- Aiewsakun, P.; Katzourakis, A. Endogenous viruses: Connecting recent and ancient viral evolution. Virology 2015, 479–480, 26–37. [Google Scholar] [CrossRef]
- Aswad, A.; Aimola, G.; Wight, D.; Roychoudhury, P.; Zimmermann, C.; Hill, J.; Lassner, D.; Xie, H.; Huang, M.-L.; Parrish, N.F.; et al. Evolutionary history of endogenous human herpesvirus 6 reflects human migration out of Africa. Mol. Biol. Evol. 2020, 38, 96–107. [Google Scholar] [CrossRef]
- Booy, F.P.; Trus, B.L.; Davison, A.D.; Steven, A.C. The capsid architecture of channel catfish virus, an evolutionarily distant herpesvirus, is largely conserved in the absence of discernible sequence homology with herpes simplex virus. Virology 1996, 215, 134–141. [Google Scholar] [CrossRef]
- Bowman, B.R.; Baker, M.L.; Rixon, F.J.; Chiu, W.; Quiocho, F.A. Structure of the herpesvirus major capsid protein. EMBO J. 2003, 22, 757–765. [Google Scholar] [CrossRef]
- Baker, M.L.; Jiang, W.; Bowman, B.R.; Zhou, Z.H.; Quiocho, F.A.; Rixon, F.J.; Chiu, W. Architecture of the Herpes Simplex Virus major capsid protein derived from structural bioinformatics. J. Mol. Biol. 2003, 331, 447–456. [Google Scholar] [CrossRef]
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef]
- EUMOFA (European Market Observatory for Fisheries and Aquaculture Products). Sturgeon Meat and Other By-Products of Caviar. Production, Trade, and Consumption in and Outside the EU; Publications Office of the European Union: Luxembourg, 2023. [Google Scholar] [CrossRef]






| Virus Taxa/Genera 1 | Acivirus 2 | Ictavirus 3 | Batravirus | Cyvirus | |
|---|---|---|---|---|---|
| AciHV-3 | AciHV-1 | ||||
| AciHV-3 | 99–100 (99.6–100) | - | - | - | - |
| AciHV-1 | 31–32 (48) | 85–90 (91–94) | - | - | - |
| Ictavirus | 24–25 (42–43) | 23–26 (41–45) | 51–90 (69–97) | - | - |
| Batravirus | 21–25 (37–40) | 20–23 (36–41) | 17–22 (35–40) | 24–39 (41–58) | - |
| Cyvirus | 18–19 (32–34) | 17–18 (31–33) | 14–18 (28–34) | 30–31 (30–32) | 29–76 (47–85) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clouthier, S.; Tomczyk, M.; Schroeder, T.; Klassen, C.; Dufresne, A.; Emmenegger, E.; Nalpathamkalam, T.; Wang, Z.; Thiruvahindrapuram, B. A New Sturgeon Herpesvirus from Juvenile Lake Sturgeon Acipenser fulvescens Displaying Epithelial Skin Lesions. Pathogens 2023, 12, 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12091115
Clouthier S, Tomczyk M, Schroeder T, Klassen C, Dufresne A, Emmenegger E, Nalpathamkalam T, Wang Z, Thiruvahindrapuram B. A New Sturgeon Herpesvirus from Juvenile Lake Sturgeon Acipenser fulvescens Displaying Epithelial Skin Lesions. Pathogens. 2023; 12(9):1115. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12091115
Chicago/Turabian StyleClouthier, Sharon, Marek Tomczyk, Tamara Schroeder, Cheryl Klassen, André Dufresne, Eveline Emmenegger, Thomas Nalpathamkalam, Zhuozhi Wang, and Bhooma Thiruvahindrapuram. 2023. "A New Sturgeon Herpesvirus from Juvenile Lake Sturgeon Acipenser fulvescens Displaying Epithelial Skin Lesions" Pathogens 12, no. 9: 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens12091115