
The Response Regulator FlmD Regulates Biofilm Formation in Comamonas testosteroni through the Transcriptional Activator SoxR

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Transcriptome Analysis by RNA-Seq
2.3. Reverse Transcription PCR (RT-PCR) and Quantitative Reverse Transcription PCR (RT-qPCR)
2.4. Biofilm Formation Assays and Growth Measurement
2.5. Genetic Cloning, Overexpression, and Protein Purification
2.6. Gel Shift Assay (EMSA)
2.7. Protein Co-Purification
2.8. Bacterial Two-Hybrid Assay
2.9. Sequence Alignment and Analysis
2.10. Statistical Analysis
3. Results
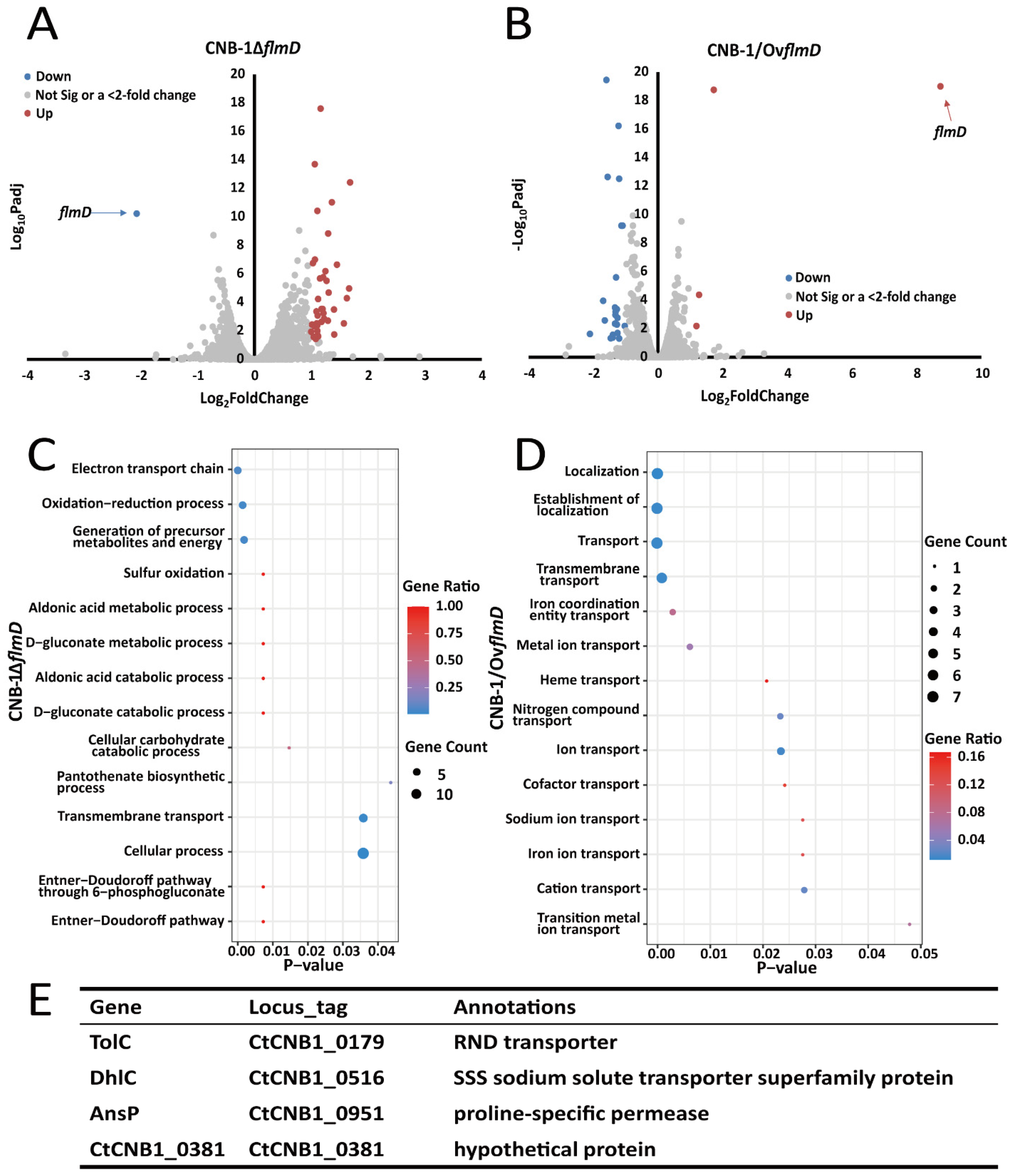
3.1. Certain Transporter Genes Regulated by FlmD
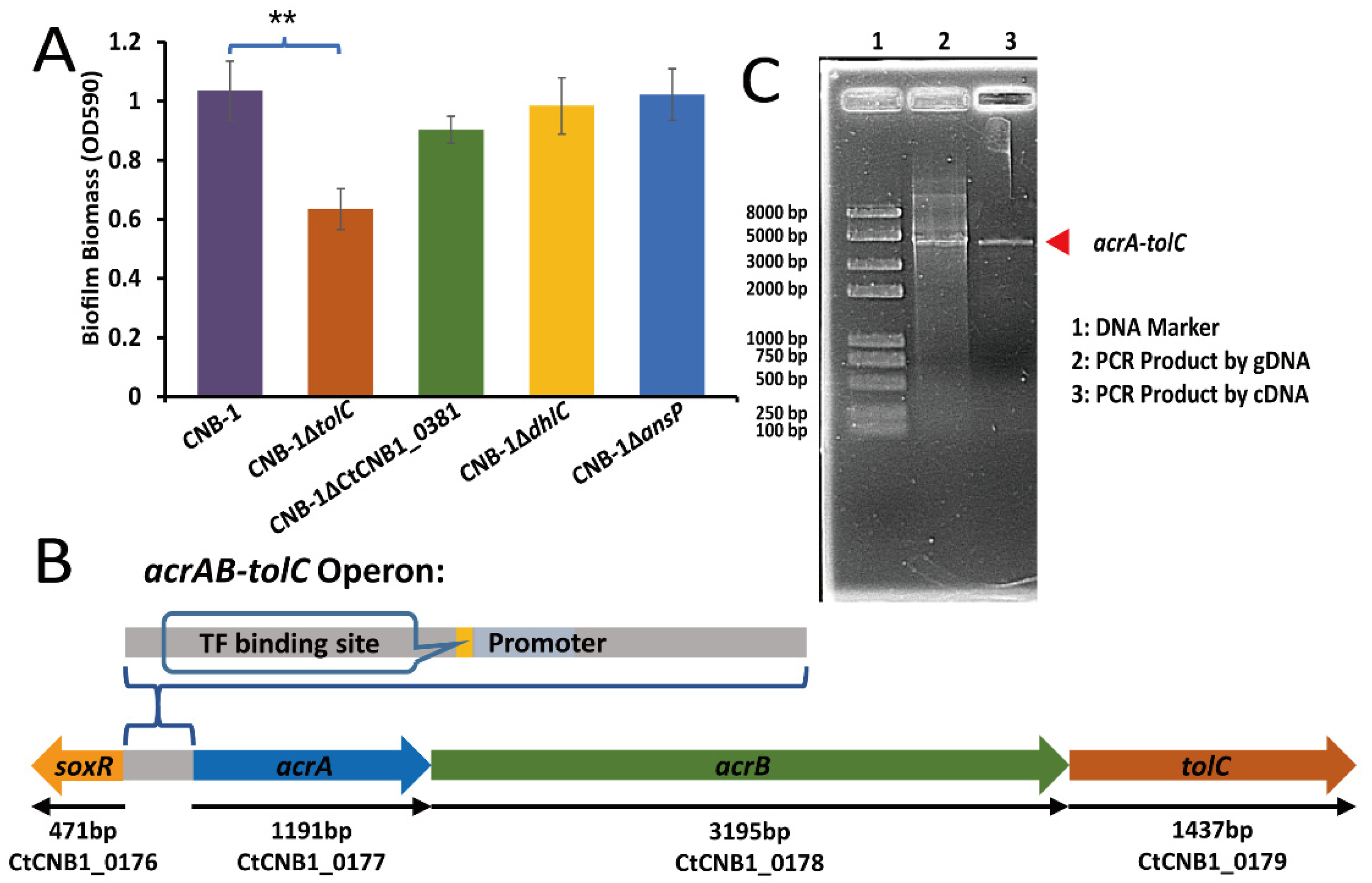
3.2. The Effect of TolC on Biofilm Formation
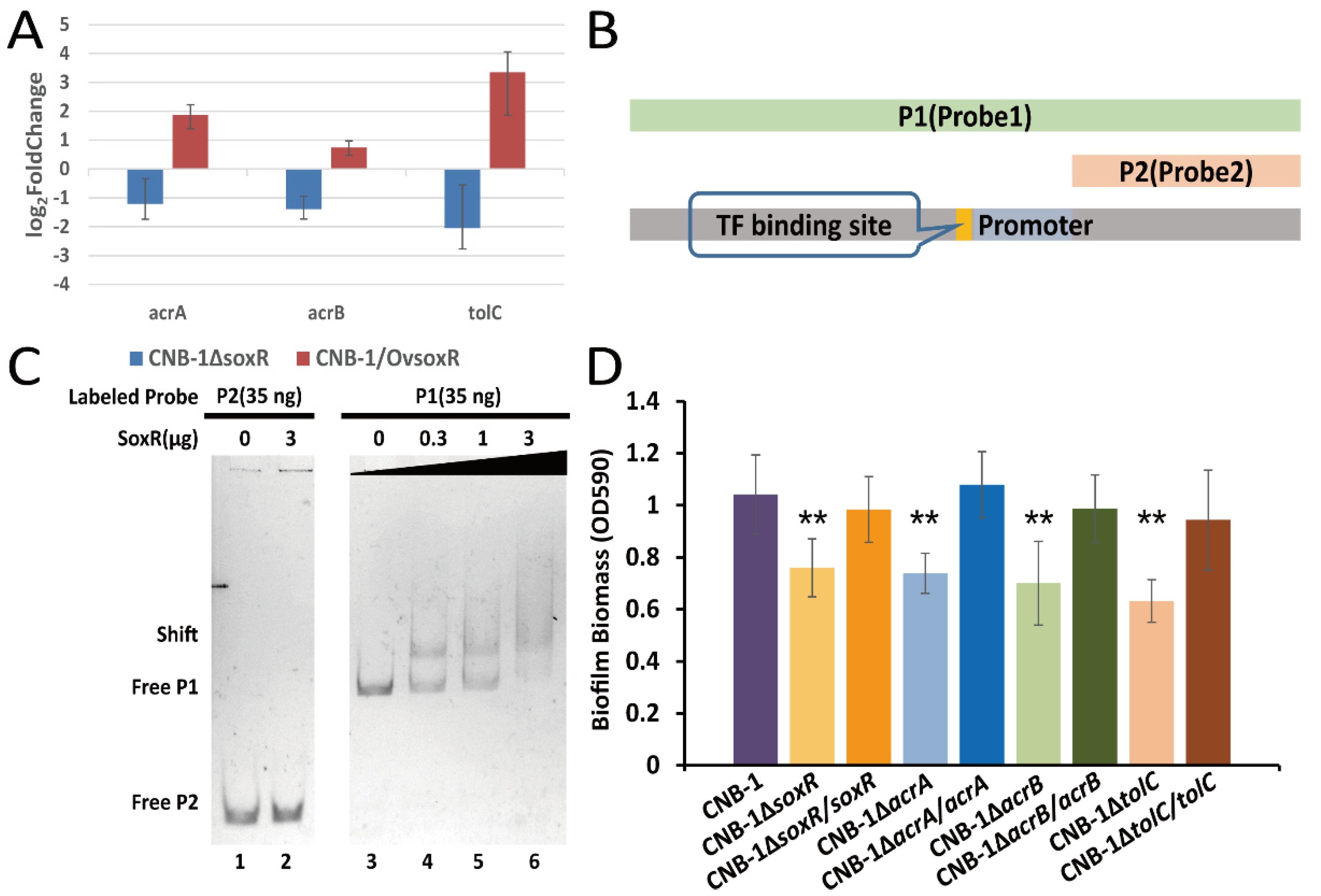
3.3. The Transcriptional Regulation of SoxR on the acrAB-tolC Operon
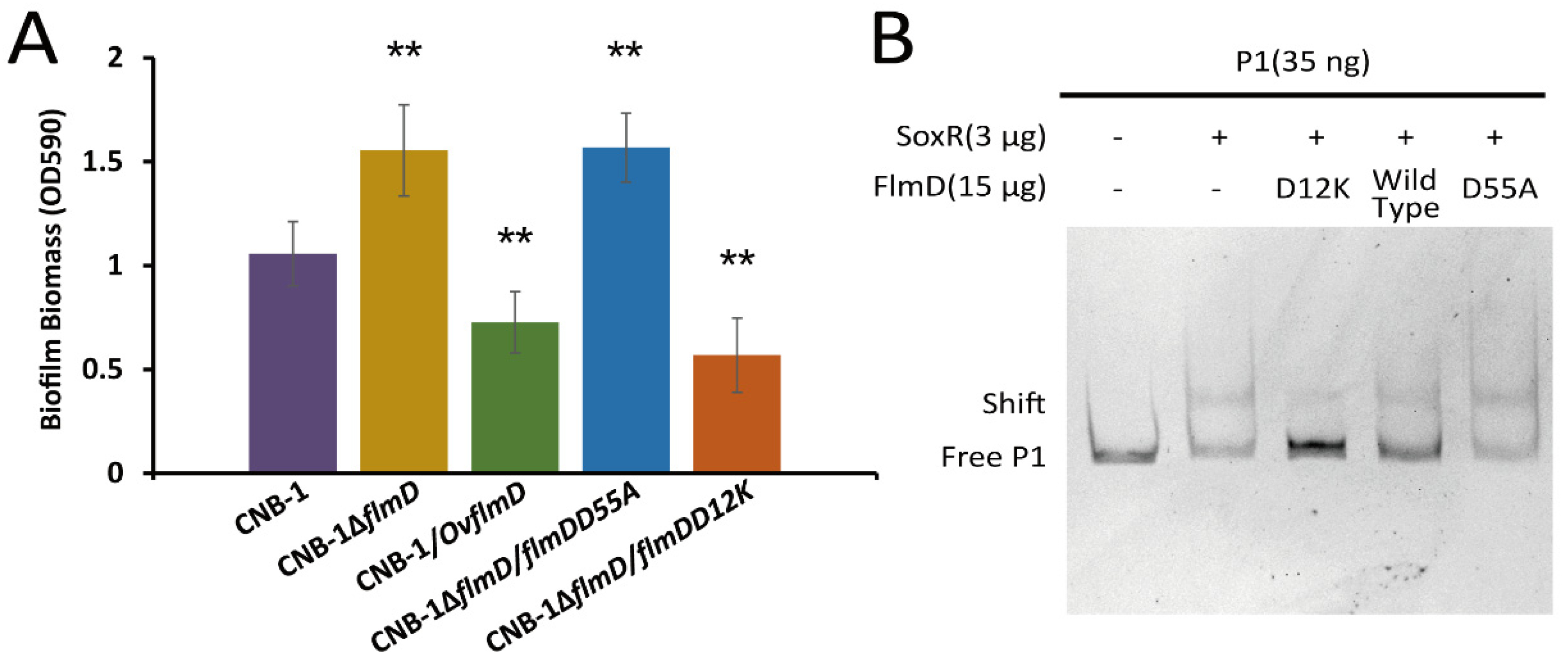
3.4. FlmD Variants Involved in Phosphorylation
3.5. The Interaction between FlmD and SoxR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Masak, J.; Cejkova, A.; Schreiberova, O.; Rezanka, T. Pseudomonas biofilms: Possibilities of their control. FEMS Microbiol. Ecol. 2014, 89, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Mah, T.F.; Pitts, B.; Pellock, B.; Walker, G.C.; Stewart, P.S.; O’Toole, G.A. A genetic basis for Pseudomonas aeruginosa biofilm antibiotic resistance. Nature 2003, 426, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Mah, T.F.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Koshland, D.E. Response regulator model in a simple sensory system. Science 1977, 196, 1055–1063. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, Y.H.; Zhu, H.Z.; Andrianova, E.P.; Jiang, C.Y.; Li, D.F.; Ma, L.Y.; Feng, J.; Liu, Z.P.; Xiang, H.; et al. Cross talk between chemosensory pathways that modulate chemotaxis and biofilm formation. mBio 2019, 10, e02876-18. [Google Scholar] [CrossRef] [Green Version]
- Moine, A.; Agrebi, R.; Espinosa, L.; Kirby, J.R.; Zusman, D.R.; Mignot, T.; Mauriello, E.M. Functional organization of a multimodular bacterial chemosensory apparatus. PLoS Genet. 2014, 10, e1004164. [Google Scholar] [CrossRef] [Green Version]
- Gumerov, V.M.; Andrianova, E.P.; Zhulin, I.B. Diversity of bacterial chemosensory systems. Curr. Opin. Microbiol. 2021, 61, 42–50. [Google Scholar] [CrossRef]
- Wadhams, G.H.; Armitage, J.P. Making sense of it all: Bacterial chemotaxis. Nat. Rev. Mol. Cell Biol. 2004, 5, 1024–1037. [Google Scholar] [CrossRef]
- Matilla, M.A.; Martin-Mora, D.; Gavira, J.A.; Krell, T. Pseudomonas aeruginosa as a model to study chemosensory pathway signaling. Microbiol. Mol. Biol. Rev. 2021, 85, e00151-20. [Google Scholar] [CrossRef] [PubMed]
- Wuichet, K.; Zhulin, I.B. Origins and diversification of a complex signal transduction system in prokaryotes. Sci. Signal. 2010, 3, ra50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.H.; Huang, Z.; Liu, S.J. Chemotaxis towards aromatic compounds: Insights from Comamonas testosteroni. Int. J. Mol. Sci. 2019, 20, 2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Zhu, W.T.; Cao, Z.; Xu, B.A.; Wang, G.J.; Luo, M.Z. High correlation between genotypes and phenotypes of environmental bacteria Comamonas testosteroni strains. BMC Genom. 2015, 16, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosaka, M.; Kamimura, N.; Toribami, S.; Mori, K.; Kasai, D.; Fukuda, M.; Masai, E. Novel tripartite aromatic acid transporter essential for terephthalate uptake in Comamonas sp. strain E6. Appl. Environ. Microbiol. 2013, 79, 6148–6155. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Jiang, C.Y.; Liu, X.Y.; Wu, J.F.; Han, J.G.; Liu, S.J. Plant-microbe association for rhizoremediation of chloronitroaromatic pollutants with Comamonas sp. strain CNB-1. Environ. Microbiol. 2007, 9, 465–473. [Google Scholar] [CrossRef]
- Kertesz, M.A. Riding the sulfur cycle--metabolism of sulfonates and sulfate esters in gram-negative bacteria. FEMS Microbiol. Rev. 2000, 24, 135–175. [Google Scholar]
- Arrigo, K.R. Marine microorganisms and global nutrient cycles. Nature 2005, 437, 349–355. [Google Scholar] [CrossRef]
- Boyd, C.D.; O’Toole, G.A. Second messenger regulation of biofilm formation: Breakthroughs in understanding c-di-GMP effector systems. Annu. Rev. Cell Dev. Biol. 2012, 28, 439–462. [Google Scholar] [CrossRef] [Green Version]
- Guvener, Z.T.; Harwood, C.S. Subcellular location characteristics of the Pseudomonas aeruginosa GGDEF protein, WspR, indicate that it produces cyclic-di-GMP in response to growth on surfaces. Mol. Microbiol. 2007, 66, 1459–1473. [Google Scholar] [CrossRef] [Green Version]
- Corral-Lugo, A.; De la Torre, J.; Matilla, M.A.; Fernandez, M.; Morel, B.; Espinosa-Urgel, M.; Krell, T. Assessment of the contribution of chemoreceptor-based signalling to biofilm formation. Environ. Microbiol. 2016, 18, 3355–3372. [Google Scholar] [CrossRef] [PubMed]
- Hickman, J.W.; Tifrea, D.F.; Harwood, C.S. A chemosensory system that regulates biofilm formation through modulation of cyclic diguanylate levels. Proc. Natl. Acad. Sci. USA 2005, 102, 14422–14427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitchurch, C.B.; Leech, A.J.; Young, M.D.; Kennedy, D.; Sargent, J.L.; Bertrand, J.J.; Semmler, A.B.T.; Mellick, A.S.; Martin, P.R.; Alm, R.A.; et al. Characterization of a complex chemosensory signal transduction system which controls twitching motility in Pseudomonas aeruginosa. Mol. Microbiol. 2004, 52, 873–893. [Google Scholar] [CrossRef] [PubMed]
- Fulcher, N.B.; Holliday, P.M.; Klem, E.; Cann, M.J.; Wolfgang, M.C. The Pseudomonas aeruginosa Chp chemosensory system regulates intracellular cAMP levels by modulating adenylate cyclase activity. Mol. Microbiol. 2010, 76, 889–904. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.F.; Jiang, C.Y.; Wang, B.J.; Ma, Y.F.; Liu, Z.P.; Liu, S.J. Novel partial reductive pathway for 4-chloronitrobenzene and nitrobenzene degradation in Comamonas sp. strain CNB-1. Appl. Environ. Microbiol. 2006, 72, 1759–1765. [Google Scholar] [CrossRef] [Green Version]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M., 2nd; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Chen, H.H.; Huang, Z.; Li, X.J.; Zhou, N.; Liu, C.; Jiang, C.Y.; Li, D.F.; Liu, S.J. PapA, a peptidoglycan-associated protein, interacts with OmpC and maintains cell envelope integrity. Environ. Microbiol. 2021, 23, 600–612. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Reese, M.G. Application of a time-delay neural network to promoter annotation in the Drosophila melanogaster genome. Comput. Chem. 2001, 26, 51–56. [Google Scholar] [CrossRef]
- Li, R.W. Metagenomics and Its Applications in Agriculture, Biomedicine, and Environmental Studies; Nova Science Publisher’s: Hauppauge, NY, USA, 2011; p. xi. 458p. [Google Scholar]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Du, D.; Wang, Z.; James, N.R.; Voss, J.E.; Klimont, E.; Ohene-Agyei, T.; Venter, H.; Chiu, W.; Luisi, B.F. Structure of the AcrAB-TolC multidrug efflux pump. Nature 2014, 509, 512–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zgurskaya, H.I.; Nikaido, H. Bypassing the periplasm: Reconstitution of the AcrAB multidrug efflux pump of Escherichia coli. Proc. Natl. Acad. Sci. USA 1999, 96, 7190–7195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donner, J.; Reck, M.; Bunk, B.; Jarek, M.; App, C.B.; Meier-Kolthoff, J.P.; Overmann, J.; Muller, R.; Kirschning, A.; Wagner-Dobler, I. The biofilm inhibitor carolacton enters gram-negative cells: Studies using a TolC-deficient strain of Escherichia coli. mSphere 2017, 2, e00375-17. [Google Scholar] [CrossRef] [Green Version]
- Subhadra, B.; Kim, J.; Kim, D.H.; Woo, K.; Oh, M.H.; Choi, C.H. Local repressor AcrR regulates AcrAB efflux pump required for biofilm formation and virulence in Acinetobacter nosocomialis. Front. Cell Infect. Microbiol. 2018, 8, 270. [Google Scholar] [CrossRef]
- Murakami, S.; Nakashima, R.; Yamashita, E.; Yamaguchi, A. Crystal structure of bacterial multidrug efflux transporter AcrB. Nature 2002, 419, 587–593. [Google Scholar] [CrossRef]
- Sa-Nogueira, I.; Mota, L.J. Negative regulation of L-arabinose metabolism in Bacillus subtilis: Characterization of the araR (araC) gene. J. Bacteriol. 1997, 179, 1598–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, F.; Monod, J. Genetic regulatory mechanisms in the synthesis of proteins. J. Mol. Biol. 1961, 3, 318–356. [Google Scholar] [CrossRef]
- Jiang, M.; Bourret, R.B.; Simon, M.I.; Volz, K. Uncoupled phosphorylation and activation in bacterial chemotaxis. The 2.3 A structure of an aspartate to lysine mutant at position 13 of CheY. J. Biol. Chem. 1997, 272, 11850–11855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alon, U.; Camarena, L.; Surette, M.G.; Aguera y Arcas, B.; Liu, Y.; Leibler, S.; Stock, J.B. Response regulator output in bacterial chemotaxis. EMBO J. 1998, 17, 4238–4248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galperin, M.Y. A census of membrane-bound and intracellular signal transduction proteins in bacteria: Bacterial IQ, extroverts and introverts. BMC Microbiol. 2005, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.T.; Bavro, V.N.; Barrera, N.P.; Frankish, H.M.; Velamakanni, S.; van Veen, H.W.; Robinson, C.V.; Borges-Walmsley, M.I.; Walmsley, A.R. MacB ABC transporter is a dimer whose ATPase activity and macrolide-binding capacity are regulated by the membrane fusion protein MacA. J. Biol. Chem. 2009, 284, 1145–1154. [Google Scholar] [PubMed] [Green Version]
- Tanabe, M.; Szakonyi, G.; Brown, K.A.; Henderson, P.J.; Nield, J.; Byrne, B. The multidrug resistance efflux complex, EmrAB from Escherichia coli forms a dimer in vitro. Biochem Biophys Res. Commun 2009, 380, 338–342. [Google Scholar] [CrossRef]
- Karatan, E.; Watnick, P. Signals, regulatory networks, and materials that build and break bacterial biofilms. Microbiol. Mol. Biol. Rev. 2009, 73, 310–347. [Google Scholar] [CrossRef] [Green Version]
- Van Wolferen, M.; Orell, A.; Albers, S.V. Archaeal biofilm formation. Nat. Rev. Microbiol. 2018, 16, 699–713. [Google Scholar] [CrossRef]
- Nagar, E.; Schwarz, R. To be or not to be planktonic? Self-inhibition of biofilm development. Environ. Microbiol. 2015, 17, 1477–1486. [Google Scholar] [CrossRef]
- Ortega, A.; Zhulin, I.B.; Krell, T. Sensory repertoire of bacterial chemoreceptors. Microbiol. Mol. Biol. Rev. 2017, 81, e00033-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, B.; Huang, Z.; Fan, Z.; Jiang, C.Y.; Liu, S.J. Comamonas testosteroni uses a chemoreceptor for tricarboxylic acid cycle intermediates to trigger chemotactic responses towards aromatic compounds. Mol. Microbiol. 2013, 90, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Vuoristo, K.S.; Mars, A.E.; Sanders, J.P.M.; Eggink, G.; Weusthuis, R.A. Metabolic engineering of TCA cycle for production of chemicals. Trends Biotechnol. 2016, 34, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Shanks, R.M.Q.; Meehl, M.A.; Brothers, K.M.; Martinez, R.M.; Donegan, N.P.; Graber, M.L.; Cheung, A.L.; O’Toole, G.A. Genetic evidence for an alternative citrate-dependent biofilm formation pathway in Staphylococcus aureus that is dependent on fibronectin binding proteins and the GraRS two-component regulatory system. Infect. Immun. 2008, 76, 2469–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, T.; Kobayashi, K. Role of glutamate synthase in biofilm formation by Bacillus subtilis. J. Bacteriol. 2020, 202, e00120-20. [Google Scholar] [CrossRef] [PubMed]
- Tatke, G.; Kumari, H.; Silva-Herzog, E.; Ramirez, L.; Mathee, K. Pseudomonas aeruginosa MifS-MifR two-component system is specific for alpha-ketoglutarate utilization. PLoS ONE 2015, 10, e0129629. [Google Scholar] [CrossRef] [PubMed]
- Orillard, E.; Watts, K.J. Deciphering the Che2 chemosensory pathway and the roles of individual Che2 proteins from Pseudomonas aeruginosa. Mol. Microbiol. 2021, 115, 222–237. [Google Scholar] [CrossRef]
- Zusman, D.R.; Scott, A.E.; Yang, Z.; Kirby, J.R. Chemosensory pathways, motility and development in Myxococcus xanthus. Nat. Rev. Microbiol. 2007, 5, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Ortega, D.R.; Yang, W.; Subramanian, P.; Mann, P.; Kjaer, A.; Chen, S.; Watts, K.J.; Pirbadian, S.; Collins, D.A.; Kooger, R.; et al. Repurposing a chemosensory macromolecular machine. Nat. Commun. 2020, 11, 2041. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Plasmid | Relevant Genotype or Description | Sources |
|---|---|---|
| Strains | ||
| Comamonas testosteroni | ||
| CNB-1 | [25] | |
| CNB-1ΔflmD | FlmD (CtCNB1_3988) disrupted in CNB-1 | [7] |
| CNB-1ΔdhlC | DhlC (CtCNB1_0516) disrupted in CNB-1 | This study |
| CNB-1ΔansP | AnsP (CtCNB1_0951) disrupted in CNB-1 | This study |
| CNB-1ΔCtCNB1_0381 | CtCNB1_0381 disrupted in CNB-1 | This study |
| CNB-1ΔacrA | AcrA (CtCNB1_0177) disrupted in CNB-1 | This study |
| CNB-1ΔacrB | AcrB (CtCNB1_0178) disrupted in CNB-1 | This study |
| CNB-1ΔtolC | TolC (CtCNB1_0179) disrupted in CNB-1 | This study |
| CNB-1ΔsoxR | SoxR (CtCNB1_0176) disrupted in CNB-1 | This study |
| Escherichia coli | ||
| DH5α | F-φ80d lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17(rK- mK+) supE44 λ- thi-1 gyrA96 relA1 phoA; host for DNA manipulations | TransGen |
| BL21(DE3) | F- ompT hsdS (rB- mB-) gal dcm (DE3) | Novagen |
| XL1-Blue | MRF’ Kan, glycerol stockb (host strain for propagating pBT and pTRG recombinants) | Stratagene |
| Plasmids | ||
| pBBR1MCS-2 | Kmr, lacPOZ’ broad host vector with R type conjugative origin | [26] |
| pBBR1MCS2-flmD | Carries flmD to generate complementation | [7] |
| pBBR1MCS2-flmDD55A | A mutation from an aspartate to an alanine in 55th residue | [7] |
| pBBR1MCS2-flmDD12K | A mutation from an aspartate to a lysine in 12th residue | This study |
| pBBR1MCS2-soxR | Carries soxR to generate complementation | This study |
| pBBR1MCS2-acrA | Carries acrA to generate complementation | This study |
| pBBR1MCS2-acrB | Carries acrB to generate complementation | This study |
| pBBR1MCS2-tolC | Carries tolC to generate complementation | This study |
| pBBR1MCS2-soxR-his | pBBR1MCS2 derivative for expression of SoxR | This study |
| pBBR1MCS2-flmDD12K-sterp + soxR-his | pBBR1MCS2 derivative for co-purification of SoxR and FlmDD12K | This study |
| pBBR1MCS2-flmDD55A-sterp + soxR-his | pBBR1MCS2 derivative for co-purification of SoxR and FlmDD55A | This study |
| pBBR1MCS2pfer | adds a strong constitutive promoter in pBBR1MCS-2 | [7] |
| pBBR1MCS2pfer-flmD | Carries flmD to overexpression | [7] |
| pET28a | Kmr, bacterial expression vector with a His-tag | Youbio |
| pET28a-flmD | pET28a derivative for expression of FlmD | [7] |
| pET28a-flmDD55A | pET28a derivative for expression of FlmD with D55A mutation | This study |
| pET28a-flmDD12K | pET28a derivative for expression of FlmD with D12K mutation | This study |
| pBT | Bacterial two-hybrid bait plasmid with λ repressor protein (λcI) | Stratagene |
| pBT-cheA | pBT derivative with λcI linked to CheA | [7] |
| pBT-flmDD12K | pBT derivative with λcI linked to FlmDD12K | This study |
| pBT-flmDD55A | pBT derivative with λcI linked to FlmDD55A | This study |
| pTRG | Bacterial two-hybrid bait plasmid with α-subunit of RNA polymerase (RNAp) | Stratagene |
| pTRG-cheW | pTRG derivative with RNAp linked to CheW | [7] |
| pTRG-soxR | pTRG derivative with RNAp linked to SoxR | This study |
| Gene | Locus_Tag | Annotations | Padj | log2FoldChange |
|---|---|---|---|---|
| hmp | CtCNB1_0015 | ferredoxin | 2.39 × 102 | 1.1233 |
| boxA | CtCNB1_0065 | benzoyl-CoA oxygenase | 3.01 × 103 | 1.5735 |
| boxB | CtCNB1_0066 | benzoyl-CoA oxygenase | 3.81 × 1013 | 1.6828 |
| boxC | CtCNB1_0067 | benzoyl-CoA-dihydrodiol lyase | 3.08 × 106 | 1.2693 |
| livK1 | CtCNB1_0096 | ABC transporter permease | 6.45 × 107 | 1.249 |
| bCL | CtCNB1_0097 | 4-hydroxybenzoate--CoA ligase | 2.09 × 106 | 1.1543 |
| tauA | CtCNB1_0160 | ABC transporter permease | 1.48 × 109 | 1.2986 |
| tauC | CtCNB1_0161 | ABC transporter permease | 8.38 × 104 | 1.1119 |
| acrA | CtCNB1_0177 | RND family efflux transporter MFP subunit | 9.97 × 108 | 1.0644 |
| tolC | CtCNB1_0179 | RND transporter | 1.74 × 106 | 1.2098 |
| CtCNB1_0381 | CtCNB1_0381 | hypothetical protein | 4.06 × 103 | 1.108 |
| caiD | CtCNB1_0392 | enoyl-CoA hydratase | 1.77 × 107 | 1.0308 |
| gpmB | CtCNB1_0417 | phosphoglycerate mutase | 2.67 × 102 | 1.0422 |
| dhlC | CtCNB1_0516 | SSS sodium solute transporter superfamily protein | 2.94 × 104 | 1.1811 |
| araJ | CtCNB1_0518 | major facilitator transporter | 3.17 × 104 | 1.4008 |
| ansP | CtCNB1_0951 | proline-specific permease | 5.09 × 105 | 1.6276 |
| dadA | CtCNB1_0952 | amino acid dehydrogenase | 4.35 × 104 | 1.0916 |
| livK2 | CtCNB1_1147 | twin-arginine translocation pathway signal protein | 1.94 × 103 | 1.2937 |
| eutG | CtCNB1_1590 | 4-hydroxybutyrate dehydrogenase | 2.33 × 103 | 1.1806 |
| CtCNB1_2363 | CtCNB1_2363 | hypothetical protein | 1.15 × 102 | 1.0003 |
| soxX | CtCNB1_2868 | SoxX protein | 3.36 × 102 | 1.0829 |
| soxA | CtCNB1_2869 | SoxA protein | 4.58 × 103 | 1.0581 |
| soxY | CtCNB1_2871 | twin-arginine translocation pathway signal | 2.94 × 104 | 1.1995 |
| soxD | CtCNB1_2872 | cytochrome C | 2.07 × 105 | 1.3067 |
| CtCNB1_3408 | CtCNB1_3408 | hypothetical protein | 2.02 × 102 | 1.1134 |
| dctP | CtCNB1_3427 | TRAP dicarboxylate transporter, DctP subunit | 2.54 × 1018 | 1.1641 |
| ilvD | CtCNB1_3428 | phosphogluconate dehydratase | 3.83 × 1011 | 1.1086 |
| eda | CtCNB1_3429 | keto-deoxy-phosphogluconate aldolase | 2.26 × 107 | 1.4534 |
| livK3 | CtCNB1_3479 | ABC transporter permease | 2.97 × 103 | 1.097 |
| sapF | CtCNB1_3509 | ABC-type antimicrobial peptide | 1.39 × 103 | 1.2249 |
| coxL | CtCNB1_3514 | carbon monoxide dehydrogenase | 9.86 × 103 | 1.1075 |
| coxS | CtCNB1_3515 | carbon monoxide dehydrogenase | 3.64 × 102 | 1.079 |
| ilvB | CtCNB1_4060 | thiamine pyrophosphate-requiring enzymes | 3.64 × 103 | 1.0155 |
| apbA | CtCNB1_4061 | 2-dehydropantoate 2-reductase | 1.80 × 102 | 1.4053 |
| tctC | CtCNB1_4499 | ABC transporter substrate-binding protein | 9.44 × 1012 | 1.3628 |
| CtCNB1_4500 | CtCNB1_4500 | hypothetical protein | 1.07 × 105 | 1.6667 |
| ubiH | CtCNB1_4501 | FAD-dependent oxidoreductase | 5.75 × 104 | 1.214 |
| thiJ | CtCNB1_4569 | putative intracellular protease/amidase | 2.03 × 1014 | 1.0617 |
| modA | CtCNB1_4661 | extracellular solute-binding protein | 5.80 × 105 | 1.1208 |
| Gene | Locus_Tag | Annotations | Padj | log2FoldChange |
|---|---|---|---|---|
| tolC | CtCNB1_0179 | RND transporter | 5.23 × 103 | −1.2708 |
| CtCNB1_0378 | CtCNB1_0378 | hypothetical protein | 2.67 × 103 | −1.6453 |
| CtCNB1_0381 | CtCNB1_0381 | hypothetical protein | 1.12 × 104 | −1.6966 |
| CtCNB1_0383 | CtCNB1_0383 | hypothetical protein | 4.46 × 103 | −1.3307 |
| dhlC | CtCNB1_0516 | SSS sodium solute transporter superfamily | 4.24 × 102 | −1.4034 |
| ansP | CtCNB1_0951 | proline-specific permease | 1.10 × 103 | −1.3189 |
| cirA | CtCNB1_1178 | TonB-dependent receptor | 6.02 × 1010 | −1.132 |
| nrdA | CtCNB1_1179 | ribonucleotide reductase | 2.33 × 102 | −2.1049 |
| hemS | CtCNB1_1180 | ribonucleotide reductase | 3.54 × 1020 | −1.5981 |
| chuT | CtCNB1_1181 | ABC transporter substrate-binding protein | 2.26 × 1013 | −1.5584 |
| fepD | CtCNB1_1182 | hemin transport system permease protein | 2.71 × 102 | −1.3963 |
| fepC | CtCNB1_1183 | ABC transporter | 4.39 × 104 | −1.2717 |
| fhuE | CtCNB1_1601 | ligand-gated channel protein | 1.73 × 103 | −1.2529 |
| cbiK | CtCNB1_1602 | TonB-dependent receptor | 2.04 × 102 | −1.2527 |
| CtCNB1_1891 | CtCNB1_1891 | hypothetical protein | 1.77 × 1019 | 1.7244 |
| CtCNB1_1892 | CtCNB1_1892 | hypothetical protein | 4.26 × 105 | 1.2684 |
| CtCNB1_2304 | CtCNB1_2304 | hypothetical protein | 6.55 × 103 | 1.1858 |
| oafA | CtCNB1_3421 | acyltransferase | 3.30 × 104 | −1.3331 |
| coxG | CtCNB1_3510 | carbon monoxide dehydrogenase subunit G | 4.97 × 102 | −1.2019 |
| hppD | CtCNB1_3836 | 4-hydroxyphenylpyruvate dioxygenase | 3.02 × 1013 | −1.2022 |
| CtCNB1_3837 | CtCNB1_3837 | hypothetical protein | 6.02 × 1010 | −1.0953 |
| lraI | CtCNB1_3842 | ABC-type metal ion transport system | 4.71 × 102 | −1.462 |
| CtCNB1_3869 | CtCNB1_3869 | hypothetical protein | 6.64 × 104 | −1.3152 |
| fepA | CtCNB1_4259 | TonB-dependent receptor | 6.55 × 103 | −1.0318 |
| livK1 | CtCNB1_4328 | ABC transporter permease | 5.94 × 1017 | −1.2227 |
| livK2 | CtCNB1_4497 | ABC transporter permease | 2.55 × 106 | −1.2985 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Huang, Z.; Zhou, N.; Liu, C.; Jiang, C.; Li, D.; Liu, S. The Response Regulator FlmD Regulates Biofilm Formation in Comamonas testosteroni through the Transcriptional Activator SoxR. Microorganisms 2022, 10, 356. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020356
Wang Y, Huang Z, Zhou N, Liu C, Jiang C, Li D, Liu S. The Response Regulator FlmD Regulates Biofilm Formation in Comamonas testosteroni through the Transcriptional Activator SoxR. Microorganisms. 2022; 10(2):356. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020356
Chicago/Turabian StyleWang, Yunhao, Zhou Huang, Nan Zhou, Chang Liu, Chengying Jiang, Defeng Li, and Shuangjiang Liu. 2022. "The Response Regulator FlmD Regulates Biofilm Formation in Comamonas testosteroni through the Transcriptional Activator SoxR" Microorganisms 10, no. 2: 356. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020356