
Detection, Molecular Identification and Transmission of the Intestinal Protozoa Blastocystis sp. in Guinea from a Large-Scale Epidemiological Study Conducted in the Conakry Area

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Questionnaire Survey
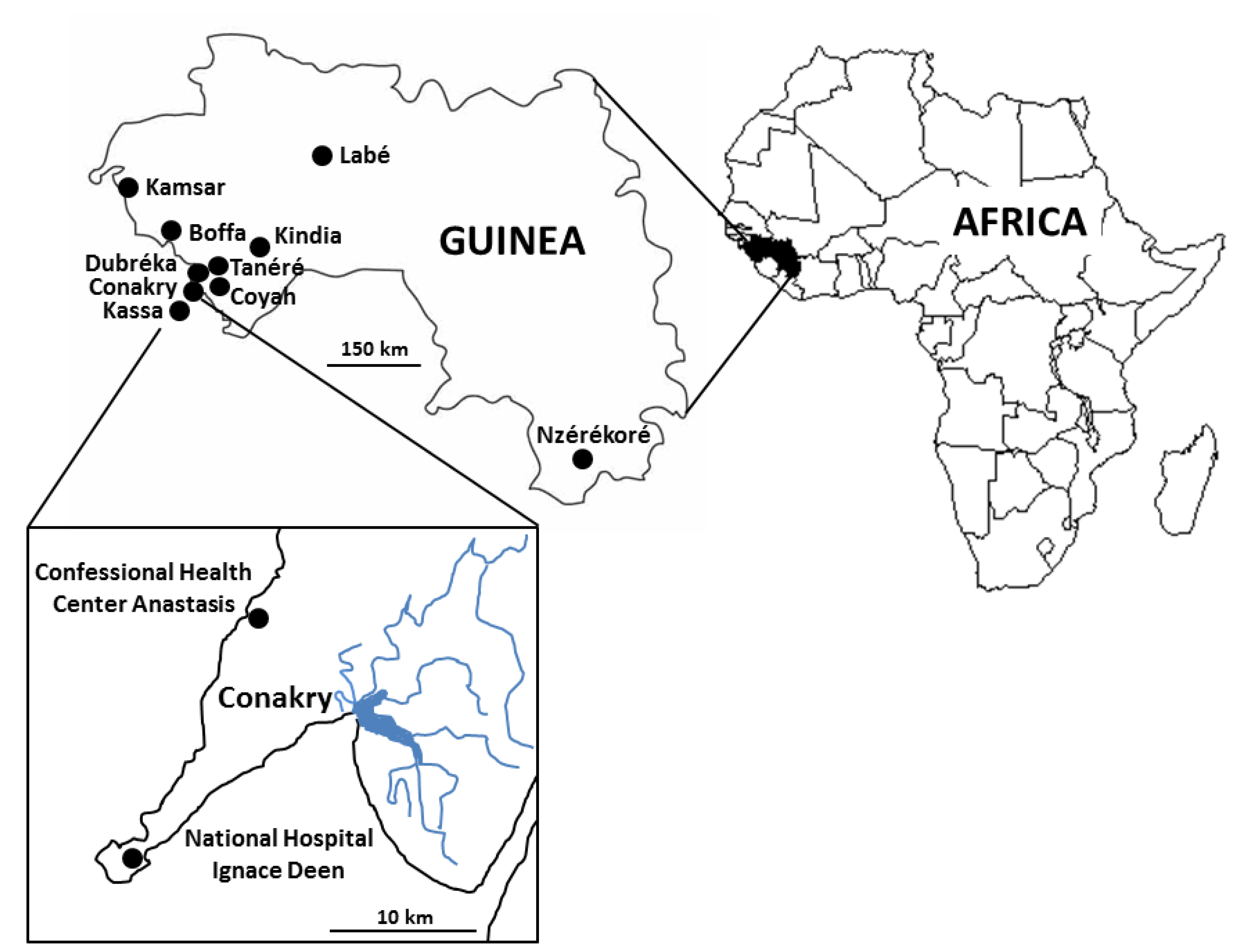
2.3. Sampling Sites and Collection of Samples
2.4. DNA Extraction and Molecular Subtyping of Blastocystis sp. Isolates
2.5. Statistical Analysis
3. Results
3.1. Analysis of the Cohort of Guinean Subjects, Prevalence and Risk Factors Associated to Blastocystis sp. Infection
3.2. Distribution of Blastocystis sp. STs
3.3. Identification of Blastocystis sp. Genotypes and Analysis of Intra-ST Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boreham, P.F.L.; Stenzel, D.J. Blastocystis in humans and animals: Morphology, biology, and epizootiology. Adv. Parasitol. 1993, 32, 1–70. [Google Scholar] [PubMed]
- Tan, K.S.W. Blastocystis in humans and animals: New insights using modern methodologies. Vet. Parasitol. 2004, 126, 121–144. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Koyama, Y.; Tsuchiya, E.; Takami, K. Blastocystis phylogeny among various isolates from humans to insects. Parasitol. Int. 2016, 65, 750–759. [Google Scholar] [CrossRef]
- Cian, A.; El Safadi, D.; Osman, M.; Moriniere, R.; Gantois, N.; Benamrouz-Vanneste, S.; Delgado-Viscogliosi, P.; Guyot, K.; Li, L.L.; Monchy, S.; et al. Molecular epidemiology of Blastocystis sp. in various animal groups from two French zoos and evaluation of potential zoonotic risk. PLoS ONE 2017, 12, e0169659. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.S.W. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clin. Microbiol. Rev. 2008, 21, 639–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanlan, P.D.; Stensvold, C.R. Blastocystis: Getting to grips with our guileful guest. Trends Parasitol. 2013, 29, 523–529. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Yoshida, K.; Nakajima, A.; Yamanari, K.; Iwatani, S.; Kimata, I. Fecal-oral transmission of the cyst form of Blastocystis hominis in rats. Parasitol. Res. 2004, 94, 391–396. [Google Scholar] [CrossRef]
- El Safadi, D.; Gaayeb, L.; Meloni, D.; Cian, A.; Poirier, P.; Wawrzyniak, I.; Delbac, F.; Dabboussi, F.; Delhaes, L.; Seck, M.; et al. Children of Senegal River Basin show the highest prevalence of Blastocystis sp. ever observed worldwide. BMC Infect. Dis. 2014, 14, 164. [Google Scholar] [CrossRef]
- Lokmer, A.; Cian, A.; Froment, A.; Gantois, N.; Viscogliosi, E.; Chabé, M.; Ségurel, L. Use of shotgun metagenomics for the identification of protozoa in the gut microbiota of healthy individuals from worldwide populations with various industrialization levels. PLoS ONE 2019, 14, e0211139. [Google Scholar] [CrossRef]
- Khaled, S.; Gantois, N.; Tidjani Ly, A.; Senghor, S.; Even, G.; Dautel, E.; Dejager, R.; Sawant, M.; Baydoun, M.; Benamrouz-Vanneste, S.; et al. Prevalence and subtype distribution of Blastocystis sp. in Senegalese school children. Microorganisms 2020, 8, 1408. [Google Scholar] [CrossRef]
- Osman, M.; El Safadi, D.; Cian, A.; Benamrouz, S.; Nourrisson, C.; Poirier, P.; Pereira, B.; Razakandrainibe, R.; Pinon, A.; Lambert, C.; et al. Prevalence and risk factors for intestinal protozoan infections with Cryptosporidium, Giardia, Blastocystis and Dientamoeba among schoolchildren in Tripoli, Lebanon. PLoS Negl. Trop. Dis. 2016, 10, e0004496. [Google Scholar]
- Noradilah, S.A.; Moktar, N.; Anuar, T.S.; Lee, I.L.; Salleh, F.M.; Manap, S.N.A.A.; Mohtar, N.S.H.M.; Azrul, S.M.; Abdullah, W.O.; Nordin, A.; et al. Molecular epidemiology of blastocystosis in Malaysia: Does seasonal variation play an important role in determining the distribution and risk factors of Blastocystis subtype infections in the Aboriginal community? Parasit. Vectors 2017, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- El Safadi, D.; Cian, A.; Nourrisson, C.; Pereira, B.; Morelle, C.; Bastien, P.; Bellanger, A.P.; Botterel, F.; Candolfi, E.; Desoubeaux, G.; et al. Prevalence, risk factors for infection and subtype distribution of the intestinal parasite Blastocystis sp. from a large-scale multi-center study in France. BMC Infect. Dis. 2016, 16, 451. [Google Scholar] [CrossRef] [PubMed]
- Muadica, A.S.; Köster, P.C.; Dashti, A.; Bailo, B.; Hernandez-de-Mingo, M.; Reh, L.; Balasegaram, S.; Verlander, N.Q.; Chercoles, E.R.; Carmena, D. Molecular diversity of Giardia duodenalis, Cryptosporidium spp. and Blastocystis sp. in asymptomatic school children in Leganés, Madrid (Spain). Microorganisms 2020, 8, 466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lhotska, Z.; Jirku, M.; Hlozkova, O.; Brozova, K.; Jirsova, D.; Stensvold, C.R.; Kolisko, M.; Pomajbikova, K.J. A study on the prevalence and subtype diversity of the intestinal protist Blastocystis sp. in a gut-healthy human population in the Czech Republic. Front. Cell. Infect. Microbiol. 2020, 10, 544335. [Google Scholar] [CrossRef]
- Clark, C.G.; van der Giezen, M.; Alfellani, M.A.; Stensvold, C.R. Recent developments in Blastocystis research. Adv. Parasitol. 2013, 82, 1–32. [Google Scholar]
- Stensvold, C.R.; Clark, C.G. Current status of Blastocystis: A personal view. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef]
- Andersen, L.O.; Stensvold, C.R. Blastocystis in health and disease: Are we moving from a clinical to a public health perspective? J. Clin. Microbiol. 2016, 54, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Ajjampur, S.S.; Tan, K.S.W. Pathogenic mechanisms in Blastocystis spp.—Interpreting results from in vitro and in vivo studies. Parasitol Int. 2016, 65, 772–779. [Google Scholar] [CrossRef]
- Fréalle, E.; El Safadi, D.; Cian, A.; Aubry, E.; Certad, G.; Osman, M.; Wacrenier, A.; Dutoit, E.; Creusy, C.; Dubos, F.; et al. Acute Blastocystis-associated appendicular peritonitis in a child, Casablanca, Morocco. Emerg. Infect. Dis. 2015, 21, 91–94. [Google Scholar] [CrossRef]
- Denoeud, F.; Roussel, M.; Noel, B.; Wawrzyniak, I.; Da Silva, C.; Diogon, M.; Viscogliosi, E.; Brochier-Armanet, C.; Couloux, A.; Poulain, J.; et al. Genome sequence of the stramenopile Blastocystis, a human anaerobic parasite. Genome Biol. 2011, 12, R29. [Google Scholar] [CrossRef] [PubMed]
- Wawrzyniak, I.; Courtine, D.; Osman, M.; Hubans-Pierlot, C.; Cian, A.; Nourrisson, C.; Chabé, M.; Poirier, P.; Bart, A.; Polonais, V.; et al. Draft genome sequence of the intestinal parasite Blastocystis subtype 4-isolate WR1. Genom. Data 2015, 4, 22–23. [Google Scholar] [CrossRef] [PubMed]
- Gentekaki, E.; Curtis, B.A.; Stairs, C.W.; Klimes, V.; Eliás, M.; Salas-Leiva, D.E.; Herman, E.K.; Eme, L.; Arias, M.C.; Henrissat, B.; et al. Extreme genome diversity in the hyper-prevalent parasitic eukaryote Blastocystis. PLoS Biol. 2017, 15, e2003769. [Google Scholar] [CrossRef] [PubMed]
- Beghini, F.; Pasolli, E.; Truong, T.D.; Putignani, L.; Caccio, S.M.; Segata, N. Large-scale comparative metagenomics of Blastocystis, a common member of the human gut microbiome. ISME J. 2017, 11, 2848–2863. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Wojciech, L.; Gascoigne, N.R.J.; Peng, G.; Tan, K.S.W. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathog. 2021, 17, e1009253. [Google Scholar] [CrossRef] [PubMed]
- Audebert, C.; Even, G.; Cian, A.; Loywick, A.; Merlin, S.; Viscogliosi, E.; Chabé, M.; Blastocystis Investigation Group. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Sci. Rep. 2016, 6, 25255. [Google Scholar] [CrossRef]
- Tito, R.Y.; Chaffron, S.; Caenepeel, C.; Lima-Mendez, G.; Wang, J.; Vieira-Silva, S.; Falony, G.; Hildebrand, F.; Darzi, Y.; Rymenans, L.; et al. Population-level analysis of Blastocystis subtype prevalence and variation in the human gut microbiota. Gut 2019, 68, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Even, G.; Lokmer, A.; Rodrigues, J.; Audebert, C.; Viscogliosi, E.; Ségurel, L.; Chabé, M. Changes in the human gut microbiota associated with colonization by Blastocystis sp. and Entamoeba spp. in non-industrialized populations. Front. Cell. Infect. Microbiol. 2021, 11, 533528. [Google Scholar] [CrossRef]
- Yason, J.A.; Liang, Y.R.; Png, C.W.; Zhang, Y.; Tan, K.S.W. Interactions between a pathogenic Blastocystis subtype and gut microbiota: in vitro and in vivo studies. Microbiome 2019, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Alfellani, M.A.; Taner-Mulla, D.; Jacob, A.S.; Imeede, C.A.; Yoshikawa, H.; Stensvold, C.R.; Clark, C.G. Genetic diversity of Blastocystis in livestock and zoo animals. Protist 2013, 164, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.H.; Hu, X.F.; Liu, T.L.; Hu, R.S.; Yu, Z.Q.; Yang, W.B.; Wu, Y.L.; Yu, S.K.; Song, J.K. Molecular characterization of Blastocystis sp. in captive wild animals in Qinling Mountains. Parasitol. Res. 2017, 116, 2327–2333. [Google Scholar] [CrossRef] [PubMed]
- Maloney, J.G.; Lombard, J.E.; Urie, N.J.; Shivley, C.B.; Santin, M. Zoonotic and genetically diverse subtypes of Blastocystis in US pre-weaned dairy heifer calves. Parasitol. Res. 2019, 118, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Maloney, J.G.; Molokin, A.; Rodrigues da Cunha, M.J.; Cury, M.C.; Santin, M. Blastocystis subtype distribution in domestic and captive wild bird species from Brazil using next generation amplicon sequencing. Parasite Epidemiol. Control 2020, 9, e00138. [Google Scholar] [CrossRef]
- Maloney, J.G.; Rodrigues da Cunha, M.J.; Molokin, A.; Cury, M.C.; Santin, M. Next-generation sequencing reveals wide genetic diversity of Blastocystis subtypes in chickens including potentially zoonotic subtypes. Parasitol. Res. 2021, 120, 2219–2231. [Google Scholar] [CrossRef] [PubMed]
- Maloney, J.G.; Jang, Y.; Molokin, A.; George, N.S.; Santin, M. Wide genetic diversity of Blastocystis in white-tailed deer (Odocoileus virginianus) from Maryland, USA. Microorganisms 2021, 9, 1343. [Google Scholar] [CrossRef] [PubMed]
- Higuera, A.; Herrera, G.; Jimenez, P.; Garcia-Corredor, D.; Pulido-Medellin, M.; Bulla-Castaneda, D.M.; Pinilla, J.C.; Moreno-Perez, D.A.; Maloney, J.G.; Santin, M.; et al. Identification of multiple Blastocystis subtypes in domestic animals from Colombia using amplicon-based next generation sequencing. Front. Vet. Sci. 2021, 8, 732129. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s box: Blastocystis subtypes revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Gantois, N.; Lamot, A.; Seesao, Y.; Creusy, C.; Li, L.L.; Monchy, S.; Benamrouz-Vanneste, S.; Karpouzopoulos, J.; Bourgain, J.L.; Rault, C.; et al. First report on the prevalence and subtype distribution of Blastocystis sp. in edible marine fish and marine mammals: A large-scale study conducted in Atlantic Northeast and on the coasts of Northern France. Microorganisms 2020, 8, 460. [Google Scholar] [CrossRef] [Green Version]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, J.D.; Sánchez, A.; Hernández, C.; Florez, C.; Bernal, M.C.; Giraldo, J.C.; Reyes, P.; Lopez, M.C.; Garcia, L.; Cooper, P.J.; et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 2016, 41, 32–35. [Google Scholar] [CrossRef]
- Khaled, S.; Gantois, N.; Ayoubi, A.; Even, G.; Sawant, M.; El Houmayraa, J.; Nabot, M.; Benamrouz-Vanneste, S.; Chabé, M.; Certad, G.; et al. Blastocystis sp. prevalence and subtypes distribution amongst Syrian refugee communities living in North Lebanon. Microorganisms 2021, 9, 184. [Google Scholar] [CrossRef] [PubMed]
- Hublin, J.S.Y.; Maloney, J.G.; Santin, M. Blastocystis in domesticated and wild mammals and birds. Res. Vet. Sci. 2021, 135, 260–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Owen, H.; Traub, R.J.; Cuttell, L.; Inpankaew, T.; Bielefeldt-Ohmann, H. Molecular epidemiology of Blastocystis in pigs and their in-contact humans in Southeast Queensland, Australia, and Cambodia. Vet. Parasitol. 2014, 203, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Greige, S.; El Safadi, D.; Bécu, N.; Gantois, N.; Pereira, B.; Chabé, M.; Benamrouz-Vanneste, S.; Certad, G.; El Hage, R.; Chemaly, M.; et al. Prevalence and subtype distribution of Blastocystis sp. isolates from poultry in Lebanon and evidence of zoonotic potential. Parasites Vectors 2018, 11, 389. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, C.S.; Efunshile, A.; Nelson, J.A.; Stensvold, C.R. Epidemiological aspects of Blastocystis colonization in children in Ilero, Nigeria. Am. J. Trop. Med. Hyg. 2016, 95, 175–179. [Google Scholar] [CrossRef] [Green Version]
- Greigert, V.; Abou-Bacar, A.; Brunet, J.; Nourrisson, C.; Pfaff, A.W.; Benarbia, L.; Pereira, B.; Randrianarivelojosia, M.; Razafindrakoto, J.-L.; Rakotomalala, R.S.; et al. Human intestinal parasites in Mahajanga, Madagascar: The kingdom of the protozoa. PLoS ONE 2018, 13, e0204576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, P.; Wawrzyniak, I.; Albert, A.; El Alaoui, H.; Delbac, F.; Livrelli, V. Development and evaluation of a real-time PCR assay for detection and quantification of Blastocystis parasites in human stool samples: Prospective study of patients with hematological malignancies. J. Clin. Microbiol. 2011, 49, 975–983. [Google Scholar] [CrossRef] [Green Version]
- D’Alfonso, R.; Santoro, M.; Essi, D.; Monsia, A.; Kaboré, Y.; Glé, C.; Di Cave, D.; Sorge, R.P.; Di Cristianzano, V.; Berrilli, F. Blastocystis in Côte d’Ivoire: Molecular identification and epidemiological data. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2243–2250. [Google Scholar] [CrossRef]
- Vennila, G.D.; Suresh, K.G. rregular shedding of B. hominis. Parasitol. Res. 1998, 85, 162–164. [Google Scholar] [CrossRef]
- Forsell, J.; Granlund, M.; Samuelsson, L.; Koskiniemi, S.; Edebro, H.; Evengard, B. High occurrence of Blastocystis sp. subtypes 1-3 and Giardia intestinalis assemblage B among patients in Zanzibar, Tanzania. Parasites Vectors 2016, 9, 370. [Google Scholar] [CrossRef] [Green Version]
- Lebba, V.; Santangelo, F.; Totino, V.; Pantanella, F.; Monsia, A.; Di Cristanziano, V.; Di Cave, D.; Schippa, S.; Berrilli, F.; D’Alfonso, R. Gut microbiota related to Giardia duodenalis, Entamoeba spp. and Blastocystis hominis infections in humans from Côte d’Ivoire. J. Infect. Dev. Ctries. 2016, 10, 1035–1041. [Google Scholar]
- Di Cristanziano, V.; Farowski, F.; Berrilli, F.; Santoro, M.; Di Cave, D.; Glé, C.; Daeumer, M.; Thielen, A.; Wirtz, M.; Kaiser, R.; et al. Analysis of human gut microbiota composition associated to the presence of commensal and pathogen microorganisms in Côte d’Ivoire. Microorganisms 2021, 9, 1763. [Google Scholar] [CrossRef] [PubMed]
- Cinek, O.; Polackova, K.; Odeh, R.; Alassaf, A.; Kramna, L.; Ibekwe, M.U.; Majaliwa, E.S.; Ahmadov, G.; Elmahi, B.M.E.; Mekki, H.; et al. Blastocystis in the faeces of children from six distant countries: Prevalence, quantity, subtypes and the relation to the gut bacteriome. Parasites Vectors 2021, 14, 399. [Google Scholar] [CrossRef]
- Kodio, A.; Coulibaly, D.; Kassoum Koné, A.; Konaté, S.; Doumbo, S.; Guindo, A.; Bittar, F.; Gouriet, F.; Raoult, D.; Aly Thera, M.; et al. Blastocystis colonization is associated with increased diversity and altered gut bacterial communities in healthy Malian children. Microorganisms 2019, 7, 649. [Google Scholar] [CrossRef] [Green Version]
- Dacal, E.; Saugar, J.M.; de Lucio, A.; Hernandez-de-Mingo, M.; Robinson, E.; Köster, P.C.; Aznar-Ruiz-de-Alegria, M.L.; Espasa, M.; Ninda, A.; Gandasegui, J.; et al. Prevalence and molecular characterization of Strongyloides stercoralis, Giardia duodenalis, Cryptosporidium spp., and Blastocystis spp. isolates in school children in Cubal, Western Angola. Parasites Vectors 2018, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muadica, A.S.; Köster, P.C.; Dashti, A.; Bailo, B.; Hernandez-de-Mingo, M.; Balasegaram, S.; Carmena, D. Molecular diversity of Giardia duodenalis, Cryptosporidium spp., and Blastocystis sp. in symptomatic and asymptomatic schoolchildren in Zambezia province (Mozambique). Pathogens 2021, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Parfrey, L.W.; Walters, W.A.; Lauber, C.L.; Clemente, J.C.; Berg-Lyons, D.; Teiling, C.; Kodira, C.; Mohiuddin, M.; Brunelle, J.; Driscoll, M.; et al. Communities of microbial eukaryotes in the mammalian gut within the context of environmental eukaryotic diversity. Front. Microbiol. 2014, 5, 298. [Google Scholar] [CrossRef]
- Bart, A.; Wentink-Bonnema, E.M.S.; Gilis, H.; Verhaar, N.; Wassenaar, C.J.A.; van Vugt, M.; Goorhuis, A.; Van Gool, T. Diagnosis and subtype analysis of Blastocystis sp. in 442 patients in a hospital setting in the Netherlands. BMC Infect. Dis. 2013, 13, 389. [Google Scholar] [CrossRef] [PubMed]
- Ben Abda, I.; Maatoug, N.; Ben Romdhane, R.; Bouhelmi, N.; Zallegua, N.; Aoun, K.; Viscogliosi, E.; Bouratbine, A. Prevalence and subtype identification of Blastocystis sp. in healthy individuals in the Tunis area, Tunisia. Am. J. Trp. Med. Hyg. 2017, 96, 202–204. [Google Scholar] [CrossRef] [Green Version]
- Abdulsalam, A.M.; Ithoi, I.; Al-Mekhlafi, H.M.; Al-Mekhlafi, A.M.; Ahmed, A.; Surin, J. Subtype distribution of Blastocystis isolates in Sebha, Libya. PLoS ONE 2013, 8, e84372. [Google Scholar] [CrossRef]
- El Deeb, H.K.; Khodeer, S. Blastocystis spp.: Frequency and subtype distribution in iron deficiency anemic versus non-anemic subjects from Egypt. J. Parasitol. 2013, 99, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Fouad, S.A.; Basyoni, M.M.A.; Fahmy, R.A.; Kobaisi, M.H. The pathogenic role of different Blastocystis hominis genotypes isolated from patients with irritable bowel syndrome. Arab J. Gastroenterol. 2011, 12, 194–200. [Google Scholar] [CrossRef]
- Hameed, D.M.A.; Hassanin, O.M.; Zuel-Fakkar, N.M. Association of Blastocystis hominis genetic subtypes with urticaria. Parasitol. Res. 2011, 108, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Souppart, L.; Moussa, H.; Cian, A.; Sanciu, G.; Poirier, P.; El Alaoui, H.; Delbac, F.; Boorom, K.; Delhaes, L.; Dei-Cas, E.; et al. Subtype analysis of Blastocystis isolates from symptomatic patients in Egypt. Parasitol. Res. 2010, 106, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Hussein, E.M.; Hussein, A.M.; Eida, M.M.; Atwa, M.M. Pathophysiological variability of different genotypes of human Blastocystis hominis Egyptian isolates in experimentally infected rats. Parasitol. Res. 2008, 102, 853–860. [Google Scholar] [CrossRef] [PubMed]
- El-Badry, A.A.; Abd El Wahab, W.M.; Hamdy, D.A.; Aboud, A. Blastocystis subtypes isolated from irritable bowel syndrome patients and co-infection with Helicobacter pylori. Parasitol. Res. 2018, 117, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, A.B.; Youssef, A.I. Subtype analysis of Blastocystis spp. isolated from domestic mammals and poultry and its relation to transmission to their in-contact humans in Ismailia governorate, Egypt. Parasitol. United J. 2018, 11, 90–98. [Google Scholar] [CrossRef]
- El Saftawy, E.A.; Amin, N.M.; Hamed, D.H.; Elkazazz, A. The hidden impact of different Blastocystis genotypes on C-3 and IgE serum levels: A matter of debate in asthmatic Egyptian children. J. Parasit. Dis. 2019, 43, 443–451. [Google Scholar] [CrossRef]
- Mokhtar, A.B.; Ahmed, S.A.; Eltamany, E.E.; Karanis, P. Anti-Blastocystis activity in vitro of Egyptian herbal extracts (Family: Asteraceae) with emphasis on Artemisia judaica. Int. J. Environ. Res. Public Health 2019, 16, 1555. [Google Scholar] [CrossRef] [Green Version]
- Mossallam, S.F.; El-Mansoury, S.A.T.; Tolba, M.M.; Kohla, A.A. In vitro susceptibility of human Blastocystis subtypes to simeprevir. Saudi J. Biol. Sci. 2021, 28, 2491–2501. [Google Scholar] [CrossRef]
- Abdo, S.M.; El-Adawy, H.; Farag, H.F.; El-Taweel, H.A.; Elhadad, H.; El-Badry, A.A.-M. Detection and molecular identification of Blastocystis isolates from humans and cattle in northern Egypt. J. Parasit. Dis. 2021, 45, 738–745. [Google Scholar] [CrossRef] [PubMed]
- Boutellis, A.; Aissi, M.; Harhoura, K.; Drali, R.; Kernif, T. First molecular characterization of Blastocystis subtypes from animals and animal-keepers stool in Algeria. Comp. Immunol. Microbiol. Infect. Dis. 2021, 78, 101695. [Google Scholar] [CrossRef] [PubMed]
- Petrasova, J.; Uzlikova, M.; Kostka, M.; Petrzelkova, K.J.; Huffman, M.A.; Modry, D. Diversity and host specificity of Blastocystis in syntopic primates on Rubondo Island, Tanzania. Int. J. Parasitol. 2011, 41, 1113–1120. [Google Scholar] [CrossRef]
- Belkessa, S.; Ait-Salem, E.; Laatamna, A.; Houali, K.; Sönksen, U.W.; Hakem, A.; Bouchene, Z.; Ghalmi, F.; Stensvold, C.R. Prevalence and clinical manifestations of Giardia intestinalis and other intestinal parasites in children and adults in Algeria. Am. J. Trop. Med. Hyg. 2021, 104, 910–916. [Google Scholar] [CrossRef]
- Sebaa, S.; Behnke, J.M.; Baroudi, D.; Hakem, A.; Abu-Madi, M. Prevalence and risk factors of intestinal protozoan infection among symptomatic and asymptomatic populations in rural and urban areas of southern Algeria. BMC Infect. Dis. 2021, 21, 888. [Google Scholar] [CrossRef] [PubMed]
- El Fatni, C.; Olmo, F.; El Fatni, H.; Romero, D.; Rosales, M.J. First genotyping of Giardia duodenalis and prevalence of enteroparasites in children from Tetouan (Morocco). Parasite 2014, 21, 48. [Google Scholar] [CrossRef] [PubMed]
- Graczyk, T.K.; Shiff, C.K.; Tamang, L.; Munsaka, F.; Beitin, A.M.; Moss, W.J. The association of Blastocystis hominis and Endolimax nana with diarrheal stools in Zambian school-age children. Parasitol. Res. 2005, 98, 38–43. [Google Scholar] [CrossRef]
- Zhu, W.; Tao, W.; Gong, B.; Yang, H.; Li, Y.; Song, M.; Lu, Y.; Li, W. First report of Blastocystis infections in cattle in China. Vet. Parasitol. 2017, 246, 38–42. [Google Scholar] [CrossRef]
- Greige, S.; El Safadi, D.; Khaled, S.; Gantois, N.; Baydoun, M.; Chemaly, M.; Benamrouz-Vanneste, S.; Chabé, M.; Osman, M.; Certad, G.; et al. First report on the prevalence and subtype distribution of Blastocystis sp. in dairy cattle in Lebanon and assessment of zoonotic transmission. Acta Trop. 2019, 194, 23–29. [Google Scholar] [CrossRef]
- Rauf-Adetotum, A.A.; Termizi, F.H.M.; Shaari, N.; Lee, I.L. The coexistence of Blastocystis spp. in humans, animals and environmental sources from 2010–2021 in Asia. Biology 2021, 10, 990. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Residence Place | Confessional Health Center Anastasis | National Hospital Ignace Deen | Total |
|---|---|---|---|
| Boffa | 0 | 1 | 1 |
| Conakry | 177 | 228 | 405 |
| Coyah | 0 | 2 | 2 |
| Dubréka | 0 | 12 | 12 |
| Kamsar | 0 | 2 | 2 |
| Kassa | 0 | 1 | 1 |
| Kindia | 72 | 2 | 74 |
| Labé | 0 | 1 | 1 |
| Nzérékoré | 1 | 0 | 1 |
| Tanéré | 0 | 1 | 1 |
| Total | 250 | 250 | 500 |
| Hospitals | Samples (n) | Positive Samples (n) | Prevalence(%) | Blastocystis sp. STs | |||||
|---|---|---|---|---|---|---|---|---|---|
| ST1 | ST2 | ST3 | ST4 | ST14 | MI a | ||||
| Confessional Health Center Anastasis | 250 | 194 | 77.6% | 62 | 27 | 49 | 1 | 7 | 48 |
| National Hospital Ignace Deen | 250 | 196 | 78.4% | 34 | 39 | 68 | 1 | 3 | 51 |
| Total | 500 | 390 | 78.0% | 96 | 66 | 117 | 2 | 10 | 99 |
| Residence Place | Samples (n) | Positive Samples (n) | Blastocystis sp. STs | |||||
|---|---|---|---|---|---|---|---|---|
| ST1 | ST2 | ST3 | ST4 | ST14 | MI a | |||
| Boffa | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Conakry | 405 | 314 | 69 | 54 | 100 | 1 | 10 | 80 |
| Coyah | 2 | 2 | 0 | 0 | 2 | 0 | 0 | 0 |
| Dubréka | 12 | 9 | 2 | 1 | 0 | 0 | 0 | 6 |
| Kamsar | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Kassa | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| Kindia | 74 | 59 | 23 | 11 | 13 | 1 | 0 | 11 |
| Labé | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Nzérékoré | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| Tanéré | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Total | 500 | 390 | 96 | 66 | 117 | 2 | 10 | 99 |
| African Region/Countries | Prevalence | Number of Subtyped Isolates | Subtyping Method | Blastocystis sp. STs | Mixed Infections d | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ST1 | ST2 | ST3 | ST4 | ST5 | ST6 | ST7 | ST10 | ST14 | ||||||
| North Africa | ||||||||||||||
| Tunisia | NA a | 61 | Sequencing | 18 | 10 | 31 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | [59] |
| Libya | 28.0% | 38 | Sequencing | 19 | 3 | 15 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | [39] |
| Libya | NA a | 48 | Sequencing | 26 | 13 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [60] |
| Egypt | NA a | 36 | PCR-STS b | 6 | 0 | 30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [61] |
| Egypt | NA a | 110 | PCR-STS b | 15 | 0 | 49 | 0 | 0 | 33 | 13 | 0 | 0 | 0 | [62] |
| Egypt | NA a | 33 | Sequencing | 0 | 0 | 33 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [63] |
| Egypt | NA a | 21 | Sequencing | 4 | 4 | 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [64] |
| Egypt | NA a | 44 | PCR-STS b | 8 | 0 | 24 | 0 | 0 | 8 | 4 | 0 | 0 | 0 | [65] |
| Egypt | NA a | 22 | Sequencing | 4 | 0 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [66] |
| Egypt | NA a | 53 | PCR-STS b | 16 | 4 | 30 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | [67] |
| Egypt | NA a | 100 | RFLP c | 0 | 0 | 84 | 16 | 0 | 0 | 0 | 0 | 0 | 0 | [68] |
| Egypt | NA a | 2 | PCR-STS b | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [69] |
| Egypt | NA a | 51 | PCR-STS | 9 | 2 | 40 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [70] |
| Egypt | NA a | 6 | Sequencing | 2 | 3 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [71] |
| Algeria | NA a | 3 | Sequencing | 0 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [72] |
| Total | 628 | 128 | 41 | 379 | 20 | 0 | 41 | 19 | 0 | 0 | 0 | |||
| West Africa | ||||||||||||||
| Nigeria | 84.0% | 127 | Sequencing | 51 | 42 | 33 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | [45] |
| Nigeria | 49.0% | 22 | Sequencing | 10 | 0 | 9 | 3 | 0 | 0 | 0 | 0 | 0 | 1 | [39] |
| Nigeria | 55.0% | 18 | Sequencing | 8 | 4 | 5 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | [53] |
| Côte d’Ivoire | 58.2% | 64 | Sequencing | 32 | 14 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [48] |
| Côte d’Ivoire | 70.0% | 0 | NA a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [51] |
| Côte d’Ivoire | 87.0% | 0 | NA a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [52] |
| Liberia | 70.0% | 25 | Sequencing | 7 | 7 | 8 | 3 | 0 | 0 | 0 | 0 | 0 | 5 | [39] |
| Senegal | 80.4% | 453 | Sequencing | 113 | 226 | 107 | 0 | 0 | 0 | 3 | 2 | 2 | 135 | [10] |
| Senegal | 100% | 103 | Sequencing | 29 | 21 | 51 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | [8] |
| Mali | 49.7% | 0 | NA a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [54] |
| Guinea | 78.0% | 291 | Sequencing | 96 | 66 | 117 | 2 | 0 | 0 | 0 | 0 | 10 | 99 | Present study |
| Total | 1103 | 346 | 380 | 348 | 10 | 0 | 1 | 4 | 2 | 12 | 240 | |||
| Central Africa | ||||||||||||||
| Angola | 25.6% | 75 | Sequencing | 23 | 23 | 27 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | [55] |
| Cameroon | 88.2% | 65 | Sequencing | 23 | 9 | 33 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [9] |
| Cameroon | 75.4% | 135 | Metagenomics | 45 | 32 | 57 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | [28] |
| Total | 275 | 91 | 64 | 117 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | |||
| East Africa | ||||||||||||||
| Tanzania | 81.8% | 34 | Metagenomics | 11 | 13 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [9] |
| Tanzania | 55.6% | 15 | Metagenomics | 1 | 12 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [24] |
| Tanzania | 60.9% | 92 | Sequencing | 36 | 28 | 27 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | [50] |
| Tanzania | NA a | 6 | Sequencing | 1 | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [73] |
| Tanzania | 53.0% | 8 | Sequencing | 2 | 2 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [53] |
| Sudan | 47.0% | 29 | Sequencing | 14 | 5 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [53] |
| Total | 184 | 65 | 63 | 55 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | |||
| Southern Africa | ||||||||||||||
| Madagascar | 64.5% | 158 | Sequencing | 80 | 36 | 42 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | [46] |
| Mozambique | 14.1% | 154 | Sequencing | 35 | 35 | 70 | 14 | 0 | 0 | 0 | 0 | 0 | 0 | [56] |
| Malawi | 69.6% | 25 | Metagenomics | 7 | 6 | 12 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | [57] |
| Total | 337 | 122 | 77 | 124 | 14 | 0 | 0 | 0 | 0 | 0 | 13 | |||
| Grand total | 2527 | 752 | 625 | 1023 | 45 | 1 | 42 | 25 | 2 | 12 | 253 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guilavogui, T.; Gantois, N.; Even, G.; Desramaut, J.; Dautel, E.; Denoyelle, C.; Cissé, F.I.; Touré, S.C.; Kourouma, B.L.; Sawant, M.; et al. Detection, Molecular Identification and Transmission of the Intestinal Protozoa Blastocystis sp. in Guinea from a Large-Scale Epidemiological Study Conducted in the Conakry Area. Microorganisms 2022, 10, 446. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020446
Guilavogui T, Gantois N, Even G, Desramaut J, Dautel E, Denoyelle C, Cissé FI, Touré SC, Kourouma BL, Sawant M, et al. Detection, Molecular Identification and Transmission of the Intestinal Protozoa Blastocystis sp. in Guinea from a Large-Scale Epidemiological Study Conducted in the Conakry Area. Microorganisms. 2022; 10(2):446. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020446
Chicago/Turabian StyleGuilavogui, Timothé, Nausicaa Gantois, Gaël Even, Jeremy Desramaut, Ellena Dautel, Constance Denoyelle, Fode Ibrahima Cissé, Salif Cherif Touré, Bakary Luther Kourouma, Manasi Sawant, and et al. 2022. "Detection, Molecular Identification and Transmission of the Intestinal Protozoa Blastocystis sp. in Guinea from a Large-Scale Epidemiological Study Conducted in the Conakry Area" Microorganisms 10, no. 2: 446. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020446