Janthinobacterium lividum as An Emerging Pathogenic Bacterium Affecting Rainbow Trout (Oncorhynchus mykiss) Fisheries in Korea


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Bacterial Isolation and Identification
2.3. 16S rRNA Gene Sequencing
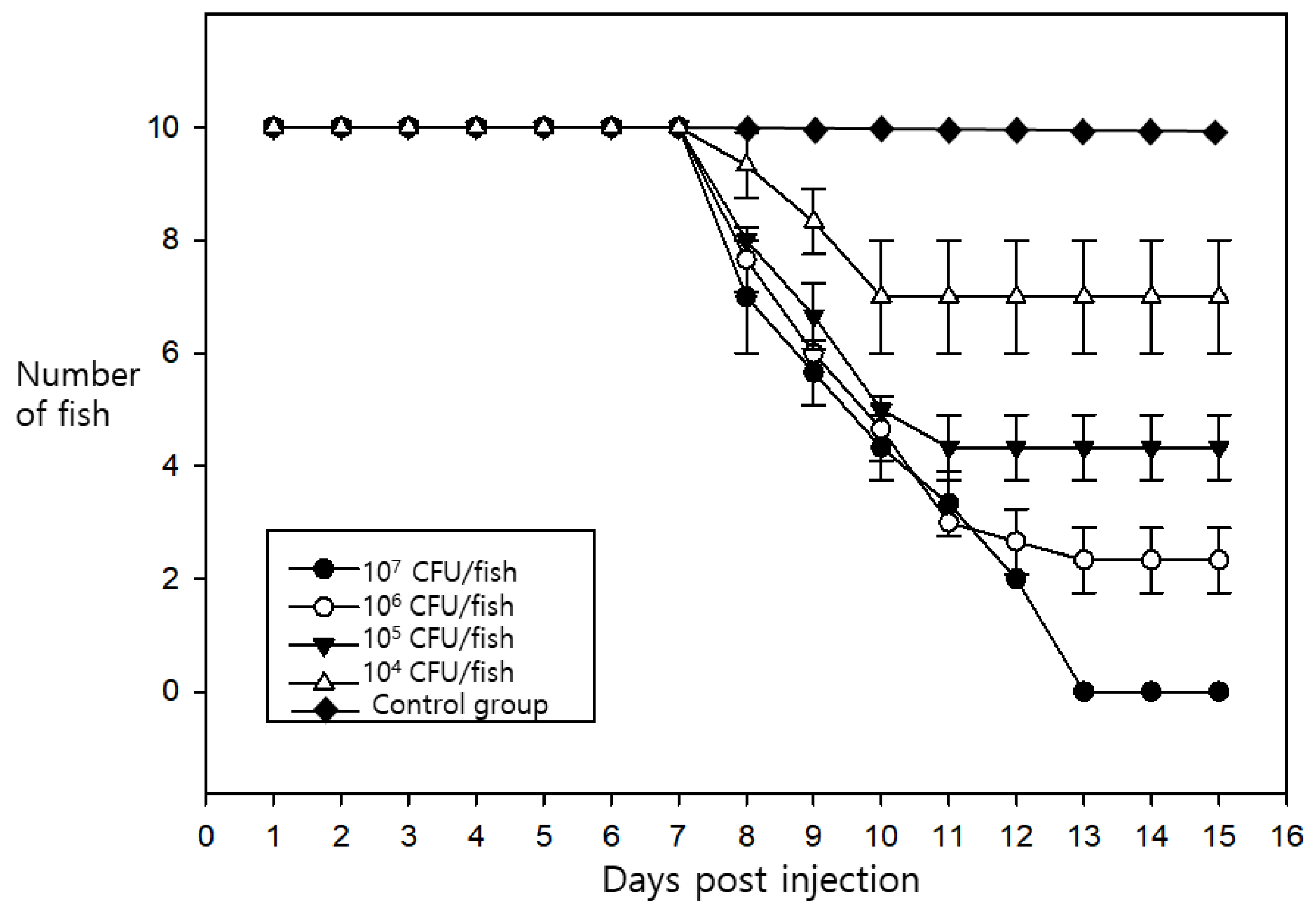
2.4. Pathogenicity Challenge Trials
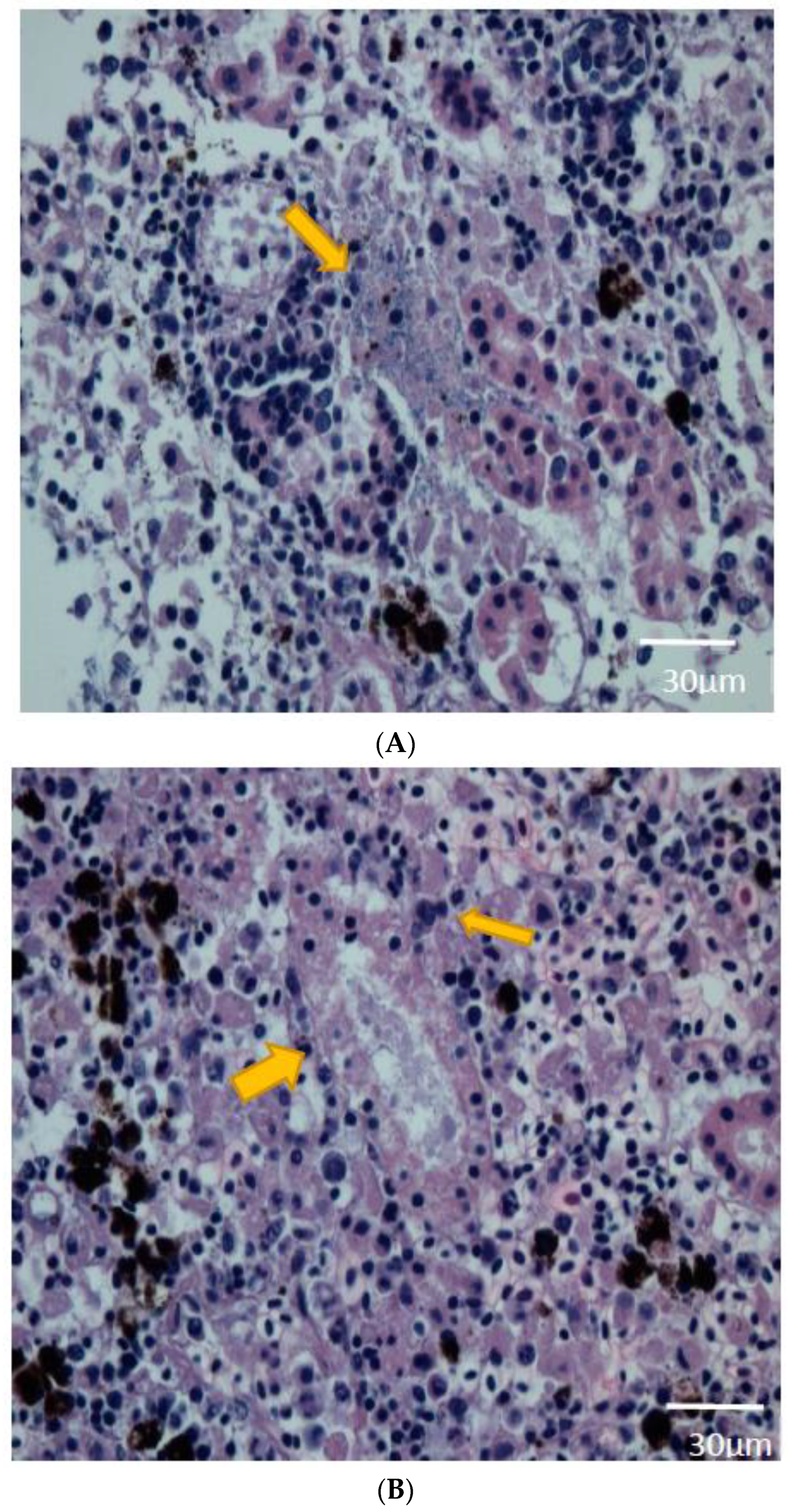
2.5. Histopathological Findings
2.6. Antibiotic Susceptibility Test
3. Results and Discussion
3.1. Identification of the Bacteria
3.2. Pathogenicity Challenge Trials and Clinical Signs
3.3. Histopathologic Analysis
3.4. Antibiotic Susceptibility
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMP | ampicillin |
| PIP | piperacillin |
| CFZ | cefazolin |
| FEP | cefepime |
| CTX | cefotaxime |
| FOX | cefoxitin |
| CAZ | ceftazidime |
| ZOX | ceftizoxime |
| ATM | aztreonam |
| IPM | imipenem |
| MEM | meropenem |
| GEN | gentamicin |
| AMK | amikacin |
| KAN | kanamycin |
| STR | streptomycin |
| TET | tetracycline |
| DOX | doxycycline |
| CIP | ciprofloxacin |
| NAL | nalidixic acid |
| NOR | norfloxacin |
| OFX | ofloxacin |
| TMP | trimethoprim sulfamethoxazole |
| CHL | chloramphenicol |
| ERY | erythromycin |
| R | resistant |
| I | intermediate |
| S | susceptible |
References
- Valdes, N.; Soto, P.; Cottet, L.; Alarcon, P.; Gonzalez, A.; Castillo, A.; Corsini, G.; Tello, M. Draft genome sequence of Janthinobacterium lividum strain MTR reveals its mechanism of capnophilic behavior. Stand. Genom. Sci. 2015, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Allen, H.K.; Klimowicz, A.K.; Mlot, C.; Gross, J.A.; Savengsuksa, S.; McEllin, J.; Clardy, J.; Ruess, R.W.; Handelsman, J. Psychrotrophic strain of Janthinobacterium lividum from a cold Alaskan soil produces prodigiosin. DNA Cell Biol. 2010, 29, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Pantanella, F.; Berlutti, F.; Passariello, C.; Sarli, S.; Morea, C.; Schippa, S. Violacein and biofilm production in Janthinobacterium lividum. J. Appl. Microbiol. 2007, 102, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Baricz, A.; Teban, A.; Chiriac, C.M.; Szekeres, E.; Farkas, A.; Nica, M.; Dascalu, A.; Oprisan, C.; Lavin, P.; Coman, C. Investigating the potential use of an Antarctic variant of Janthinobacterium lividum for tackling antimicrobial resistance in a One Health approach. Sci. Rep. 2018, 8, 15272. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.N.; Brucker, R.M.; Walke, J.B.; Becker, M.H.; Schwantes, C.R.; Flaherty, D.C.; Lam, B.A.; Woodhams, D.C.; Briggs, C.J.; Vredenburg, V.T.; et al. Skin microbes on frogs prevent morbidity and mortality caused by a lethal skin fungus. ISME J. 2009, 3, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Matz, C.; Deines, P.; Boenigk, J.; Arndt, H.; Eberl, L.; Kjelleberg, S.; Jurgens, K. Impact of violacein-Producing bacteria on survival and feeding of bacterivorous nanoflagellates. Appl. Environ. Microbiol. 2004, 70, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Austin, B.; Gonzalez, C.J.; Stobie, M.; Curry, J.I.; McLoughlin, M.F. Recovery of Janthinobacterium lividum from diseased rainbow trout, Oncorhynchus mykiss (Walbaum), in Northern Ireland and Scotland. J. Fish Dis. 1992, 15, 357–359. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The gut microbiota of marine fish. Front. Microbiol. 2018, 9, 873. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [PubMed]
- Dixon, P.; Paley, R.; Alegria-Moran, R.; Oidtmann, B. Epidemiological characteristics of infectious hematopoietic necrosis virus (IHNV): A review. Vet. Res. 2016, 47, 63. [Google Scholar] [CrossRef] [PubMed]
- Altinok, I. Multiplex PCR assay for detection of four major bacterial pathogens causing rainbow trout disease. Dis. Aquat. Organ. 2011, 93, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, B.H.; Cunningham, S.A.; Dailey, A.L.; Gustafson, D.R.; Patel, R. Identification of anaerobic bacteria by Bruker Biotyper matrix-Assisted laser desorption ionization–Time of flight mass spectrometry with on-Plate formic acid preparation. J. Clin. Microbiol. 2013, 51, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Patjanasoontorn, B.; Boonma, P.A.I.T.; Wilailackana, C.; Sittikesorn, J. Hospital acquired Janthinobacterium lividum septicemia in Srinagarind Hospital. J. Med. Assoc. Thail. 1992, 75, 6–10. [Google Scholar]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Ling, T.A. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: A 10.5-year analysis of susceptibilities of Candida species to fluconazole and voriconazole as determined by CLSI standardized disk diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Shin, S.C.; Hong, S.G.; Lee, Y.M.; Lee, H.; Lee, J.; Choi, I.G.; Park, H. Genome sequence of Janthinobacterium sp. strain PAMC 25724, isolated from alpine glacier cryoconite. J. Bacteriol. 2012, 194, 2096. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bergh, P.; Frey, J. Aeromonas salmonicida subsp. salmonicida in the light of its type-Three secretion system. Microb. Biotechnol. 2014, 7, 381–400. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antibiotics | AMP | PIP | CFZ | FEP | CTX | FOX | CAZ | ZOX |
| Resistance | R | R | R | R | R | R | R | R |
| Inhibition zone diameter (mm) | 10 | 11 | 9 | 5 | 10 | 15 | 13 | 13 |
| Antibiotics | ATM | IPM | MEM | GEN | AMK | KAN | STR | TET |
| Resistance | R | R | R | R | R | R | R | I |
| Inhibition zone diameter (mm) | 9 | 10 | 11 | 6 | 8 | 8 | 7 | 14 |
| Antibiotics | DOX | CIP | NAL | NOR | OFX | TMP | CHL | ERY |
| Resistance | S | R | R | R | R | S | R | R |
| Inhibition zone diameter (mm) | 17 | 13 | 5 | 8 | 9 | 20 | 6 | 11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, W.T.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; Jun, J.W.; et al. Janthinobacterium lividum as An Emerging Pathogenic Bacterium Affecting Rainbow Trout (Oncorhynchus mykiss) Fisheries in Korea. Pathogens 2019, 8, 146. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8030146
Oh WT, Giri SS, Yun S, Kim HJ, Kim SG, Kim SW, Kang JW, Han SJ, Kwon J, Jun JW, et al. Janthinobacterium lividum as An Emerging Pathogenic Bacterium Affecting Rainbow Trout (Oncorhynchus mykiss) Fisheries in Korea. Pathogens. 2019; 8(3):146. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8030146
Chicago/Turabian StyleOh, Woo Taek, Sib Sankar Giri, Saekil Yun, Hyoun Joong Kim, Sang Guen Kim, Sang Wha Kim, Jeong Woo Kang, Se Jin Han, Jun Kwon, Jin Woo Jun, and et al. 2019. "Janthinobacterium lividum as An Emerging Pathogenic Bacterium Affecting Rainbow Trout (Oncorhynchus mykiss) Fisheries in Korea" Pathogens 8, no. 3: 146. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens8030146