Proteomic Profiling of the Liver, Hepatic Lymph Nodes, and Spleen of Buffaloes Infected with Fasciola gigantica

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
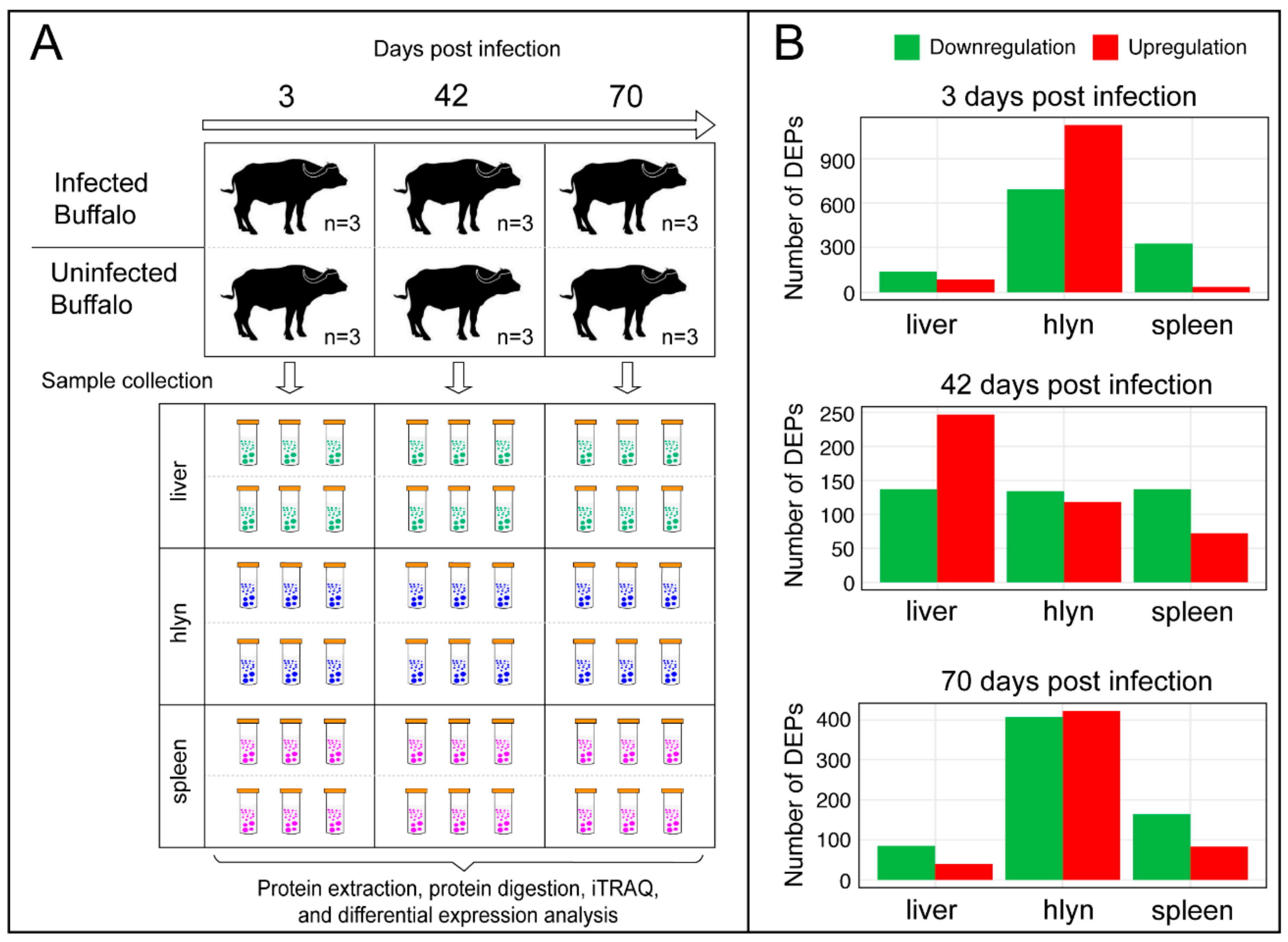
2.1. Animals and Experimental Infection
2.2. Protein Extraction and iTRAQ Analysis
2.3. Bioinformatic Analysis
2.4. Verification of Proteomic Results Using Parallel Reaction Monitoring (PRM)
3. Results
3.1. Detection of F. gigantica in Infected Buffaloes
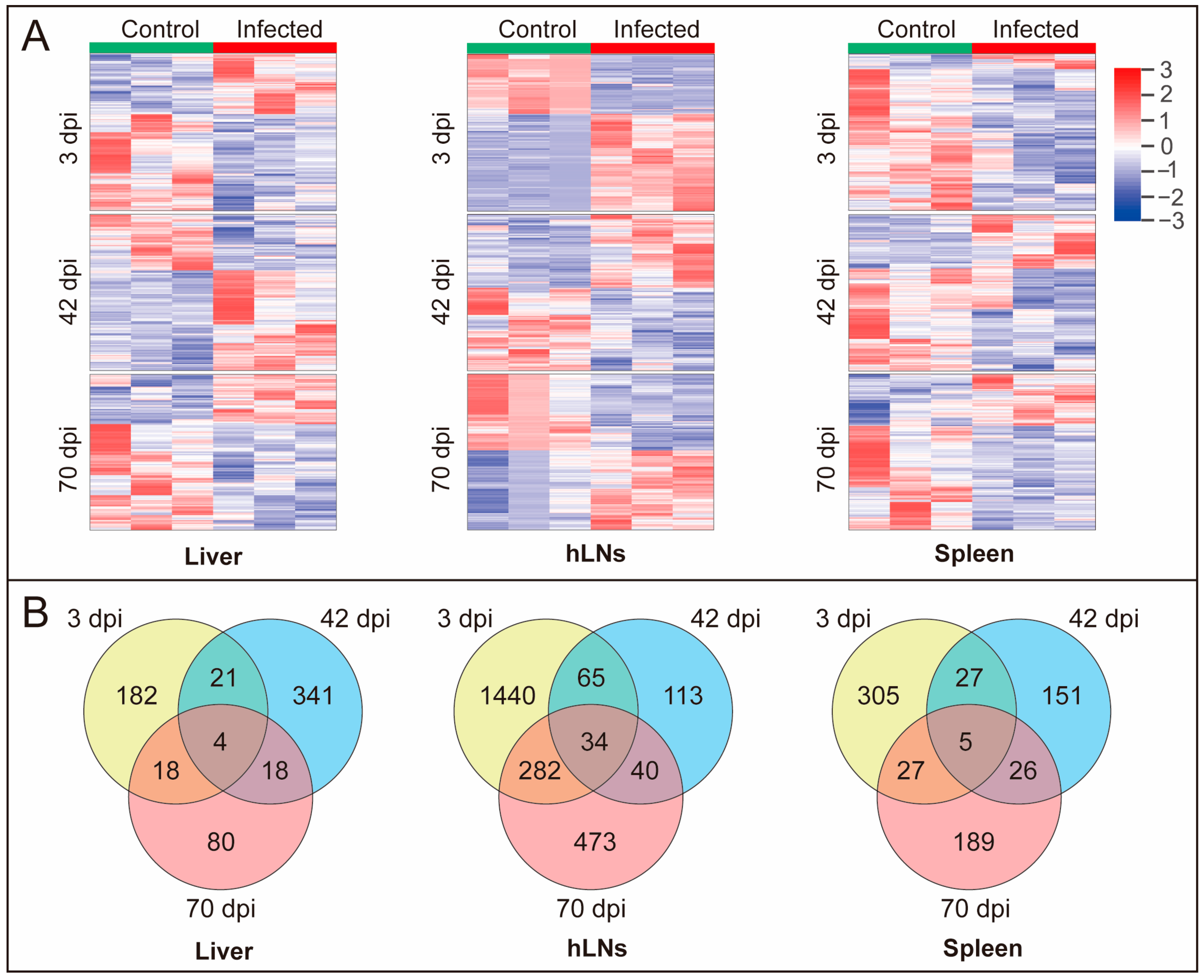
3.2. DEPs affected by F. gigantica Infection
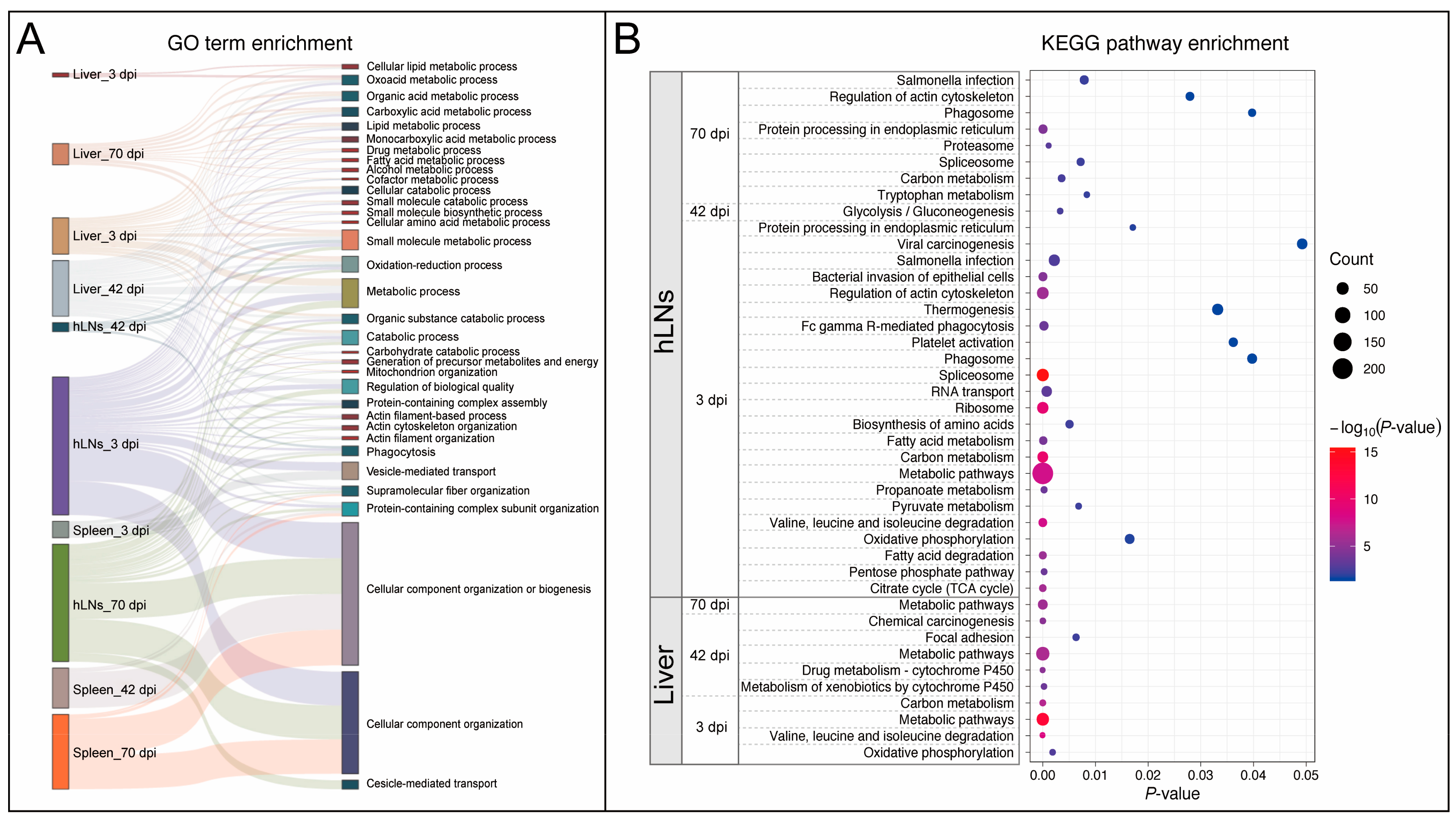
3.3. Functional Enrichment Analysis of the Identified DEPs
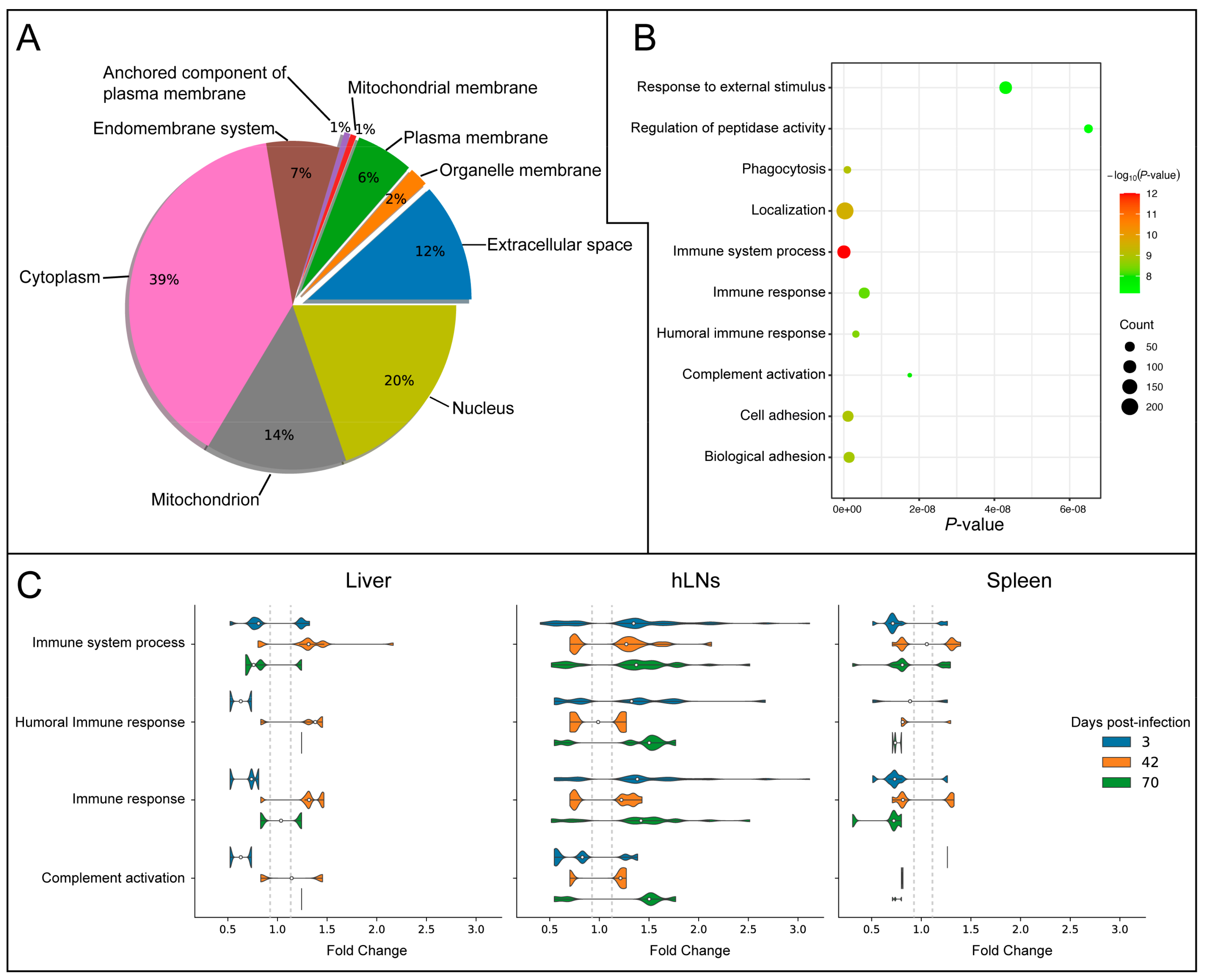
3.4. Subcellular Localization Analysis of DEPs
4. Discussion
4.1. Hepatic Response to F. gigantica Infection
4.2. More Immune Response in hLNs at Early Compared to Late Stages of Infection
4.3. Downregulated Proteomic Signature in Spleen Throughout the Infection Course
4.4. Membrane and Extracellular Space Proteins Mediate Host Immune Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethics Approval
References
- Fürst, T.; Keiser, J.; Utzinger, J. Global burden of human food-borne trematodiasis: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 210–221. [Google Scholar] [CrossRef]
- Cwiklinski, K.; O’Neill, S.M.; Donnelly, S.; Dalton, J.P. A prospective view of animal and human Fasciolosis. Parasite Immunol. 2016, 38, 558–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S.J. The Life Cycle of Fasciola hepatica; Dalton, J.P., Ed.; CABI Publishing: Wallingford, UK, 1999; pp. 1–29. [Google Scholar]
- Howell, A. Snail-Borne Diseases in Bovids at High and Low Altitude in Eastern Uganda: Integrated Parasitological and Malacological Mapping; Liverpool School of Tropical Medicine: Liverpool, UK, August 2011. [Google Scholar]
- Tian, A.L.; Lu, M.; Calderón-Mantilla, G.; Petsalaki, E.; Dottorini, T.; Tian, X.; Wang, Y.; Huang, S.Y.; Hou, J.L.; Li, X.; et al. A recombinant Fasciola gigantica 14-3-3 epsilon protein (rFg14-3-3e) modulates various functions of goat peripheral blood mononuclear cells. Parasit. Vectors 2018, 11, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, A.L.; Lu, M.; Zhang, F.K.; Calderón-Mantilla, G.; Petsalaki, E.; Tian, X.; Wang, W.; Huang, S.Y.; Li, X.; Elsheikha, H.M.; et al. The pervasive effects of recombinant Fasciola gigantica Ras-related protein Rab10 on the functions of goat peripheral blood mononuclear cells. Parasit. Vectors 2018, 11, 579. [Google Scholar] [CrossRef] [Green Version]
- Mei, X.; Shi, W.; Zhao, W.; Luo, H.; Zhang, Y.; Wang, Y.; Sheng, Z.; Wang, D.; Zhu, X.Q.; Huang, W. Fasciola gigantica excretory-secretory products (FgESPs) modulate the differentiation and immune functions of buffalo dendritic cells through a mechanism involving DNMT1 and TET1. Parasit. Vectors 2020, 13, 355. [Google Scholar] [CrossRef]
- Zhang, Y.; Mei, X.; Liang, Y.; Zhu, B.; Sheng, Z.; Shi, W.; Wang, D.; Huang, W. Newly excysted juveniles (NEJs) of Fasciola gigantica induce mice liver fibrosis and M2 macrophage-like phenotype in vivo. Microb. Pathog. 2020, 139, 103909. [Google Scholar] [CrossRef]
- Cwiklinski, K.; Jewhurst, H.; McVeigh, P.; Barbour, T.; Maule, A.G.; Tort, J.; O’Neill, S.M.; Robinson, M.W.; Donnelly, S.; Dalton, J.P. Infection by the helminth parasite Fasciola hepatica requires rapid regulation of metabolic, virulence, and invasive factors to adjust to its mammalian host. Mol. Cell. Proteom. 2018, 17, 792–809. [Google Scholar] [CrossRef] [Green Version]
- Verissimo, C.D.M.; Jewhurst, H.L.; Tikhonova, I.G.; Urbanus, R.T.; Maule, A.G.; Dalton, J.P.; Cwiklinski, K. Fasciola hepatica serine protease inhibitor family (serpins): Purposely crafted for regulating host proteases. PLoS Negl. Trop. Dis. 2020, 14, e0008510. [Google Scholar] [CrossRef]
- Zhang, F.K.; Hu, R.S.; Elsheikha, H.M.; Sheng, Z.A.; Zhang, W.; Zheng, W.B.; Zhu, X.Q.; He, J.J. Global serum proteomic changes in water buffaloes infected with Fasciola gigantica. Parasit. Vectors 2019, 12, 281. [Google Scholar] [CrossRef] [Green Version]
- Changklungmoa, N.; Kueakhai, P.; Sangpairoj, K.; Osotprasit, S.; Chaiwichien, A.; Samrit, T.; Sobhon, P.; Chaithirayanon, K. A novel thioredoxin-related protein 14 from Fasciola gigantica has an immunodiagnostic potential for fasciolosis. Acta Trop. 2020, 207, 105471. [Google Scholar] [CrossRef]
- Pandey, T.; Ghosh, A.; Todur, V.N.; Rajendran, V.; Kalita, P.; Kalita, J.; Shukla, R.; Chetri, P.B.; Shukla, H.; Sonkar, A.; et al. Draft genome of the liver fluke Fasciola gigantica. ACS Omega 2020, 5, 11084–11091. [Google Scholar] [CrossRef] [PubMed]
- Tian, A.L.; Tian, X.; Chen, D.; Lu, M.; Calderón-Mantilla, G.; Yuan, X.D.; Li, X.; Elsheikha, H.M.; Zhu, X.Q. Modulation of the functions of goat peripheral blood mononuclear cells by Fasciola gigantica thioredoxin peroxidase in vitro. Pathogens 2020, 9, 758. [Google Scholar] [CrossRef] [PubMed]
- Panic, G.; Duthaler, U.; Speich, B.; Keiser, J. Repurposing drugs for the treatment and control of helminth infections. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 185–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, J.M.; Elliott, T.P.; Beddoe, T.; Anderson, G.; Skuce, P.; Spithill, T.W. Current threat of triclabendazole resistance in Fasciola hepatica. Trends Parasitol. 2016, 32, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toet, H.; Piedrafita, D.M.; Spithill, T.W. Liver fluke vaccines in ruminants: Strategies, progress and future opportunities. Int. J. Parasitol. 2014, 44, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.K.; Zhang, X.X.; Elsheikha, H.M.; He, J.J.; Sheng, Z.A.; Zheng, W.B.; Ma, J.G.; Huang, W.Y.; Guo, A.J.; Zhu, X.Q. Transcriptomic responses of water buffalo liver to infection with the digenetic fluke Fasciola gigantica. Parasit. Vectors 2017, 10, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, M.; Tian, Z. Liver-mediated adaptive immune tolerance. Front. Immunol. 2019, 10, 2525. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.X.; Cwiklinski, K.; Hu, R.S.; Zheng, W.B.; Sheng, Z.A.; Zhang, F.K.; Elsheikha, H.M.; Dalton, J.P.; Zhu, X.Q. Complex and dynamic transcriptional changes allow the helminth Fasciola gigantica to adjust to its intermediate snail and definitive mammalian hosts. BMC Genom. 2019, 20, 729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savojardo, C.; Martelli, P.L.; Fariselli, P.; Profiti, G.; Casadio, R. BUSCA: An integrative web server to predict subcellular localization of proteins. Nucleic Acids Res. 2018, 46, W459–W466. [Google Scholar] [CrossRef] [PubMed]
- Zauber, H.; Kirchner, M.; Selbach, M. Picky: A simple online PRM and SRM method designer for targeted proteomics. Nat. Methods 2018, 15, 156–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.M.; Arnold, L.L. Chemical carcinogenesis. Toxicol. Sci. 2010, 120, S76–S92. [Google Scholar] [CrossRef] [PubMed]
- Machicado, C.; Machicado, J.D.; Maco, V.; Terashima, A.; Marcos, L.A. Association of Fasciola hepatica infection with liver fibrosis, cirrhosis, and cancer: A systematic review. PLoS Negl. Trop. Dis. 2016, 10, e0004962. [Google Scholar] [CrossRef] [Green Version]
- Lynch, T.; Price, A. The effect of cytochrome P450 metabolism on drug response, interactions, and adverse effects. Am. Fam. Physician 2007, 76, 391–396. [Google Scholar] [PubMed]
- Maté, L.; Giantin, M.; Viviani, P.; Ballent, M.; Tolosi, R.; Lifschitz, A.; Lanusse, C.; Dacasto, M.; Virkel, G. Effects of fenbendazole and triclabendazole on the expression of cytochrome P450 1A and flavin-monooxygenase isozymes in bovine precision-cut liver slices. Vet. J. 2019, 245, 61–69. [Google Scholar] [CrossRef]
- Fairweather, I.; Brennan, G.; Hanna, R.; Robinson, M.; Skuce, P. Drug resistance in liver flukes. Int. J. Parasitol. Drugs Drug Resist. 2020, 12, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.-H.; Simbi, B.H.; Ford, L.; Cole, S.R.; Taylor, M.J.; Lawson, C.; Lawrence, R.A. Live Brugia malayi microfilariae inhibit transendothelial migration of neutrophils and monocytes. PLoS Negl. Trop. Dis. 2012, 6, e1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham-Brown, J.; Hartley, C.; Clough, H.; Kadioglu, A.; Baylis, M.; Williams, D.J.L.; Palmer, G.H. Dairy heifers naturally exposed to Fasciola hepatica develop a Type 2 immune response and concomitant suppression of leukocyte proliferation. Infect. Immun. 2017, 86, e00607-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knipper, J.A.; Ivens, A.; Taylor, M.D. Helminth-induced Th2 cell dysfunction is distinct from exhaustion and is maintained in the absence of antigen. PLoS Negl. Trop. Dis. 2019, 13, e0007908. [Google Scholar] [CrossRef]
- Yong, L.; Tang, Y.; Ren, C.; Liu, M.; Shen, J.; Hou, X. B1 cells protect against Schistosoma japonicum–induced liver inflammation and fibrosis by controlling monocyte infiltration. PLoS Negl. Trop. Dis. 2019, 13, e0007474. [Google Scholar] [CrossRef]
- Peixoto, R.; Silva, L.M.; López-Osório, S.; Zhou, E.; Gärtner, U.; Conejeros, I.; Taubert, A.; Hermosilla, C. Fasciola hepatica induces weak NETosis and low production of intra- and extracellular ROS in exposed bovine polymorphonuclear neutrophils. Dev. Comp. Immunol. 2021, 114, 103787. [Google Scholar] [CrossRef]
- Schmitt, M.E.; Lutz, J.; Haase, P.; Bösl, M.R.; Wienands, J.; Engels, N.; Voehringer, D. The B-cell antigen receptor of IgE-switched plasma cells regulates memory IgE responses. J. Allergy Clin. Immunol. 2020, 146, 642–651.e5. [Google Scholar] [CrossRef] [PubMed]
- Cosenza-Contreras, M.; Castro, R.A.D.O.E.; Mattei, B.; Campos, J.M.; Silva, G.G.; De Paiva, N.C.N.; Aguiar-Soares, R.D.D.O.; Carneiro, C.M.; Afonso, L.C.C.; Castro-Borges, W. The schistosomiasis spleenOME: Unveiling the proteomic landscape of splenomegaly using label-free mass spectrometry. Front. Immunol. 2019, 9, 3137. [Google Scholar] [CrossRef] [PubMed]
- De La Torre-Escudero, E.; Gerlach, J.Q.; Bennett, A.P.S.; Cwiklinski, K.; Jewhurst, H.L.; Huson, K.M.; Joshi, L.; Kilcoyne, M.; O’Neill, S.; Dalton, J.P.; et al. Surface molecules of extracellular vesicles secreted by the helminth pathogen Fasciola hepatica direct their internalisation by host cells. PLoS Negl. Trop. Dis. 2019, 13, e0007087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swan, J.; Sakthivel, D.; Cameron, T.C.; Faou, P.; Downs, R.; Rajapaksha, H.; Piedrafita, D.; Beddoe, T. Proteomic identification of galectin-11 and -14 ligands from Fasciola hepatica. Int. J. Parasitol. 2019, 49, 921–932. [Google Scholar] [CrossRef]
- Davis, F.P.; Barkan, D.T.; Eswar, N.; McKerrow, J.H.; Sali, A. Host-pathogen protein interactions predicted by comparative modeling. Protein Sci. 2007, 16, 2585–2596. [Google Scholar] [CrossRef] [Green Version]
- Geurts, N.; Opdenakker, G.; Steen, P.E.V.D. Matrix metalloproteinases as therapeutic targets in protozoan parasitic infections. Pharmacol. Ther. 2012, 133, 257–279. [Google Scholar] [CrossRef]
- Cuesta-Astroz, Y.; Santos, A.; Oliveira, G.; Jensen, L.J. Analysis of predicted host–parasite interactomes reveals commonalities and specificities related to parasitic lifestyle and tissues tropism. Front. Immunol. 2019, 10, 212. [Google Scholar] [CrossRef] [Green Version]
- Nowacki, F.C.; Swain, M.T.; Klychnikov, O.I.; Niazi, U.; Ivens, A.; Quintana, J.F.; Hensbergen, P.J.; Hokke, C.H.; Buck, A.H.; Hoffmann, K.F. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. J. Extracell. Vesicles 2015, 4, 28665. [Google Scholar] [CrossRef] [Green Version]
- Fromm, B.; Ovchinnikov, V.Y.; Høye, E.; Bernal, L.; Hackenberg, M.; Marcilla, A. On the presence and immunoregulatory functions of extracellular microRNAs in the trematode Fasciola hepatica. Parasite Immunol. 2017, 39, e12399. [Google Scholar] [CrossRef]
- Sotillo, J.; Robinson, M.W.; Kimber, M.J.; Cucher, M.; Ancarola, M.E.; Nejsum, P.; Marcilla, A.; Eichenberger, R.M.; Tritten, L. The protein and microRNA cargo of extracellular vesicles from parasitic helminths–current status and research priorities. Int. J. Parasitol. 2020, 50, 635–645. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, R.-S.; Zhang, F.-K.; Elsheikha, H.M.; Ma, Q.-N.; Ehsan, M.; Zhao, Q.; Zhu, X.-Q. Proteomic Profiling of the Liver, Hepatic Lymph Nodes, and Spleen of Buffaloes Infected with Fasciola gigantica. Pathogens 2020, 9, 982. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9120982
Hu R-S, Zhang F-K, Elsheikha HM, Ma Q-N, Ehsan M, Zhao Q, Zhu X-Q. Proteomic Profiling of the Liver, Hepatic Lymph Nodes, and Spleen of Buffaloes Infected with Fasciola gigantica. Pathogens. 2020; 9(12):982. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9120982
Chicago/Turabian StyleHu, Rui-Si, Fu-Kai Zhang, Hany M. Elsheikha, Qiao-Ni Ma, Muhammad Ehsan, Quan Zhao, and Xing-Quan Zhu. 2020. "Proteomic Profiling of the Liver, Hepatic Lymph Nodes, and Spleen of Buffaloes Infected with Fasciola gigantica" Pathogens 9, no. 12: 982. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens9120982