Identification of Differentially Expressed Genes in BALB/c Mouse Liver upon Primary Infection with DENV1 and Sequential Heterologous Infection with DENV2


Abstract
:1. Introduction
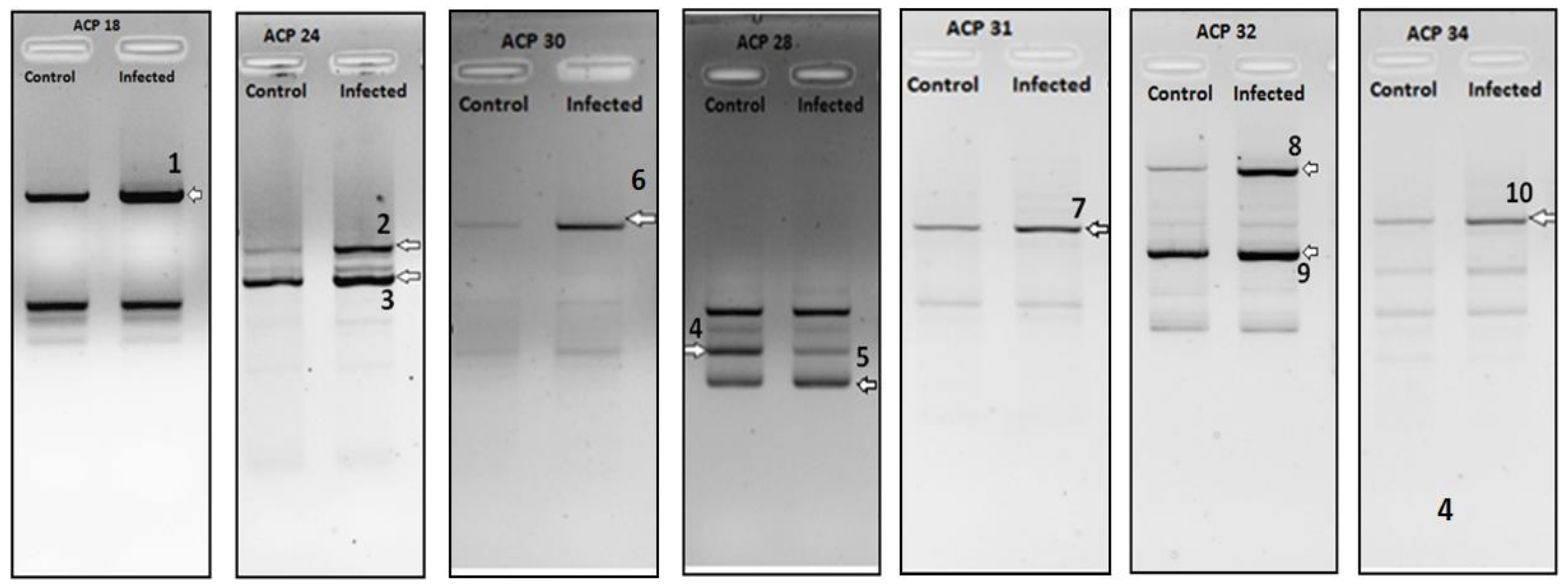
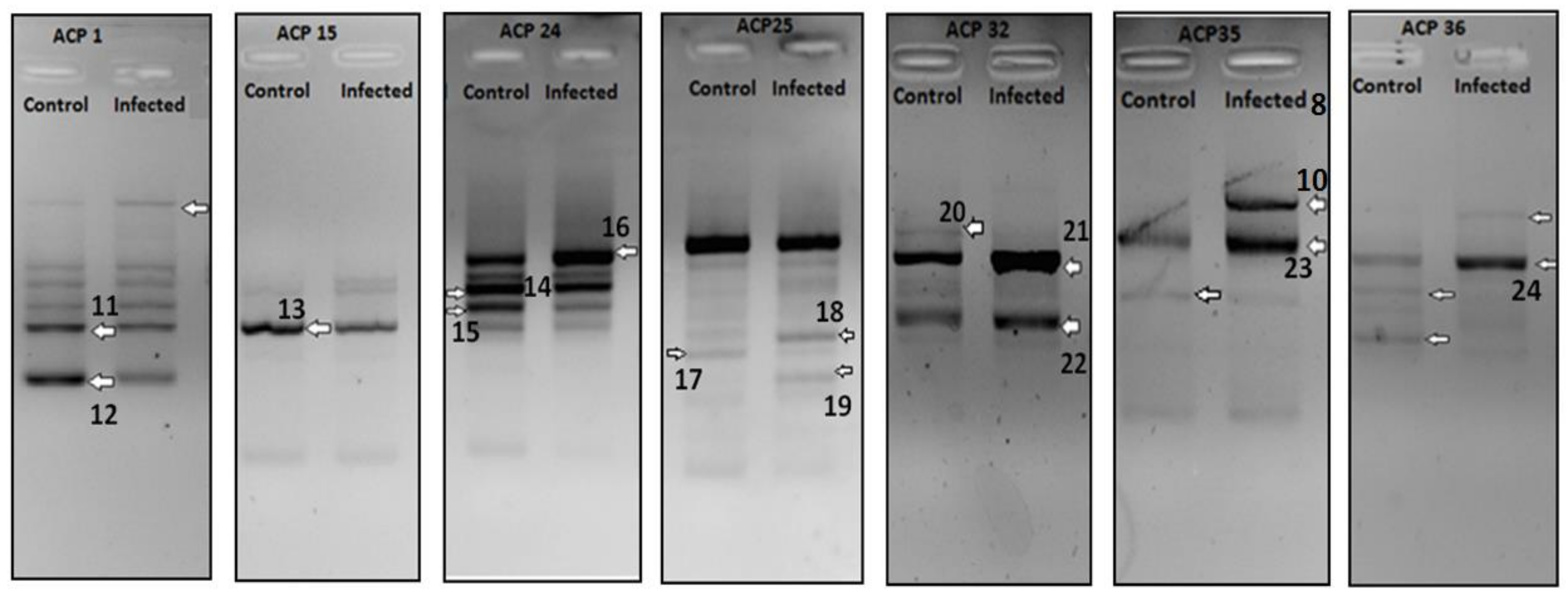
2. Results
3. Discussion
4. Materials and Methods
4.1. Viruses
4.2. Animal Work
4.3. mRNA Differential Display PCR
4.4. Quantitative Real-Time PCR
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McBride, W.J.; Bielefeldt-Ohmann, H. Dengue viral infections; pathogenesis and epidemiology. Microbe. Infect. 2000, 2, 1041–1050. [Google Scholar] [CrossRef]
- Tham, H.W.; Balasubramaniam, V.; Chew, M.F.; Ahmad, H.; Hassan, S.S. Protein-protein interactions between A. aegypti midgut and dengue virus 2: Two-hybrid screens using the midgut cDNA library. J. Infect. Dev. Ctries. 2015, 9, 1338–1349. [Google Scholar] [CrossRef] [PubMed]
- Murrell, S.; Wu, S.C.; Butler, M. Review of dengue virus and the development of a vaccine. Biotechnol. Adv. 2011, 29, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Rothman, A.L. Dengue: Defining protective versus pathologic immunity. J. Clin. Invest. 2004, 113, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Rothman, A.L.; Ennis, F.A. Immunopathogenesis of dengue hemorrhagic fever. Virology 1999, 257, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Liu, C.C.; Chuang, J.I.; Lei, H.Y.; Yeh, T.M.; Lin, Y.S.; Huang, Y.H.; Liu, H.S. Involvement of oxidative stress, NF-IL-6 and RANTES expression in dengue-2 virus infected human liver cells. Virology 2000, 276, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.Y.; Yeh, T.M.; Liu, H.S.; Lin, Y.S.; Chen, S.H.; Liu, C.C. Immunopathogenesis of dengue virus infection. J. Biomed. Sci. 2001, 8, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Yauch, L.E.; Shresta, S. Mouse models of dengue virus infection and disease. Antiviral Res. 2008, 80, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneviratne, S.L.; Malavige, G.N.; de Silva, H.J. Pathogenesis of liver involvement during dengue viral infections. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Bhamarapravati, N. Hemostatic defects in dengue hemorrhagic fever. Rev. Infect. Dis. 1989, 11, 826–829. [Google Scholar] [CrossRef]
- Subramanian, V.; Shenoy, S.; Joseph, A.J. Dengue hemorrhagic fever and fulminant hepatic failure. Dig. Dis. Sci. 2005, 50, 1146–1147. [Google Scholar] [CrossRef] [PubMed]
- Pancharoen, C.; Rungsarannont, A.; Thisyakorn, U. Hepatic dysfunction in dengue patients with various severity. J. Med. Assoc. Thail. 2002, 85, 298–301. [Google Scholar]
- Smith, D.R.; Khakpoor, A. Involvement of the liver in dengue infections. Dengue Bull. 2009, 33, 75–86. [Google Scholar]
- Kuo, C.H.; Tai, D.I.; Chang-Chien, C.S.; Lan, C.K.; Chiou, S.S.; Liaw, Y.F. Liver biochemical tests and dengue fever. Am. J. Trop. Med. Hyg. 1992, 47, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, O.; Hongsiriwon, S.; Bowonwatanuwong, C.; Chotivanich, K.; Sukthana, Y.; Pukrittayakamee, S. Risk factors and clinical features associated with severe dengue infection in adults and children during the 2001 epidemic in Chonburi, Thailand. Trop. Med. Int. Health 2004, 9, 1022–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekkapongpisit, E.; Wannatung, T.; Susantad, T.; Triwitayakorn, K.; Smith, D.R. cDNA-AFLP analysis of differential gene expression in human hepatoma (HepG2) cells upon dengue virus infection. J. Med. Virol. 2007, 79, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Loke, P.; Hammond, S.N.; Leung, J.M.; Kim, C.C.; Batra, S.; Rocha, C.; Balmaseda, A.; Harris, E. Gene expression patterns of dengue virus-infected children from Nicaragua reveal a distinct signature of increased metabolism. PLoS Negl. Trop. Dis. 2010, 4, e710. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.C.; Friedmann, T.; Driever, W.; Burrascano, M.; Yee, J.K. Vesicular stomatitis virus G glycoprotein pseudotyped retroviral vectors: Concentration to very high titer and efficient gene transfer into mammalian and non-mammalian cells. Proc. Natl. Acad. Sci. USA 1993, 90, 8033–8037. [Google Scholar] [CrossRef] [PubMed]
- Hierholzer, J.; Killington, R. Virus isolation and quantitation. In Virology Methods Manual; Hillar, O.K., Brian, W.J.M., Eds.; Academic Press: Cambridge, MA, USA, 1996; pp. 25–46. [Google Scholar]
- Balasubramaniam, V.R.M.T.; Wai, T.H.; Omar, A.R.; Othman, I.; Hassan, S.S. Cellular transcripts of chicken brain tissue in response to H5N1 and Newcastle disease virus. Virol. J. 2012, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, V.R.M.T.; Hassan, S.S.; Omar, A.R.; Mohamed, M.; Noor, S.M.; Mohamed, R.; Othman, I. Cellular transcripts regulated during infections with Highly Pathogenic H5N1 Avian Influenza virus in 3 host systems. Virol. J. 2011, 8, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.T.; Kim, Y.J.; Kim, S.H.; Kwak, C.I.; Gu, Y.Y.; Chun, J.Y. Annealing control primer system for improving specificity of PCR amplification. BioTechniques 2003, 35, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DEG No. | Regulation (Infected) | Identified mRNA | Max Identity | Accession |
|---|---|---|---|---|
| 1 | Down | Mus musculus kelch-like 25 (Drosophila), mRNA (cDNA clone IMAGE:4159269) | 99% | BC022600.1 |
| 2 | Down | Mus musculus histidine ammonia lyase, mRNA (cDNA clone MGC:67748 IMAGE:4211545), complete cds | 99% | BC057637.1 |
| 3 | Down | Mus musculus cDNA sequence BC024137, mRNA (cDNA clone IMAGE:5136153), partial cds | 99% | BC024137.1 |
| 4 | Down | Mus musculus SEC62 homolog (S. cerevisiae) (Sec62), mRNA | 99% | NM_027016.2 |
| 5 | Down | Mus musculus prefoldin 5 (Pfdn5), mRNA | 99% | NM_027044.3 |
| 6 | Up | Mus musculus translocase of outer mitochondrial membrane 6 homolog (yeast), mRNA (cDNA clone MGC:46796 IMAGE:5007624), complete cds | 98% | BC037589.1 |
| 7 | Down | Mus musculus C-reactive protein, pentraxin-related, mRNA (cDNA clone MGC:18634 IMAGE:4195658), complete cds | 100% | BC011124.1 |
| 8 | Up | Mus musculus S-adenosylhomocysteine hydrolase (Ahcy), mRNA | 99% | NM_016661.3 |
| 9 | Up | Mus musculus alpha 1 microglobulin/bikunin, mRNA (cDNA clone MGC:14070 IMAGE:4193922), complete cds | 99% | BC021660.1 |
| 10 | Down | Mus musculus mitochondrial DNA from Lewis lung carcinoma, complete sequence | 88% | AP013054.1 |
| DEG No. | Regulation (Infected) | Identified mRNA | Max Identity | Accession |
|---|---|---|---|---|
| 11 | Down | Mus musculus fibronectin 1, mRNA (cDNA clone IMAGE:4985138), partial cds | 99% | BC036167.1 |
| 12 | Down | Mus musculus mitochondrial DNA from Lewis lung carcinoma, complete sequence | 96% | AP013054.1 |
| 13 | Down | Mus musculus mRNA for serum albumin | 99% | AJ011413.1 |
| 14 | Down | Mus musculus interferon induced transmembrane protein 1 (Ifitm1), transcript variant 2, mRNA | 94% | NM_001112715.1 |
| 15 | Down | Mus musculus ribosomal protein L12 (Rpl12), mRNA | 99% | NM_009076.3 |
| 16 | Up | Mus musculus MACRO domain containing 1, mRNA | 78% | BC008653.1 |
| (cDNA clone MGC:11843 IMAGE:3597023), complete cds | ||||
| 17 | Down | Mus musculus capping protein (actin filament) muscle Z-line, beta | 100% | NM_001271405.1 |
| (Capzb), transcript variant 3, mRNA | ||||
| 18 | Up | Mus musculus alpha-2-HS-glycoprotein (Ahsg), mRNA | 83% | NM_013465.1 |
| 19 | Up | Mus musculus hydroxysteroid (17-beta) dehydrogenase 11 | 99% | NM_053262.3 |
| (Hsd17b11), mRNA | ||||
| 20 | Down | Mus musculus cytochrome P450, family 2, subfamily a, polypeptide 5 | 99% | NM_007812.4 |
| (Cyp2a5), mRNA | ||||
| 21 | Up | Mus musculus alpha 1 microglobulin/bikunin, mRNA | 99% | BC021660.1 |
| (cDNA clone MGC:14070 IMAGE:4193922), complete cds | ||||
| 22 | Up | Mus musculus cytochrome P450, family 2, subfamily d, polypeptide 26, mRNA | 100% | BC023241.1 |
| (cDNA clone MGC:28632 IMAGE:4222538), complete cds | ||||
| 23 | Up | Mus musculus FCF1 small subunit (SSU) processome component homolog | 97% | NM_028632.2 |
| (S. cerevisiae) (Fcf1), mRNA | ||||
| 24 | Up | Mus musculus pyrophosphatase (inorganic) 1, mRNA | 99% | BC010468.1 |
| (cDNA clone MGC:6716 IMAGE:3585780), complete cds |
| DEG No. | Regulation (Infected) | Identified mRNA | Max Identity | Accession |
|---|---|---|---|---|
| 25 | Up | Mus musculus esterase D/formylglutathione hydrolase, mRNA (cDNA clone MGC:57923 IMAGE:5694094), complete cds | 99% | BC046766.1 |
| 26 | Down | Mus musculus pregnane X receptor mRNA, complete cds | 97% | AF031814.1 |
| 27 | Down | Mus musculus hairy and enhancer of split 6 (Drosophila) (Hes6), mRNA | 99% | NM_019479.3 |
| 28 | Down | Mus musculus mitochondrial pyruvate carrier 1, pseudogene (Mpc1-ps) on chromosome 12 | 99% | NG_032669.1 |
| 29 | Up | Mus musculus interferon, alpha-inducible protein 27 like 1 (Ifi271), transcript variant 5, mRNA | 99% | NM_194069.2 |
| 30 | Down | Mus musculus ribosomal protein L18A (Rpl18a), mRNA | 81% | NM_029751.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wickremsinghe, I.A.C.; R. M. T. Balasubramaniam, V.; Mot, Y.Y.; Dhanoa, A.; S. Hassan, S. Identification of Differentially Expressed Genes in BALB/c Mouse Liver upon Primary Infection with DENV1 and Sequential Heterologous Infection with DENV2. Pathogens 2018, 7, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens7040078
Wickremsinghe IAC, R. M. T. Balasubramaniam V, Mot YY, Dhanoa A, S. Hassan S. Identification of Differentially Expressed Genes in BALB/c Mouse Liver upon Primary Infection with DENV1 and Sequential Heterologous Infection with DENV2. Pathogens. 2018; 7(4):78. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens7040078
Chicago/Turabian StyleWickremsinghe, Indeevari A. C., Vinod R. M. T. Balasubramaniam, Y. Yik Mot, Amreeta Dhanoa, and Sharifah S. Hassan. 2018. "Identification of Differentially Expressed Genes in BALB/c Mouse Liver upon Primary Infection with DENV1 and Sequential Heterologous Infection with DENV2" Pathogens 7, no. 4: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens7040078