Effects of Specific Electric Field Stimulation on the Release and Activity of Secreted Acid Phosphatases from Leishmania tarentolae and Implications for Therapy

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Leishmaniasis and Established Treatments
1.2. L. tarentolae Lifecycle and Secreted Acid Phosphatases
1.3. Electric Fields and L. tarentolae
2. Materials and Methods
2.1. In Vitro L. tarentolae Culture and Cell Viability Determination by MTT Analysis
2.2. Microscopy of L. tarentolae
2.3. L. tarentolae Acid Phosphatase Enzyme Preparation
2.4. Secreted Acid Phosphatase Enzyme Assay and Kinetic Assays
2.5. Secreted Acid Phosphatase Enzyme Kinetic Assays with and without Pre-incubation with a Glycosidase
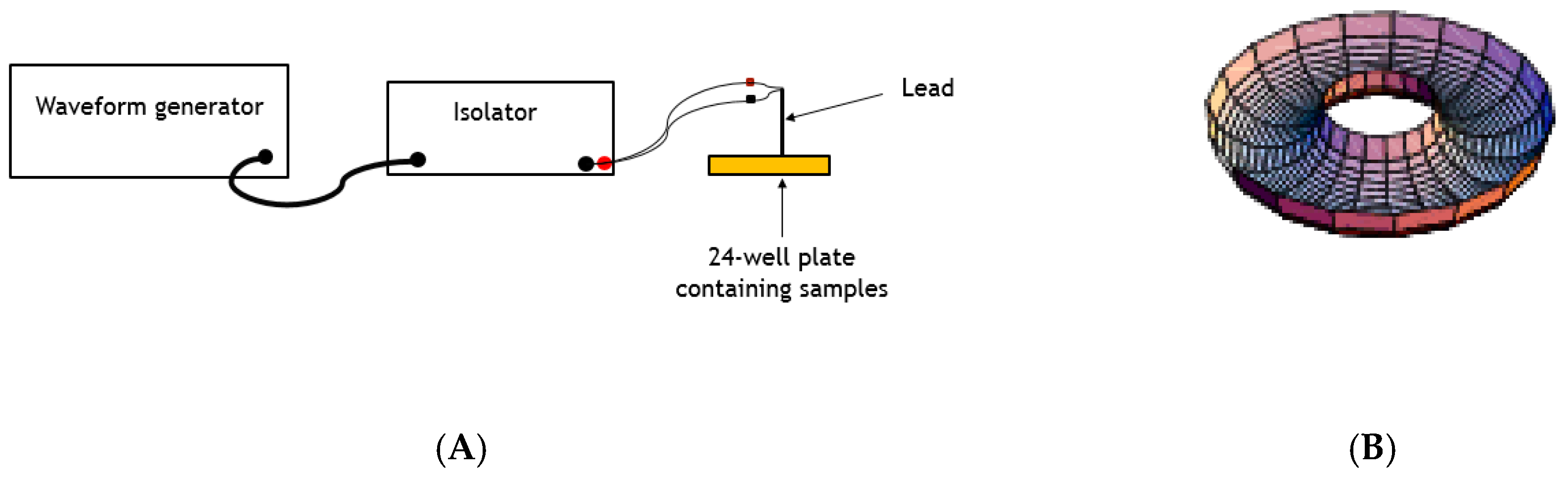
2.6. Testing the Effects of Electric Fields on L. tarentolae Acid Phosphatase Activity and Secretion of Acid Phosphatase
2.6.1. Effect on Secreted Enzyme Activity (Method 1)
2.6.2. Effect on Enzyme Secretion (Method 2)
3. Statistical Analysis
4. Results
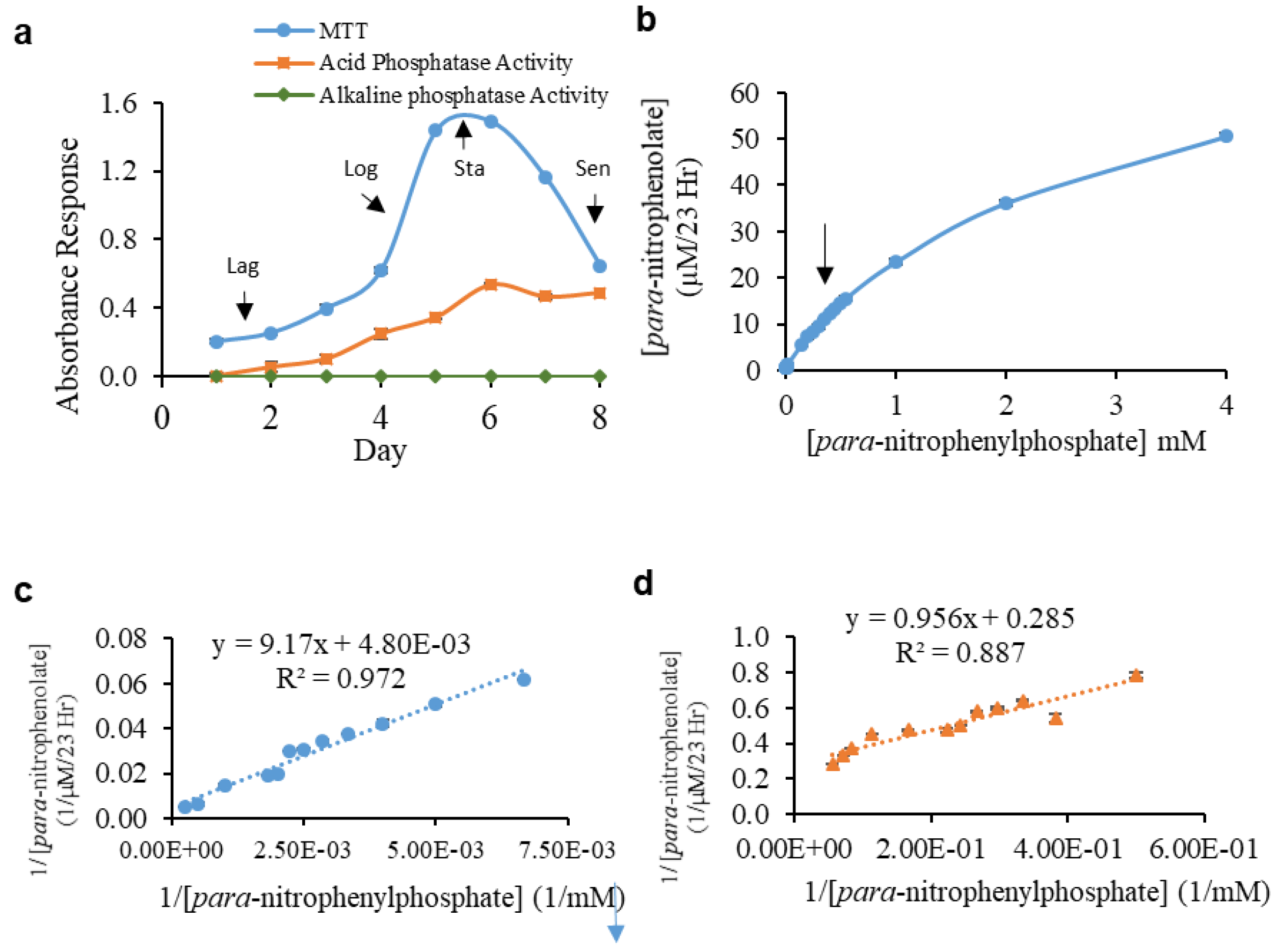
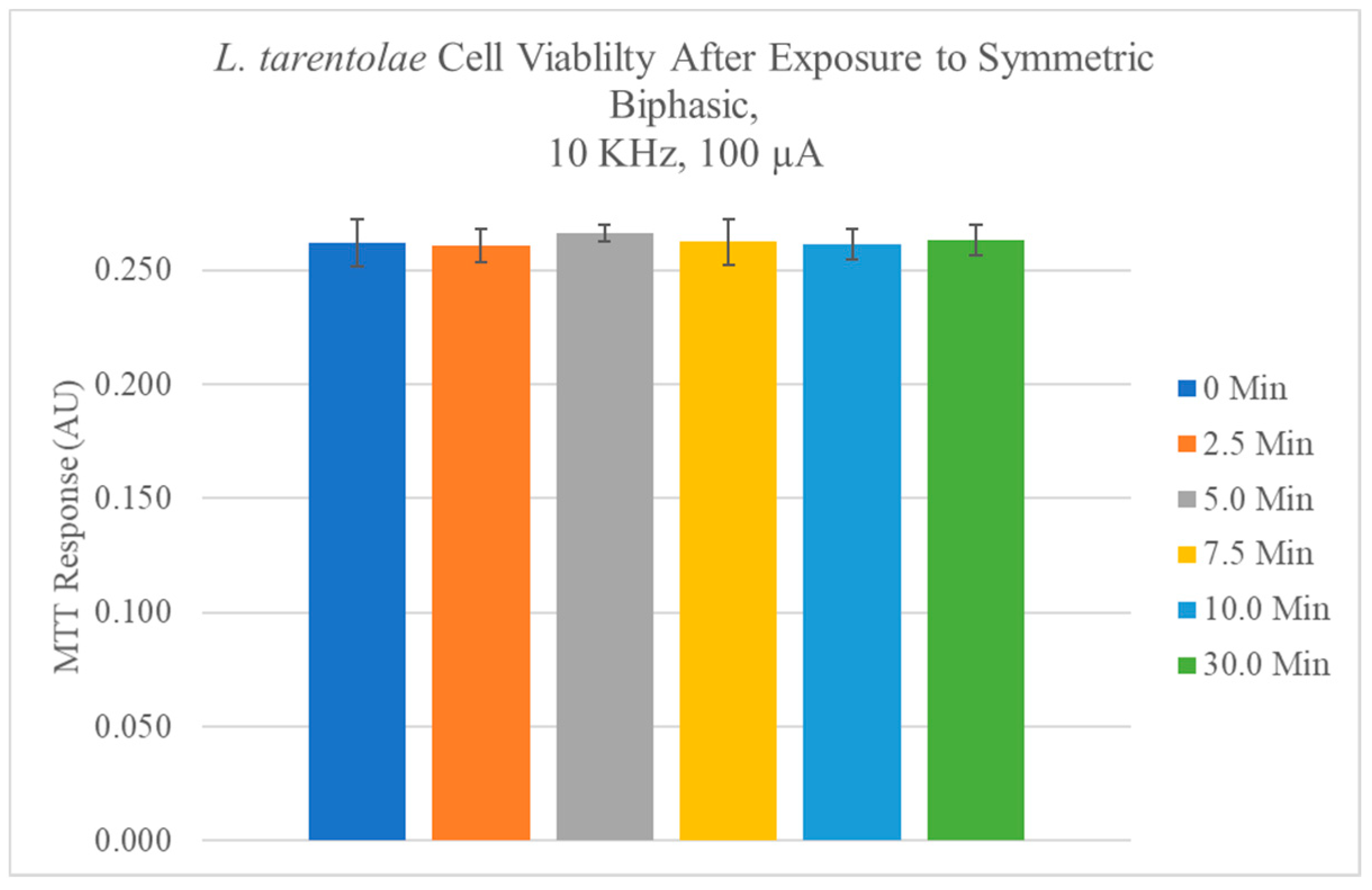
4.1. L. tarentolae Cell Viability with and without the Application of Electric Fields
4.2. Microscopy of L. tarentolae with and without the Application of Electric Fields
4.3. Enzyme Kinetics with and without Glycosidase Incubation
4.4. Effect of Electric Fields on Secreted and Pelleted Acid Phosphatase Activity (Method 1)
4.5. Effect of Electric Fields on SAP Secretion (Method 2)
5. Discussion
5.1. Phenotypic and Viability Effects of Electric Fields on L. tarentolae Promastigotes
5.2. SAP Enzyme Kinetics and the Effect of Glycosylation
5.3. Electrical Stimulation Wave Form Comparisons
6. Conclusions
Potential for Synergistic Therapeutic Treatment for Leishmaniasis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Steverding, D. The history of leishmaniasis. Parasites Vectors 2017, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Epidemiology & Risk Factors Leishmaniasis. Available online: https://www.cdc.gov/parasites/leishmaniasis/epi.html (accessed on 20 December 2016).
- World Health Organization. WHO Model Lists of Essential Medicines. Available online: http://www.who.int/medicines/publications/essentialmedicines/en/ (accessed on 15 August 2017).
- Centers for Disease Control and Prevention. Leishmaniasis Professionals. Available online: https://www.cdc.gov/parasites/leishmaniasis/health_professionals/index.html#tx (accessed on 20 December 2016).
- World Health Organization. WHO Leishmaniasis Treatment Cost. Available online: http://www.who.int/leishmaniasis/research/978_92_4_12_949_6_Annex6.pdf (accessed on 21 June 2017).
- Taylor, V.M.; Munoz, D.L.; Cedeno, D.L.; Velez, I.V.; Jones, M.A.; Robledo, S.M. Leishmania tarentolae: Utility as an in vitro model for screening of antileishmanial agents. Exp. Parasitol. 2010, 126, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Mojtahedi, Z.; Clos, J.; Kamali-Sarvestani, E. Leishmania major: Identification of developmentally regulated proteins in procyclic and metacyclic promastigotes. Exp. Parasitol. 2008, 119, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Vannier-Stantos, M.A.; Martiny, A.; Meyer-Fernandes, J.R.; De Souza, W. Leishmanial protein kinase C modulates host cell infection via secreated acid phosphatase. Eur. J. Cell Biol. 1995, 67, 112–119. [Google Scholar]
- Shakarian, A.M.; Ellisa, S.L.; Mallinsonac, D.J.; Olafsonb, R.W.; Dwyer, D.M. Two tandemly arrayed genes encode the (histidine) secretory acid phosphatases of Leishmania donovani. Gene 1997, 196, 127–137. [Google Scholar] [CrossRef]
- Fernandes, A.C.; Soares, D.C.; Saraiva, E.M.; Meyer-Fernandez, J.R.; Souto-Padron, T. Different secreted phosphatase activites in Leishmania amazonensis. FEMS Microbiol. Lett. 2013, 340, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Isnard, A.; Shio, M.T.; Oliver, M. Impact of Leishmania metalloprotease GP63 on macrophage signaling. Front. Cell. Infect. Microbiol. 2012, 2, 72. [Google Scholar] [CrossRef] [PubMed]
- Mendez, R.S.; Dorsey, B.M.; McLauchlan, C.C.; Beio, M.; Turner, T.L.; Nguyen, V.H.; Su, A.; Beynon, W.; Friesen, J.A.; Jones, M.A. Vanadium complexes are in vitro inhibitors of leishmania secreted acid phosphatases. Int. J. Chem. 2014, 6, 35–49. [Google Scholar] [CrossRef]
- Glew, R.H.; Saha, A.K.; Das, S.; Remaley, A.T. Biochemistry of the Leishmania species. Microbiol. Rev. 1988, 52, 412–432. [Google Scholar] [PubMed]
- Baghaei, M.; Baghaei, M. Characterization of acid phosphatase in the promastigotes of three isolates of Leishmania major. Iran. J. Med. Sci. 2003, 28, 1–8. [Google Scholar]
- Vannier-Stantos, M.A.; Martiny, A.; De Souza, W. Cell biology of Leishmania spp.: Invading and evading. Curr. Pharm. Des. 2002, 8, 297–318. [Google Scholar] [CrossRef]
- Raymond, F.; Boisvert, S.; Roy, G.; Ritt, J.-F.; Légaré, D.; Isnard, A.; Stanke, M.; Olivier, M.; Tremblay, M.J.; Papadopoulou, B.; et al. Genome sequencing of the lizard parasite Leishmania tarentolae reveals loss of genes associated to the intracellular stage of human pathogenic species. Nucleic Acids Res. 2012, 40, 1131–1147. [Google Scholar] [CrossRef] [PubMed]
- Navabi, A.; Soleimanifard, S. Enzymatic characterization of acid phosphatase in the logarithmic and stationary phase of Leishmania major promastigotes. Shiraz E Med. J. 2015, 16, e26246. [Google Scholar] [CrossRef]
- Lippert, D.N.; Dwyer, D.W.; Li, F.; Olafson, R.W. Phosphoglycosylation of a secreted acid phosphatase from Leishmania donovani. Glycobiology 1999, 9, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Holden, K.R. Biological Effects of Pulsed Electromagnetic Field (PEMF) Therapy. Available online: http://www.ondamed.net/us/biological-effects-of-pulsed-electromagnetic-field-pemf-therapy (accessed on 2 February 2017).
- Giladi, M.; Porat, Y.; Blatt, A.; Shmueli, E.; Wasserman, Y.; Kirson, E.D.; Palti, Y. Microbial growth inhibition by alternating electric fields in mice with Pseudomonas aeruginosa lung infection. Antimicrob. Agents Chemother. 2010, 54, 3212–3218. [Google Scholar] [CrossRef] [PubMed]
- Hejazi, H.; Eslami, G.; Dalimi, A. The parasiticidal effect of electricity on Leishmania major, both in vitro and in vivo. Ann. Trop. Med. Parasit. 1972, 98, 37–42. [Google Scholar]
- Morgenthaler, J.B.; Peters, S.J.; Cedeno, D.L.; Constantino, M.H.; Edwards, K.A.; Kamowski, E.M.; Jones, M.A. Carbaporphyrin ketals as potential agents for a new photodynamic therapy treatment of leishmaniasis. Bioorg. Med. Chem. 2008, 16, 7033–7038. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [PubMed]
- Lovelace, J.K.; Gottlieb, M. Comparison of extracellular acid phosphatases from various isolates of Leishmania. Am. J. Trop. Med. Hyg. 1986, 35, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Shakarian, A.M.; Dwyer, D.M. Structurally conserved soluble acid phosphatases are synthesized and released by Leishmania major promastigotes. Exp. Parasitol. 2000, 95, 79–84. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorsey, B.M.; Cass, C.L.; Cedeño, D.L.; Vallejo, R.; Jones, M.A. Effects of Specific Electric Field Stimulation on the Release and Activity of Secreted Acid Phosphatases from Leishmania tarentolae and Implications for Therapy. Pathogens 2018, 7, 77. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens7040077
Dorsey BM, Cass CL, Cedeño DL, Vallejo R, Jones MA. Effects of Specific Electric Field Stimulation on the Release and Activity of Secreted Acid Phosphatases from Leishmania tarentolae and Implications for Therapy. Pathogens. 2018; 7(4):77. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens7040077
Chicago/Turabian StyleDorsey, Benjamin M., Cynthia L. Cass, David L. Cedeño, Ricardo Vallejo, and Marjorie A. Jones. 2018. "Effects of Specific Electric Field Stimulation on the Release and Activity of Secreted Acid Phosphatases from Leishmania tarentolae and Implications for Therapy" Pathogens 7, no. 4: 77. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens7040077