
Comparative Characterization of CpCDPK1 and CpCDPK9, Two Potential Drug Targets Against Cryptosporidiosis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cryptosporidium parvum Isolate and Invasion Assay
2.2. Construction of Recombinant Plasmid
2.3. Expression of Recombinant CpCDPKs in E. coli
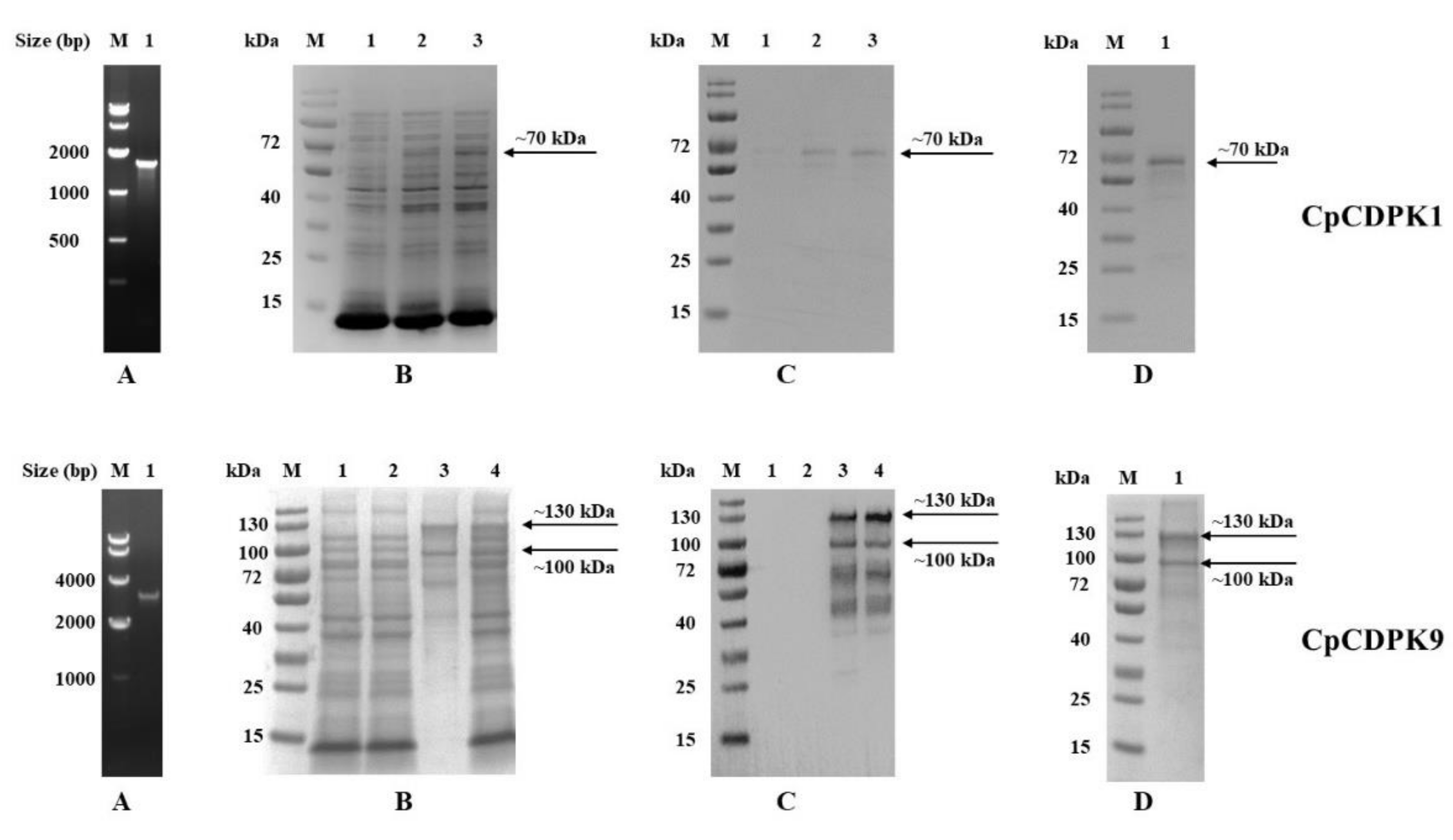
2.4. Purification of CDPKs
2.5. Assessment of Enzymatic Activity of Recombinant CDPKs
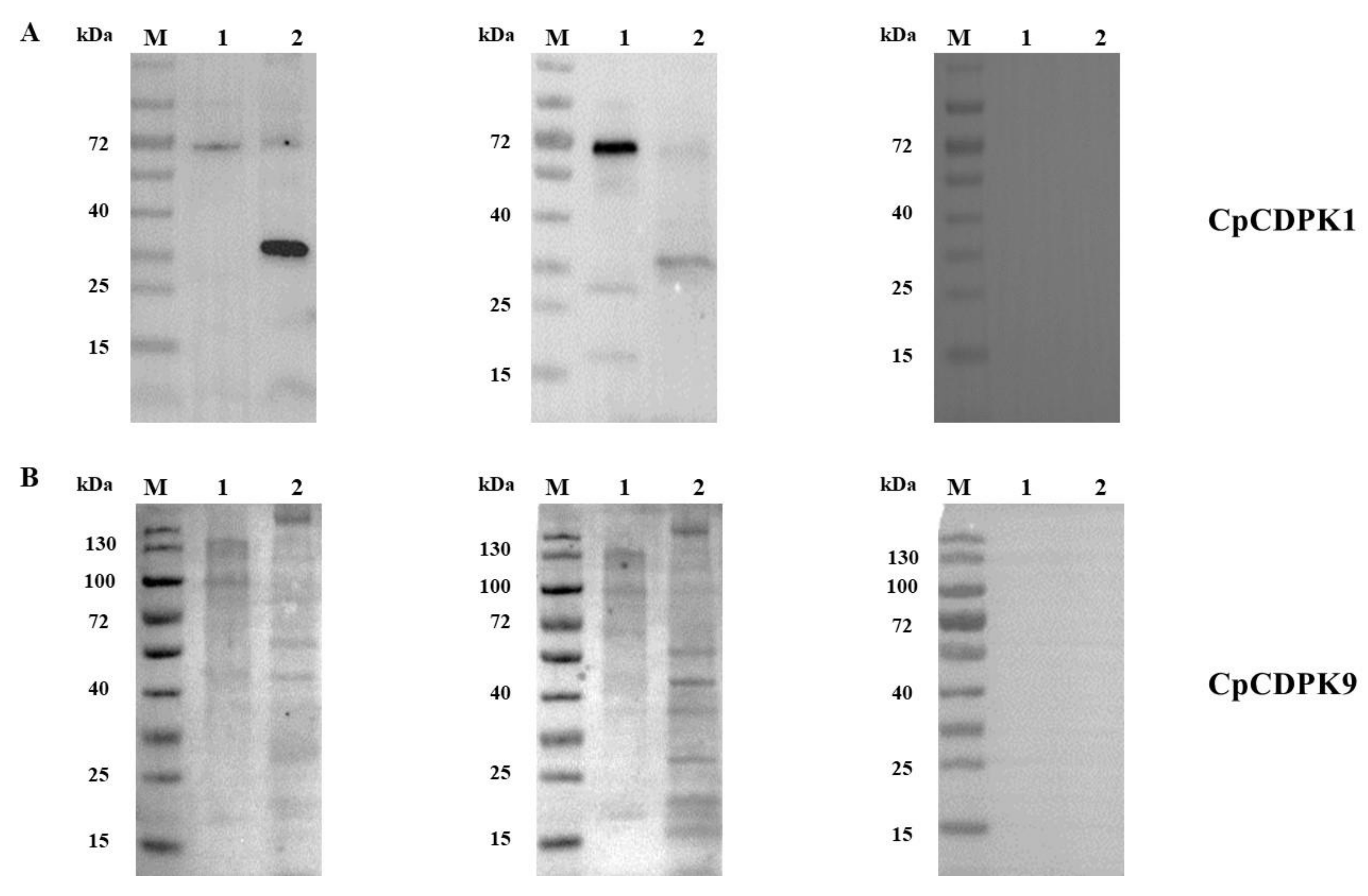
2.6. Preparation of Anti-CDPK Antibodies and Assessment of Their Reactivity to Native Proteins
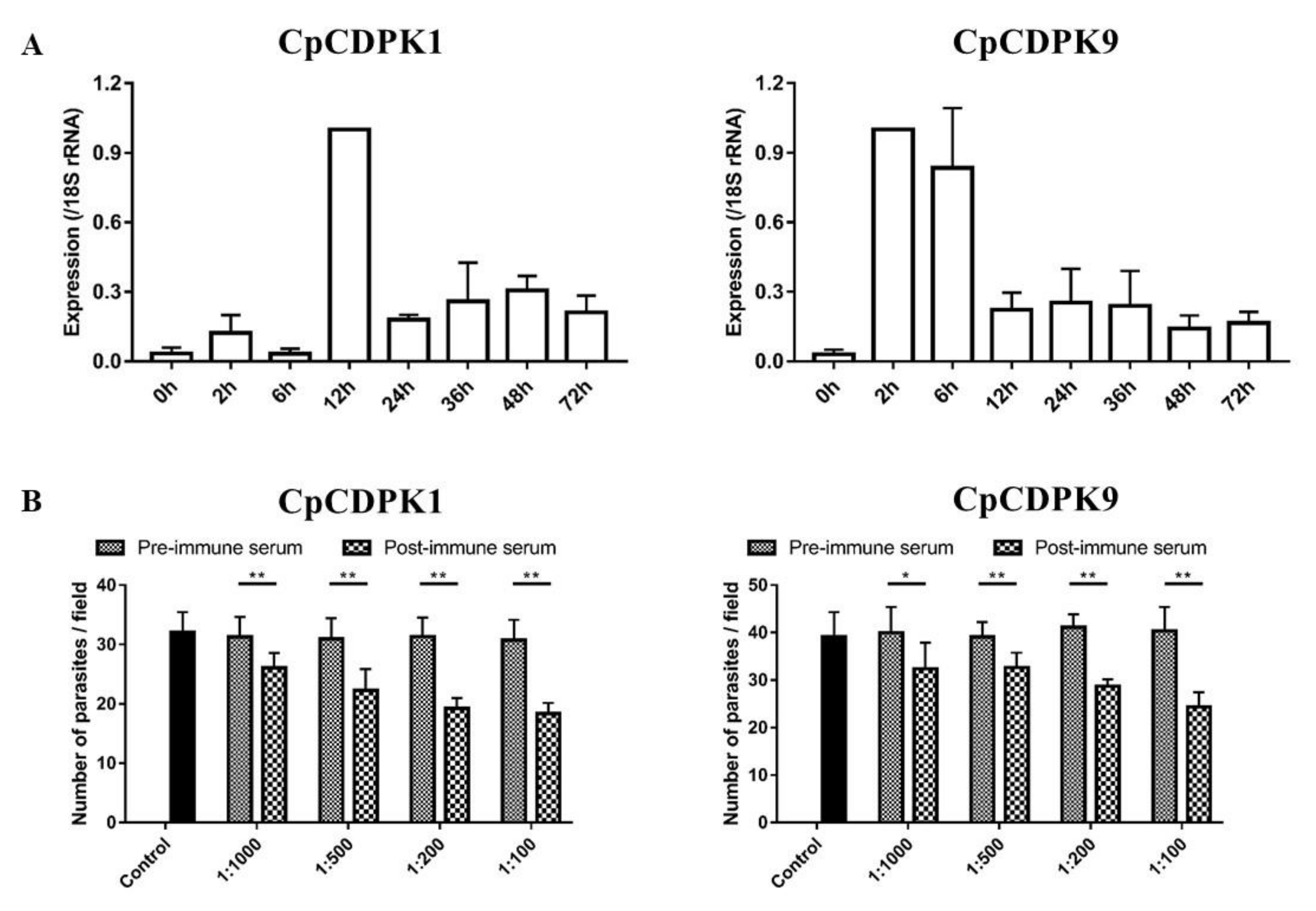
2.7. Quantitation of CDPK Gene Expression
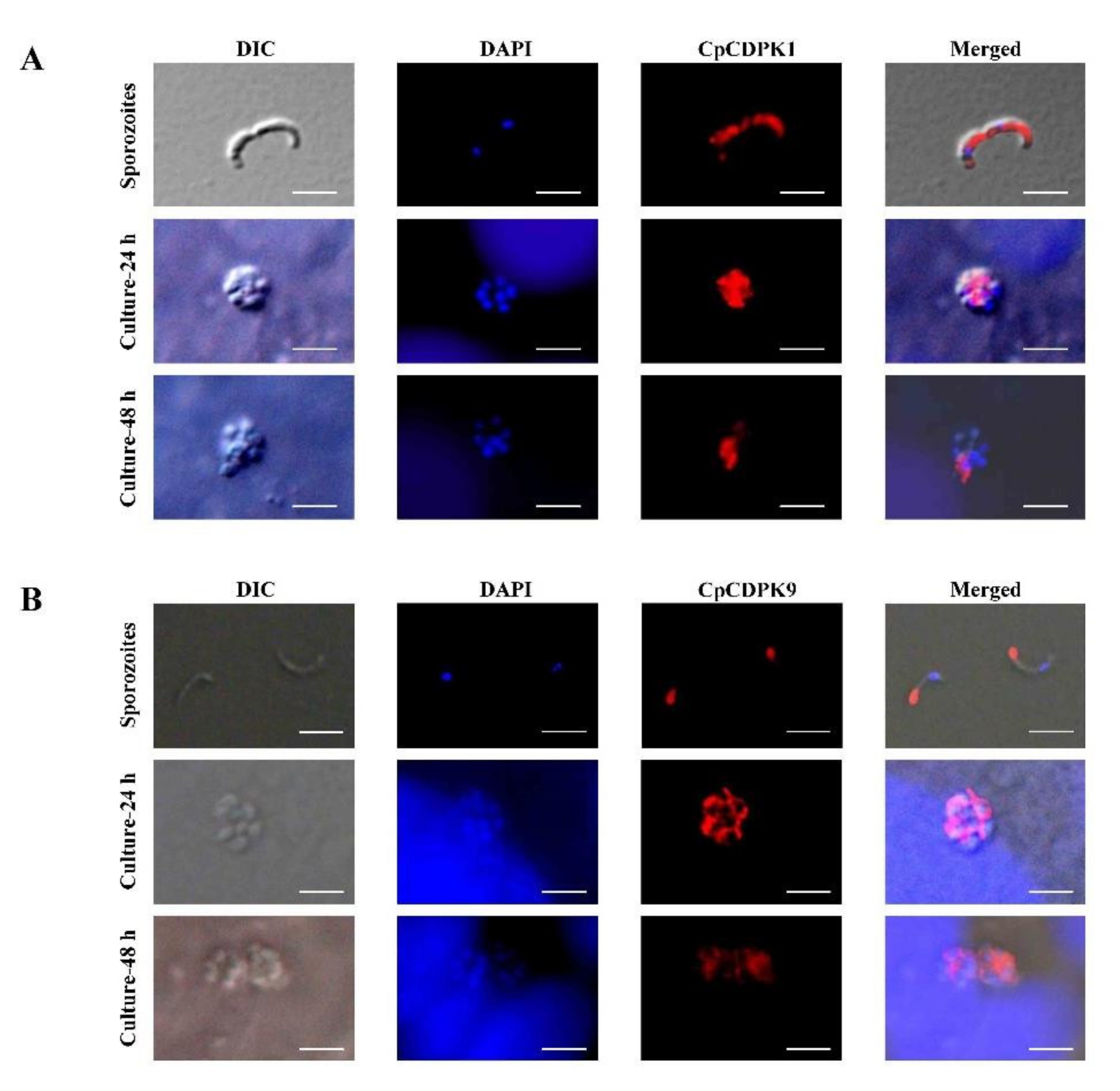
2.8. Localization of CDPK Protein Expression in Life Cycle Stages
2.9. In Vitro Neutralization of Sporozoite Invasion
In Vitro Inhibition of Invasion and Development using Candidate CpCDPK Inhibitors
3. Results
3.1. Production of Recombinant CpCDPKs in E. coli
3.2. Expression of Native CDPKs in Sporozoites
3.3. CDPK expression in Developmental Stages
3.4. Expression of CDPK Genes in C. parvum Culture
3.5. Neutralization of Sporozoite Invasion by CDPK Antibodies
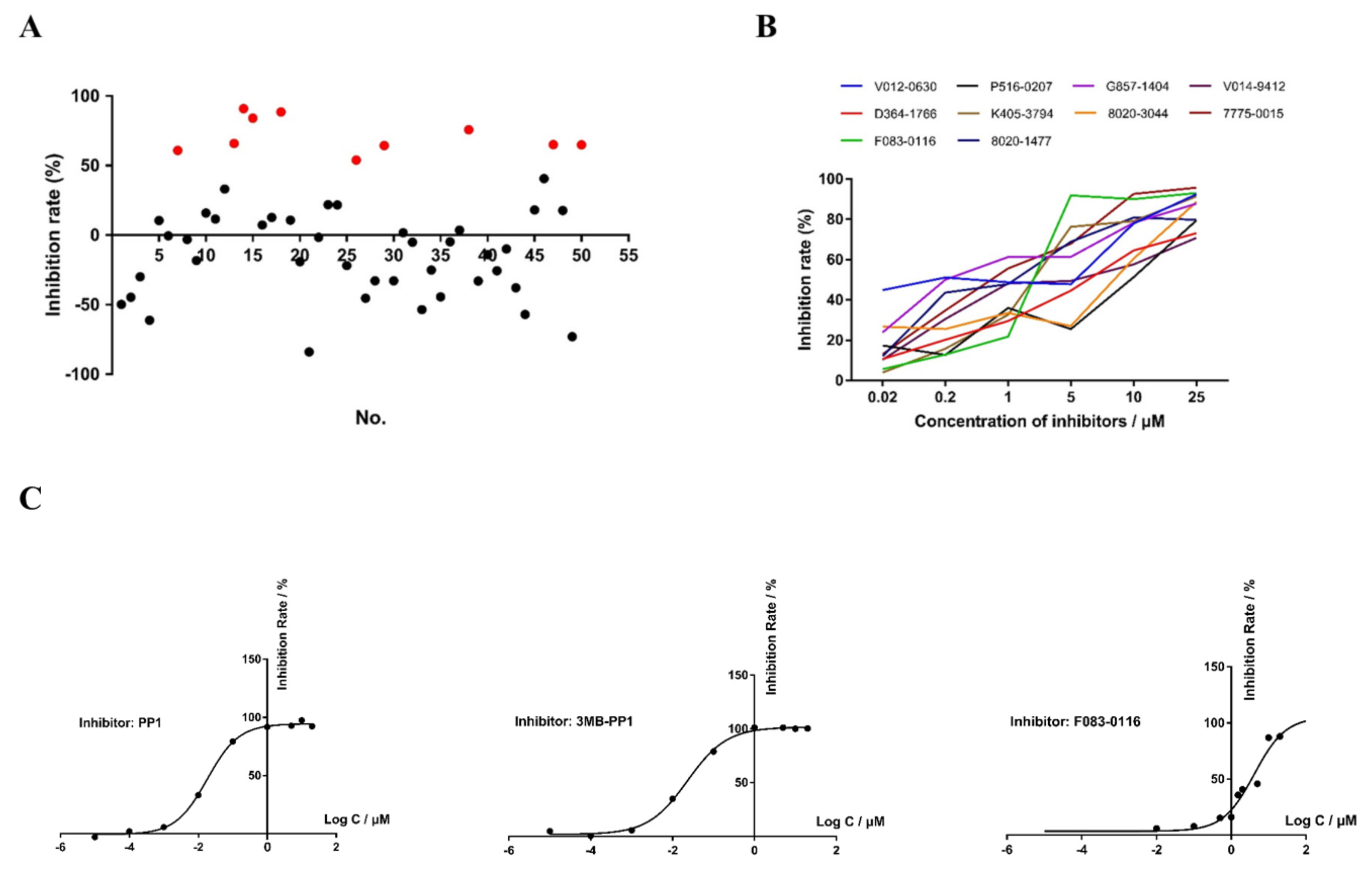
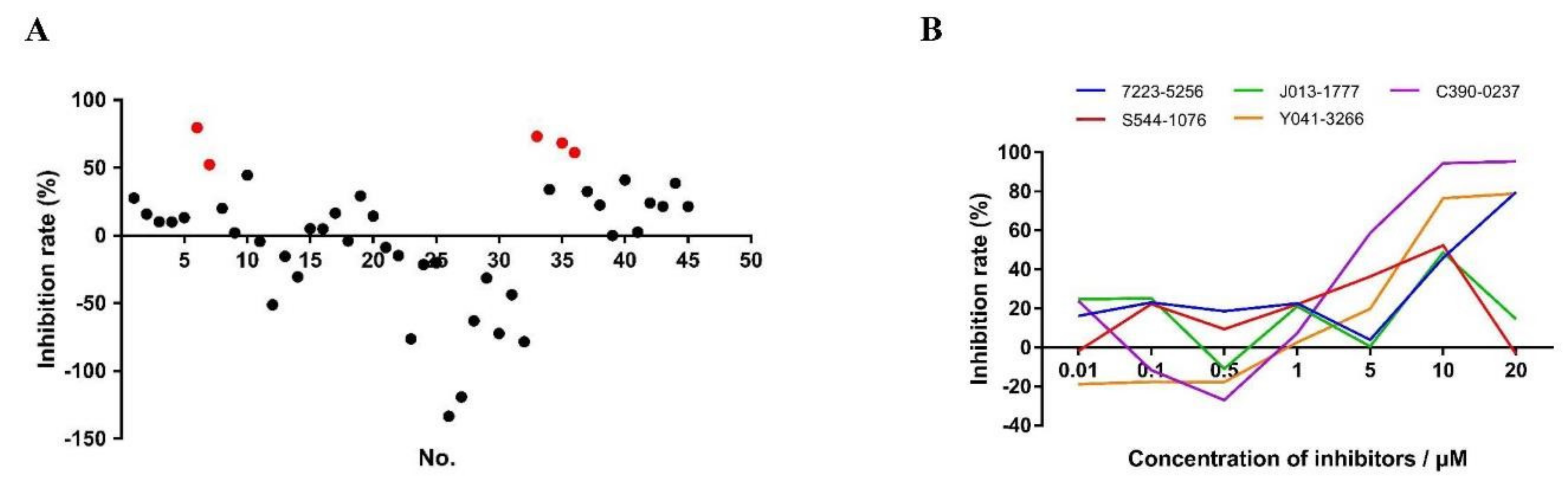
3.6. In Vitro Anti-Cryptosporidial Activities of Small Molecule Compounds
3.7. Effects of Candidate CDPK Inhibitors on Enzymatic Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Checkley, W.; White, A.C., Jr.; Jaganath, D.; Arrowood, M.J.; Chalmers, R.M.; Chen, X.M.; Fayer, R.; Griffiths, J.K.; Guerrant, R.L.; Hedstrom, L.; et al. A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for Cryptosporidium. Lancet Infect. Dis. 2015, 15, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, R.M.; Davies, A.P. Minireview: Clinical cryptosporidiosis. Exp. Parasitol. 2010, 124, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ryan, U.M.; Xiao, L. Genetic diversity and population structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Pantenburg, B.; Cabada, M.M.; White, A.C., Jr. Treatment of cryptosporidiosis. Expert Rev. Anti Infect Ther. 2009, 7, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Ghartey-Kwansah, G.; Yin, Q.; Li, Z.; Gumpper, K.; Sun, Y.; Yang, R.; Wang, D.; Jones, O.; Zhou, X.; Wang, L.; et al. Calcium-dependent Protein Kinases in Malaria Parasite Development and Infection. Cell Transplant. 2020, 29, 963689719884888. [Google Scholar] [CrossRef] [PubMed]
- Etzold, M.; Lendner, M.; Daugschies, A.; Dyachenko, V. CDPKs of Cryptosporidium parvum-stage-specific expression in vitro. Parasitol. Res. 2014, 113, 2525–2533. [Google Scholar] [CrossRef] [PubMed]
- Wernimont, A.K.; Artz, J.D.; Finerty, P., Jr.; Lin, Y.H.; Amani, M.; Allali-Hassani, A.; Senisterra, G.; Vedadi, M.; Tempel, W.; Mackenzie, F.; et al. Structures of apicomplexan calcium-dependent protein kinases reveal mechanism of activation by calcium. Nat. Struct. Mol. Biol. 2010, 17, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.F.; Harmon, A. Plants, symbiosis and parasites: A calcium signalling connection. Nat. Rev. Mol. Cell Biol. 2005, 6, 555–566. [Google Scholar] [CrossRef]
- Lourido, S.; Shuman, J.; Zhang, C.; Shokat, K.M.; Hui, R.; Sibley, L.D. Calcium-dependent protein kinase 1 is an essential regulator of exocytosis in Toxoplasma. Nature 2010, 465, 359–362. [Google Scholar] [CrossRef] [Green Version]
- Chavez, M.A.; White, A.C., Jr. Novel treatment strategies and drugs in development for cryptosporidiosis. Expert Rev. Anti Infect. Ther. 2018, 16, 655–661. [Google Scholar] [CrossRef]
- Murphy, R.C.; Ojo, K.K.; Larson, E.T.; Castellanos-Gonzalez, A.; Perera, B.G.; Keyloun, K.R.; Kim, J.E.; Bhandari, J.G.; Muller, N.R.; Verlinde, C.L.; et al. Discovery of Potent and Selective Inhibitors of Calcium-Dependent Protein Kinase 1 (CDPK1) from C. parvum and T. gondii. ACS Med. Chem. Lett. 2010, 1, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Guo, Y.; Li, N.; Li, Y.; Su, J.; Xu, R.; Zhang, Z.; Feng, Y.; Xiao, L. Characterization of Calcium-Dependent Protein Kinases 3, a Protein Involved in Growth of Cryptosporidium Parvum. Front. Microbiol. 2020, 11, 907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Shao, Q.; Guo, Y.; Li, N.; Li, Y.; Su, J.; Xu, R.; Zhang, Z.; Xiao, L.; Feng, Y. Characterization of Three Calcium-Dependent Protein Kinases of Cryptosporidium parvum. Front. Microbiol. 2020, 11, 622203. [Google Scholar] [CrossRef] [PubMed]
- Castellanos-Gonzalez, A.; Sparks, H.; Nava, S.; Huang, W.; Zhang, Z.; Rivas, K.; Hulverson, M.A.; Barrett, L.K.; Ojo, K.K.; Fan, E.; et al. A Novel Calcium-Dependent Kinase Inhibitor, Bumped Kinase Inhibitor 1517, Cures Cryptosporidiosis in Immunosuppressed Mice. J. Infect. Dis. 2016, 214, 1850–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, H.H.; Nava, M.G.; Gartlan, B.E.; Rose, S.; Vinayak, S. A Conditional Protein Degradation System To Study Essential Gene Function in Cryptosporidium parvum. mBio 2020, 11, e01231-20. [Google Scholar] [CrossRef]
- Hulverson, M.A.; Vinayak, S.; Choi, R.; Schaefer, D.A.; Castellanos-Gonzalez, A.; Vidadala, R.S.R.; Brooks, C.F.; Herbert, G.T.; Betzer, D.P.; Whitman, G.R.; et al. Bumped-Kinase Inhibitors for Cryptosporidiosis Therapy. J. Infect. Dis. 2017, 215, 1275–1284. [Google Scholar] [CrossRef] [Green Version]
- Kuhlenschmidt, T.B.; Rutaganira, F.U.; Long, S.; Tang, K.; Shokat, K.M.; Kuhlenschmidt, M.S.; Sibley, L.D. Inhibition of Calcium-Dependent Protein Kinase 1 (CDPK1) In Vitro by Pyrazolopyrimidine Derivatives Does Not Correlate with Sensitivity of Cryptosporidium parvum Growth in Cell Culture. Antimicrob. Agents Chemother. 2016, 60, 570–579. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Wu, H.; Li, N.; Su, J.; Jia, R.; Jiang, J.; Feng, Y.; Xiao, L. Preliminary Characterization of MEDLE-2, a Protein Potentially Involved in the Invasion of Cryptosporidium parvum. Front. Microbiol. 2017, 8, 1647. [Google Scholar] [CrossRef]
- Su, J.; Jin, C.; Wu, H.; Fei, J.; Li, N.; Guo, Y.; Feng, Y.; Xiao, L. Differential Expression of Three Cryptosporidium Species-Specific MEDLE Proteins. Front. Microbiol. 2019, 10, 1177. [Google Scholar] [CrossRef]
- Cai, X.; Woods, K.M.; Upton, S.J.; Zhu, G. Application of quantitative real-time reverse transcription-PCR in assessing drug efficacy against the intracellular pathogen Cryptosporidium parvum in vitro. Antimicrob. Agents Chemother. 2005, 49, 4437–4442. [Google Scholar] [CrossRef] [Green Version]
- Mauzy, M.J.; Enomoto, S.; Lancto, C.A.; Abrahamsen, M.S.; Rutherford, M.S. The Cryptosporidium parvum transcriptome during in vitro development. PLoS ONE 2012, 7, e31715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jakobi, V.; Petry, F. Differential expression of Cryptosporidium parvum genes encoding sporozoite surface antigens in infected HCT-8 host cells. Microbes Infect. 2006, 8, 2186–2194. [Google Scholar] [CrossRef] [PubMed]
- Upton, S.J.; Tilley, M.; Brillhart, D.B. Effects of select medium supplements on in vitro development of Cryptosporidium parvum in HCT-8 cells. J. Clin. Microbiol. 1995, 33, 371–375. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, G. Quantitative RT-PCR assay for high-throughput screening (HTS) of drugs against the growth of Cryptosporidium parvum in vitro. Front. Microbiol. 2015, 6, 991. [Google Scholar] [CrossRef] [Green Version]
- Persechini, A.; Moncrief, N.D.; Kretsinger, R.H. The EF-hand family of calcium-modulated proteins. Trends Neurosci. 1989, 12, 462–467. [Google Scholar] [CrossRef]
- Harmon, A.C.; Gribskov, M.; Harper, J.F. CDPKs-a kinase for every Ca2+ signal? Trends Plant Sci. 2000, 5, 154–159. [Google Scholar] [CrossRef]
- Billker, O.; Lourido, S.; Sibley, L.D. Calcium-dependent signaling and kinases in apicomplexan parasites. Cell Host Microbe 2009, 5, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Bradley, P.J.; Boothroyd, J.C. Identification of the pro-mature processing site of Toxoplasma ROP1 by mass spectrometry. Mol. Biochem. Parasitol. 1999, 100, 103–109. [Google Scholar] [CrossRef]
- Miller, S.A.; Thathy, V.; Ajioka, J.W.; Blackman, M.J.; Kim, K. TgSUB2 is a Toxoplasma gondii rhoptry organelle processing proteinase. Mol. Microbiol. 2003, 49, 883–894. [Google Scholar] [CrossRef]
- Kieschnick, H.; Wakefield, T.; Narducci, C.A.; Beckers, C. Toxoplasma gondii attachment to host cells is regulated by a calmodulin-like domain protein kinase. J. Biol. Chem. 2001, 276, 12369–12377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Mirdha, B.R.; Srinivasan, A.; Rukmangadachar, L.A.; Singh, S.; Sharma, P.; Hariprasad, G.; Gururao, H.; Luthra, K. Identification of invasion proteins of Cryptosporidium parvum. World J. Microbiol. Biotechnol. 2015, 31, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Sibley, L.D. Intracellular parasite invasion strategies. Science 2004, 304, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Govindasamy, K.; Bhanot, P. Overlapping and distinct roles of CDPK family members in the pre-erythrocytic stages of the rodent malaria parasite, Plasmodium berghei. PLoS Pathog. 2020, 16, e1008131. [Google Scholar] [CrossRef]
- Huang, W.; Choi, R.; Hulverson, M.A.; Zhang, Z.; McCloskey, M.C.; Schaefer, D.A.; Whitman, G.R.; Barrett, L.K.; Vidadala, R.S.R.; Riggs, M.W.; et al. 5-aminopyrazole-4-carboxamide-based compounds prevent the growth of Cryptosporidium parvum. Antimicrob. Agents Chemother. 2017, 61, e00020-17. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | No. | Compound | No. | Compound |
|---|---|---|---|---|---|
| 1 | D008-0179 | 18 | 8020-3044 | 35 | C499-0205 |
| 2 | F472-0429 | 19 | 8019-3516 | 36 | D730-0071 |
| 3 | S021-0111 | 20 | G856-6766 | 37 | J106-0712 |
| 4 | E518-0433 | 21 | F722-0391 | 38 | 8020-1477 |
| 5 | C753-1493 | 22 | 8015-7028 | 39 | F470-0515 |
| 6 | D046-0013 | 23 | J101-0311 | 40 | C118-0100 |
| 7 | V012-0630 | 24 | T482-1629 | 41 | D126-0439 |
| 8 | D392-0696 | 25 | S824-0226 | 42 | D006-0238 |
| 9 | D008-0217 | 26 | P516-0207 | 43 | 4514-2069 |
| 10 | C879-1492 | 27 | 5685-0834 | 44 | Y041-7886 |
| 11 | S021-0065 | 28 | D226-0165 | 45 | 3028-4958 |
| 12 | 8012-3948 | 29 | K405-3794 | 46 | 7919-0012 |
| 13 | D364-1766 | 30 | Y020-1235 | 47 | V014-9412 |
| 14 | F083-0116 | 31 | G419-0776 | 48 | P759-1388 |
| 15 | G857-1404 | 32 | D271-0476 | 49 | D401-0791 |
| 16 | E455-0304 | 33 | C197-0058 | 50 | 7775-0015 |
| 17 | D426-1054 | 34 | C066-1821 |
| No. | Compound | No. | Compound | No. | Compound |
|---|---|---|---|---|---|
| 1 | 8012-9491 | 16 | S616-8028 | 31 | Y041-3613 |
| 2 | P896-0008 | 17 | P896-0153 | 32 | C276-1003 |
| 3 | D271-0053 | 18 | F401-0004 | 33 | 7223-5256 |
| 4 | P937-2651 | 19 | 4428-0282 | 34 | M976-0119 |
| 5 | S641-4525 | 20 | Y041-6010 | 35 | S544-1076 |
| 6 | C390-0237 | 21 | F722-0985 | 36 | J013-1777 |
| 7 | Y041-3206 | 22 | 2995-0027 | 37 | 4358-6015 |
| 8 | J106-0113 | 23 | D074-0339 | 38 | D126-0082 |
| 9 | Y020-6058 | 24 | 7655-0020 | 39 | 8013-1632 |
| 10 | G639-3793 | 25 | 8018-3260 | 40 | D585-0146 |
| 11 | G072-0423 | 26 | S021-0079 | 41 | S629-0195 |
| 12 | D021-0194 | 27 | D389-0696 | 42 | Y020-2366 |
| 13 | P814-5254 | 28 | D330-0081 | 43 | D361-0101 |
| 14 | E687-0240 | 29 | D718-0573 | 44 | Y021-0940 |
| 15 | M510-0341 | 30 | J106-0289 | 45 | C200-7683 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, J.; Shen, Y.; Li, N.; Li, Y.; Zhang, Z.; Xiao, L.; Guo, Y.; Feng, Y. Comparative Characterization of CpCDPK1 and CpCDPK9, Two Potential Drug Targets Against Cryptosporidiosis. Microorganisms 2022, 10, 333. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020333
Su J, Shen Y, Li N, Li Y, Zhang Z, Xiao L, Guo Y, Feng Y. Comparative Characterization of CpCDPK1 and CpCDPK9, Two Potential Drug Targets Against Cryptosporidiosis. Microorganisms. 2022; 10(2):333. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020333
Chicago/Turabian StyleSu, Jiayuan, Yiting Shen, Na Li, Yu Li, Ziding Zhang, Lihua Xiao, Yaqiong Guo, and Yaoyu Feng. 2022. "Comparative Characterization of CpCDPK1 and CpCDPK9, Two Potential Drug Targets Against Cryptosporidiosis" Microorganisms 10, no. 2: 333. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020333