
Unraveling the Uncharacterized Domain of Carocin S2: A Ribonuclease Pectobacterium carotovorum subsp. carotovorum Bacteriocin



 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
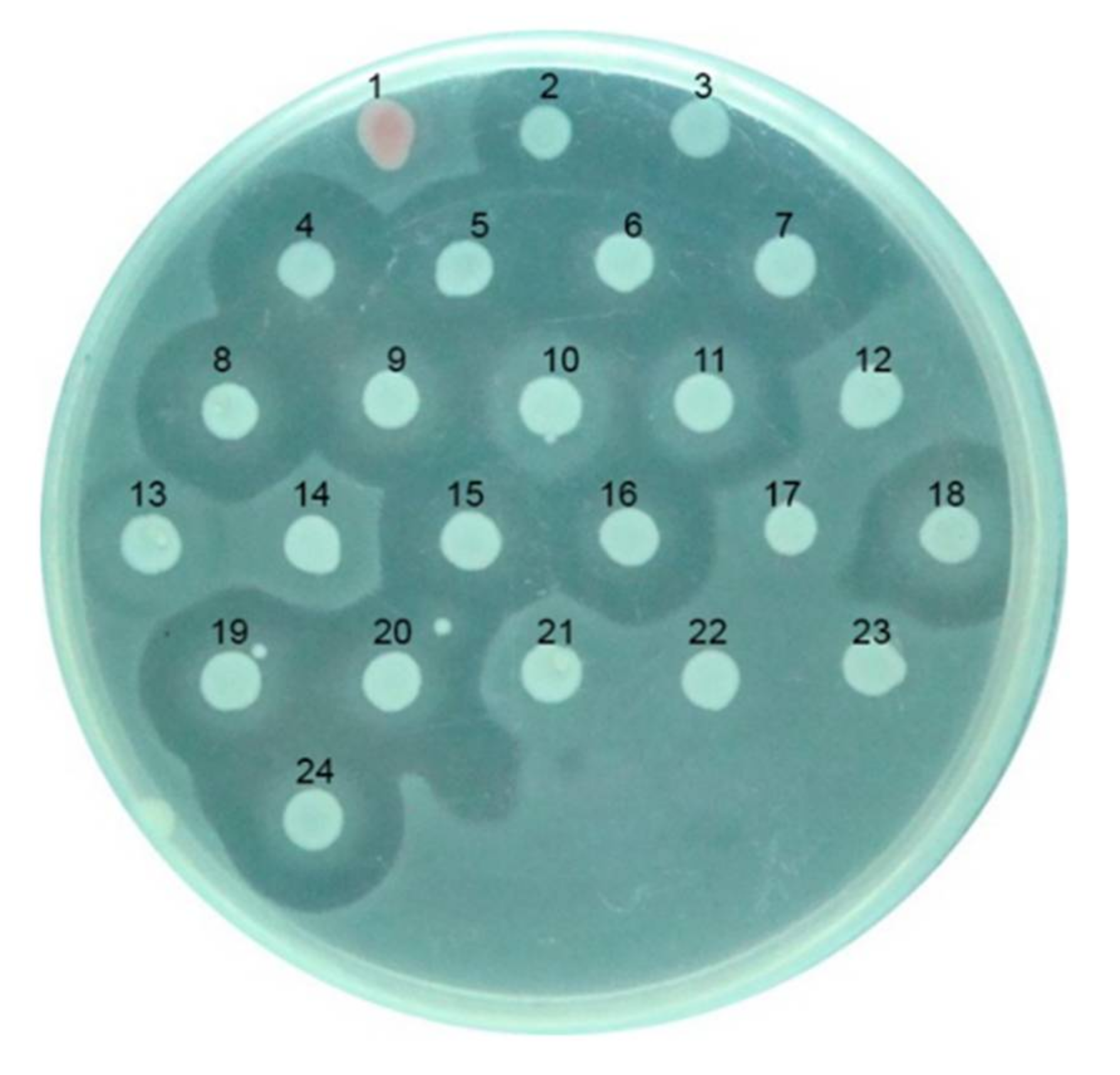
3.1. Characterization of the Killer Protein (CaroS2K) Using Structure-Based Mutagenesis
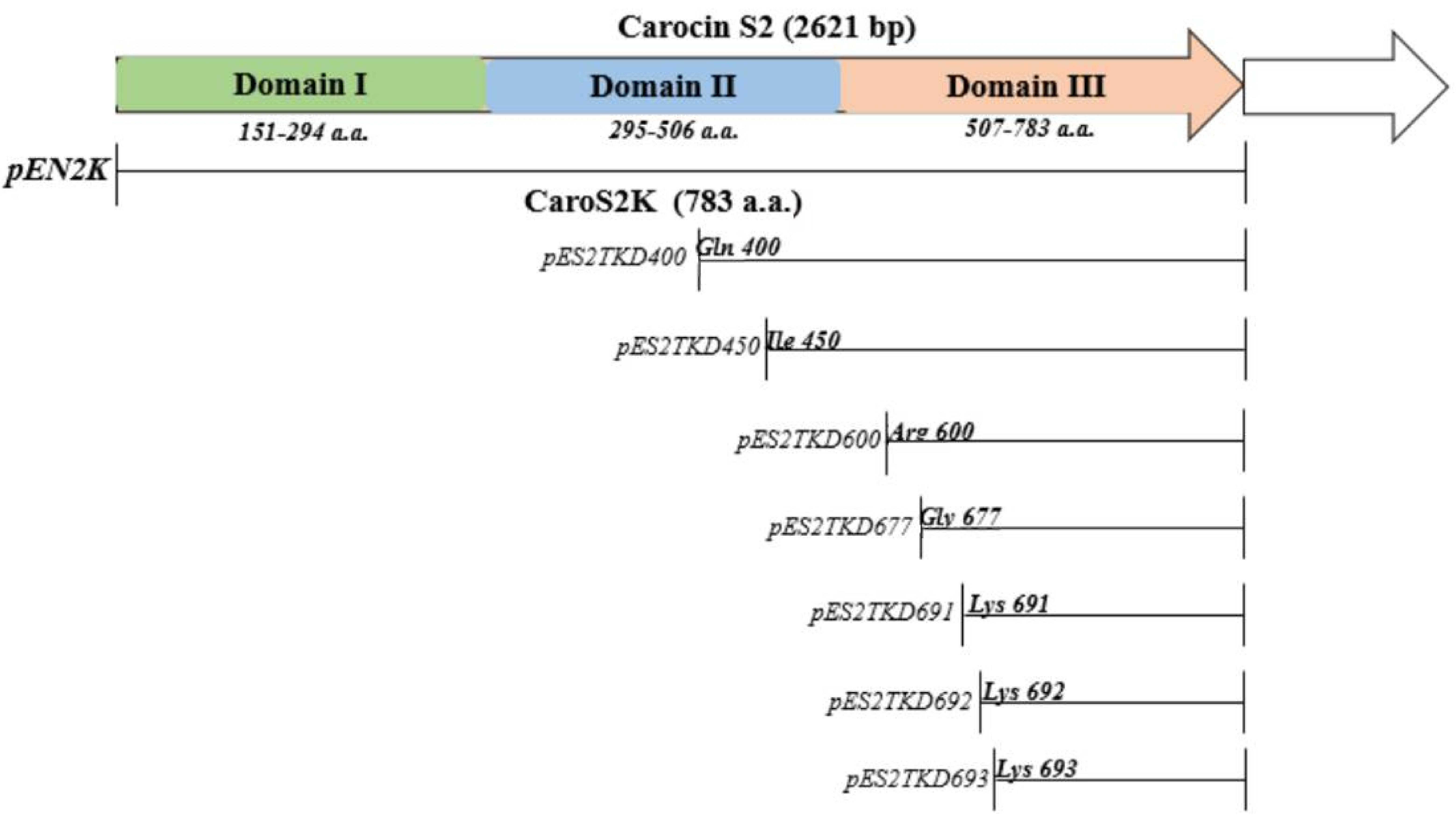
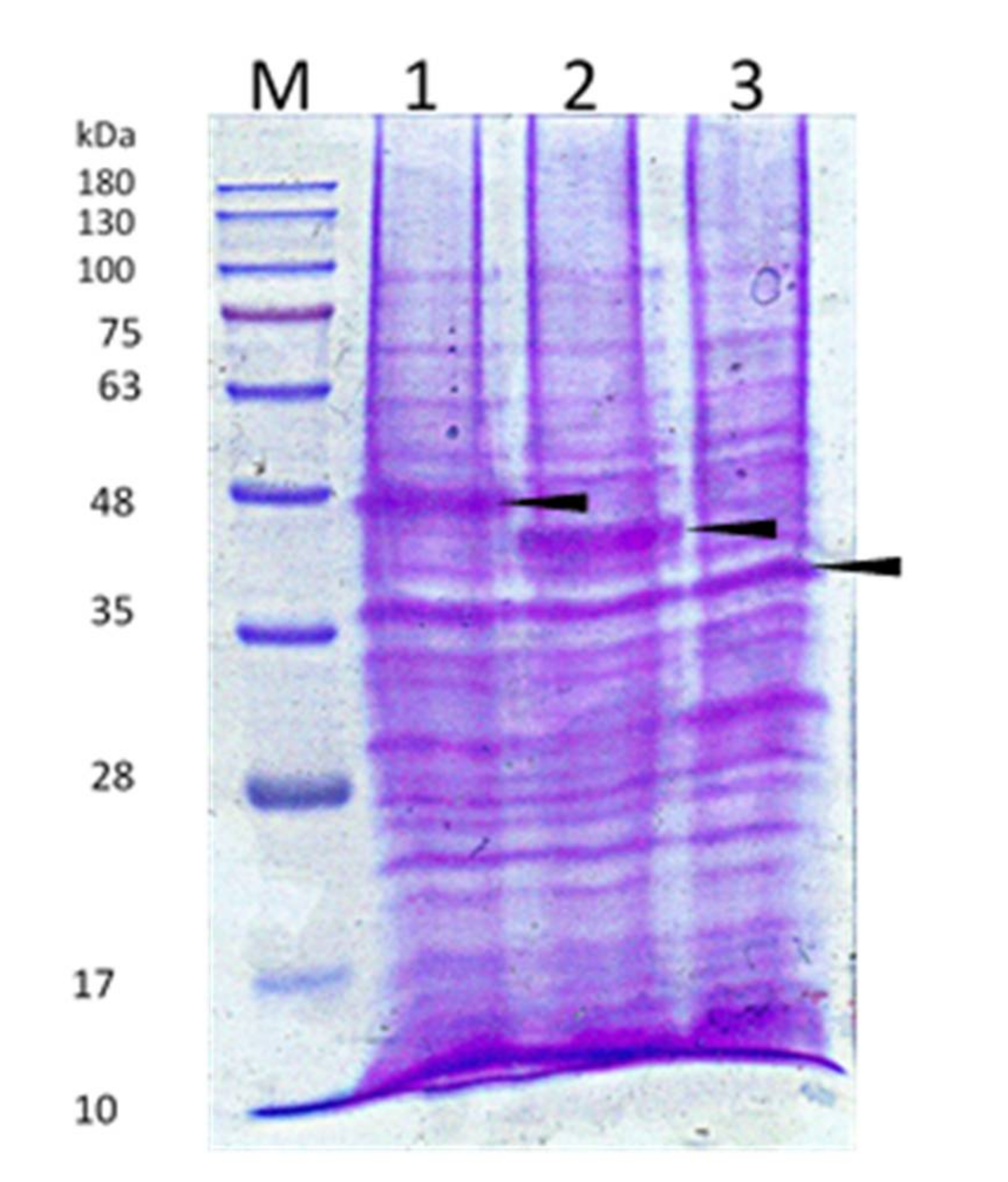
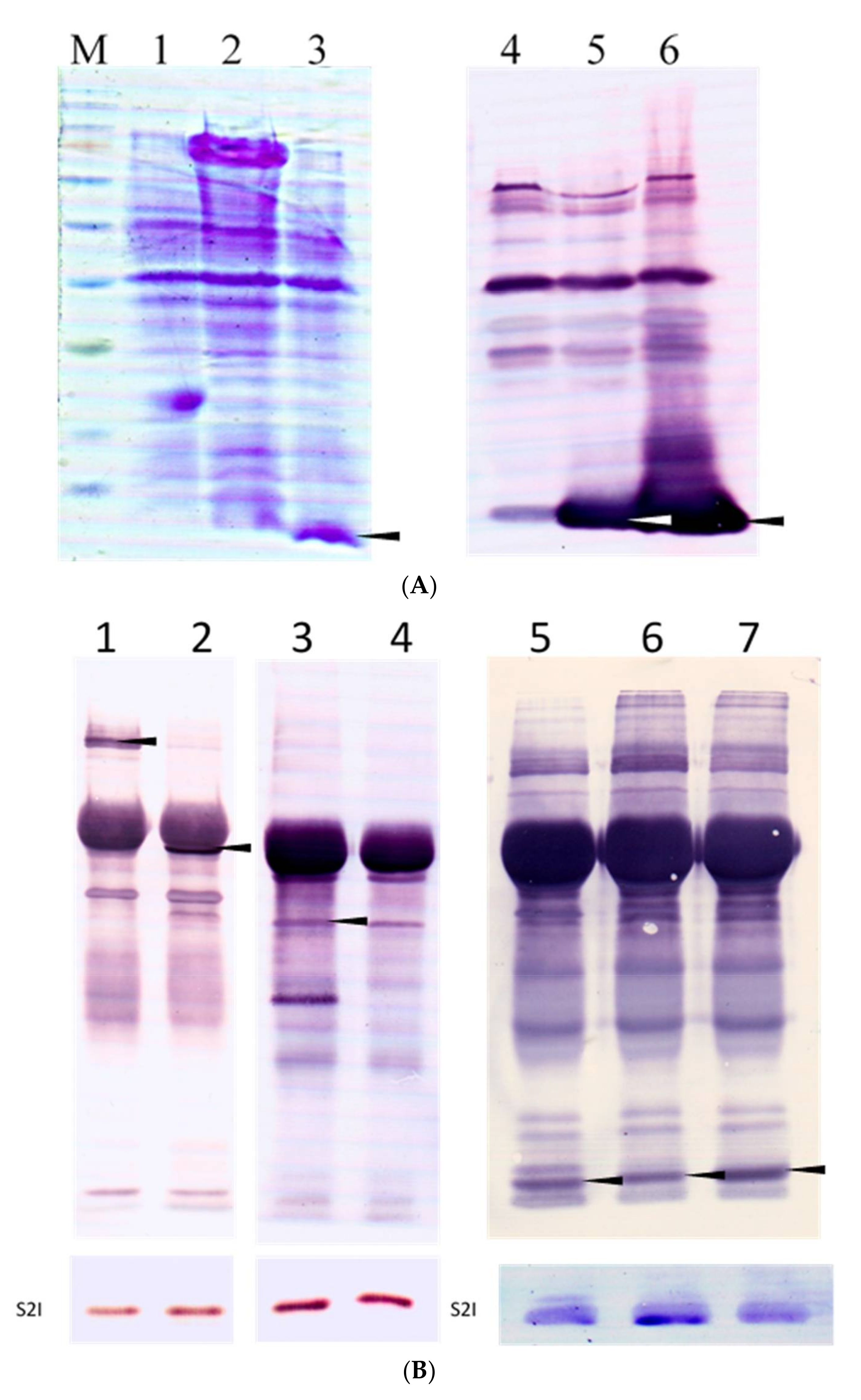
3.2. Truncated Bacteriocin Isolation Expression, and Purification
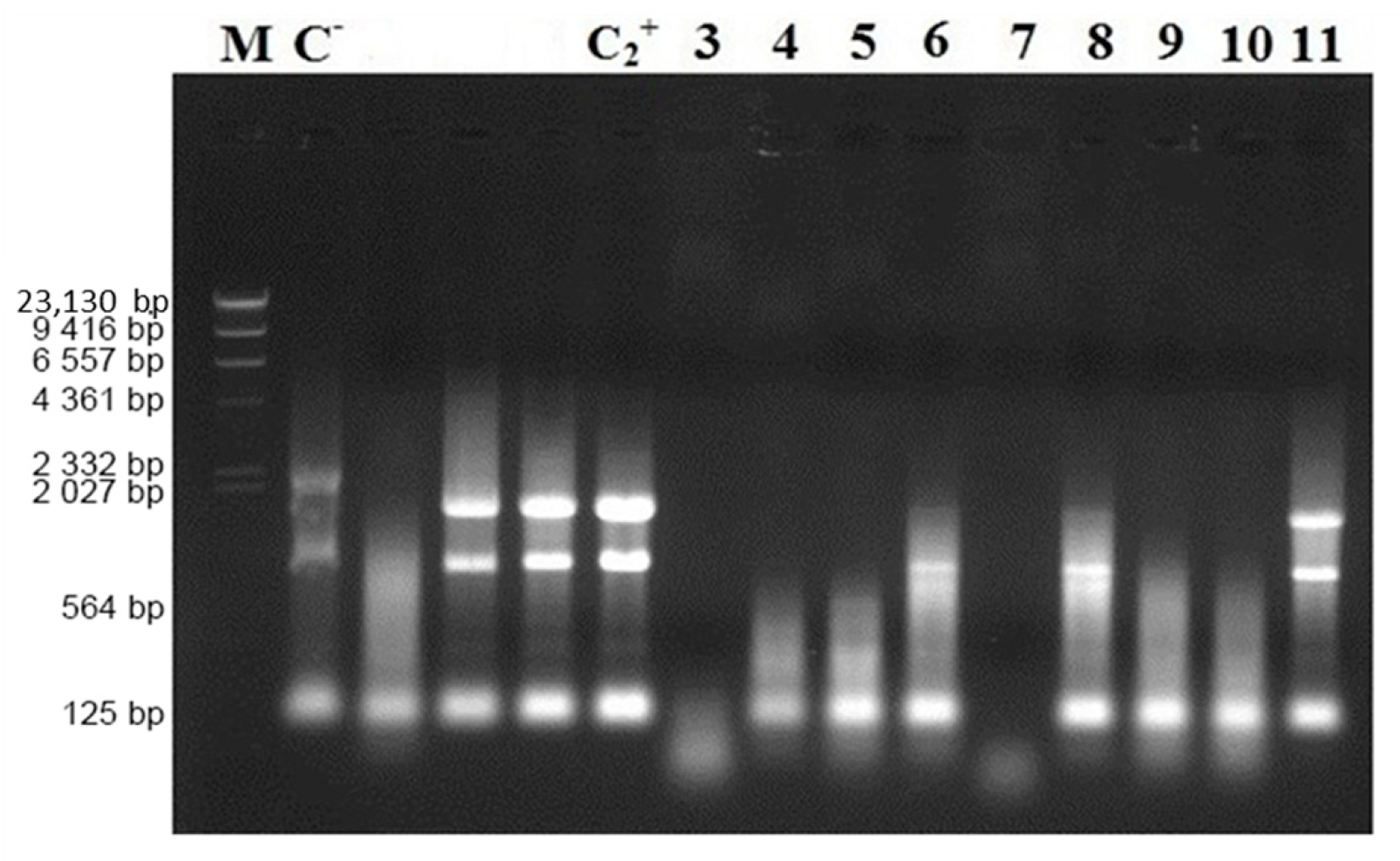
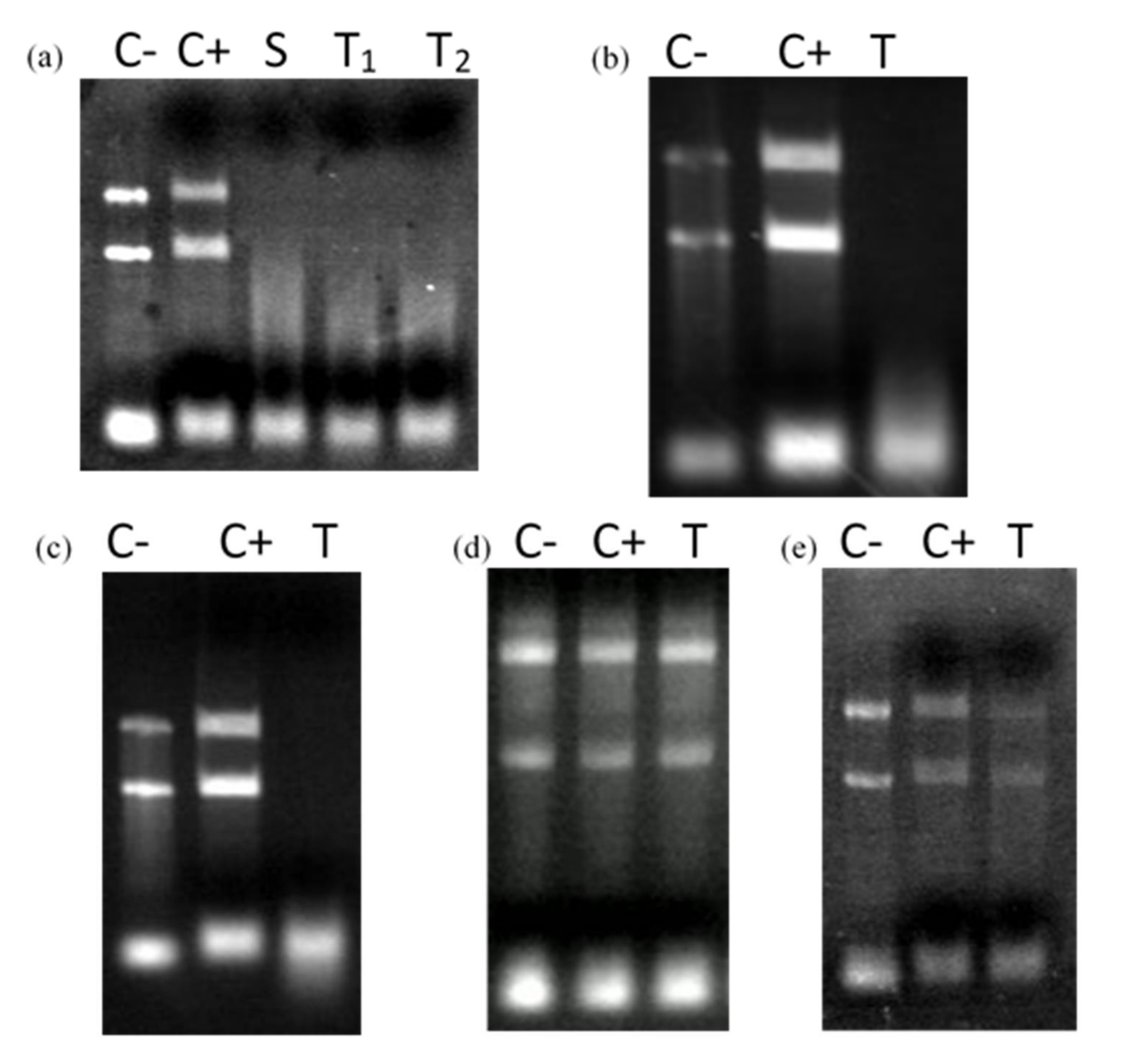
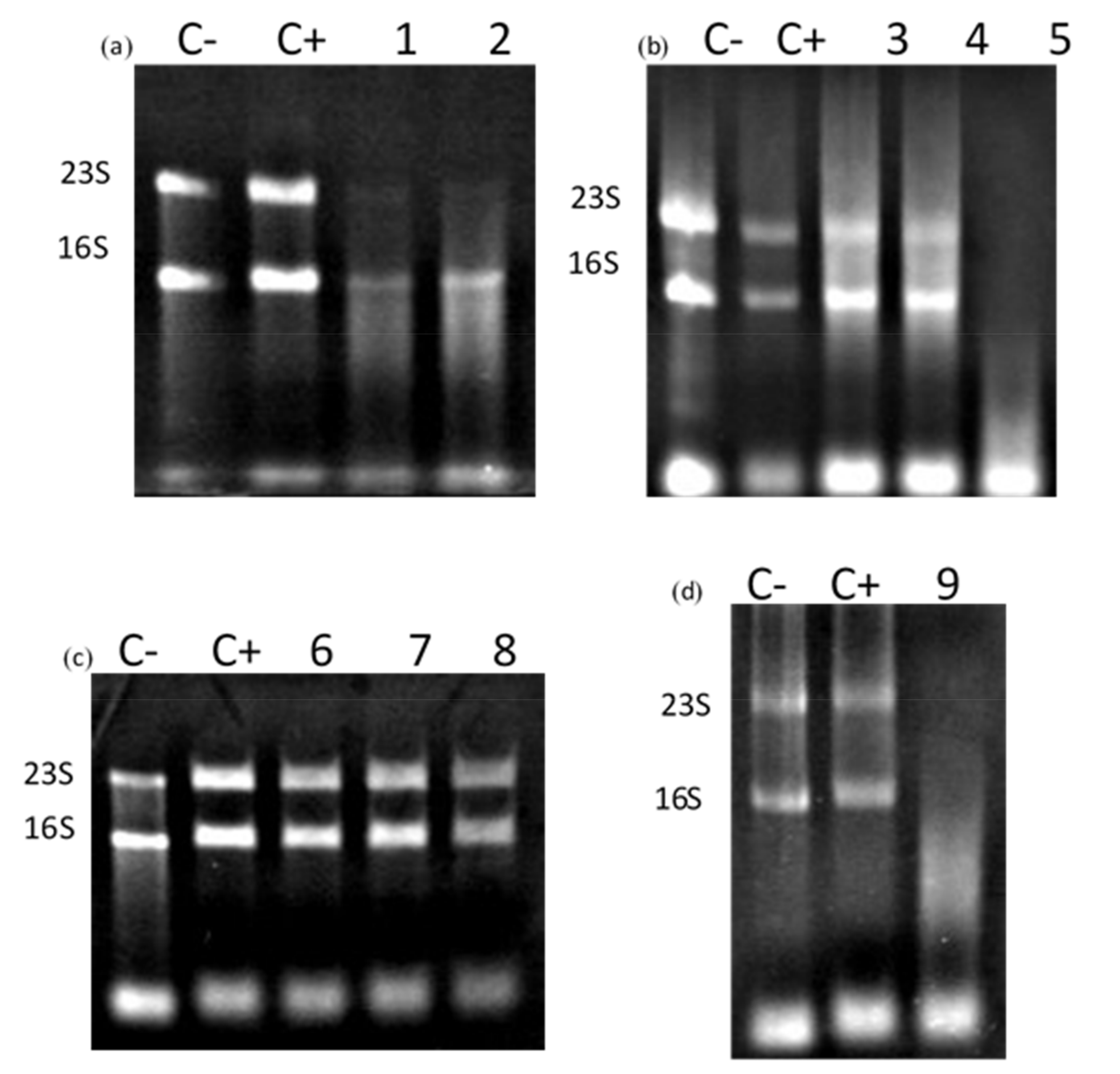
3.3. Truncated Bacteriocin CaroS2TKD Mutants Were Tested for Ribosomal RNA Hydrolysis Activity
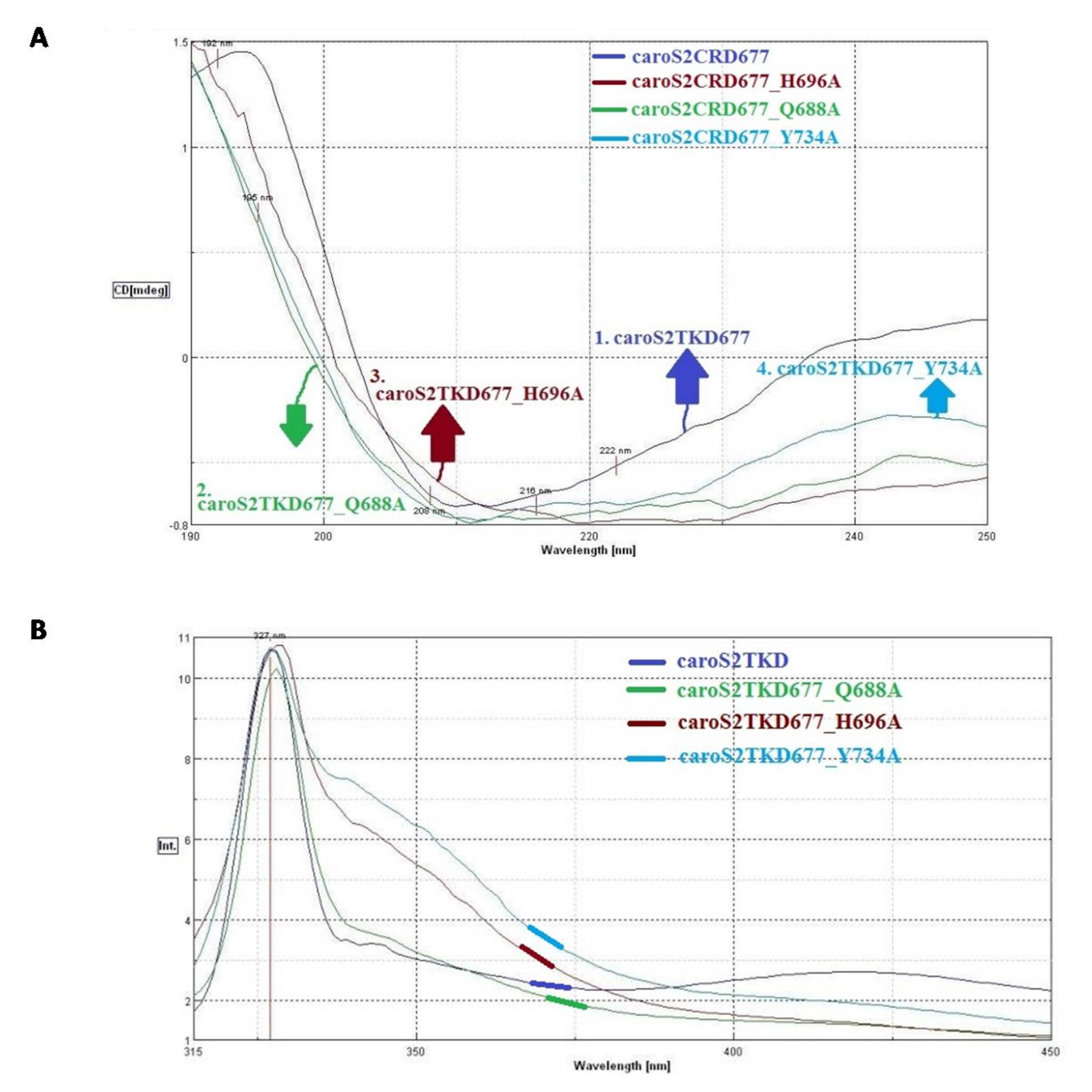
3.4. Structural Depiction of Bacteriocin, Carocin S2
3.5. The Binding Region of CaroS2K and Its Immunity Protein CaroS2I
4. Discussion
Proposed Catalytic Mechanism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Daw, M.A.; Falkiner, F.R. Bacteriocins: Nature, function and structure. Micron 1996, 27, 467–479. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M.T. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Martínez, B.; Rodríguez, A.; Suárez, E. Antimicrobial Peptides Produced by Bacteria: The bacteriocins. In New Weapons to Control. Bacterial Growth; Villa, T.G., Vinas, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, antimicrobial peptides from bacterial origin: Overview of their biology and their impact against multidrug-resistant bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Rea, M.C.; Sit, C.S.; Clayton, E.; O’Connor, P.M.; Whittal, R.M.; Zheng, J.; Vederas, J.C.; Ross, P.; Hill, C. Thuricin CD, a posttranslationally modified bacteriocin with a narrow spectrum of activity against Clostridium difficile. Proc. Natl. Acad. Sci. USA 2010, 107, 9352–9357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.C.; Tarr, A.W.; Penfold, C.N. Colicin import into E. coli cells: A model system for insights into the import mechanisms of bacteriocins. Biochim. Biophys. Acta 2014, 1843, 1717–1731. [Google Scholar] [CrossRef] [Green Version]
- Kleanthous, C. Swimming against the tide: Progress and challenges in our understanding of colicin translocation. Nat. Rev. Microbiol. 2010, 8, 843–848. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Ben Said, L.; Gaudreau, H.; Bedard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials:toxicity aspects and regulations. FEMS Microbio. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.A. Molecular mechanism bacteriocin evolution. Ann. Rev. Genet. 1998, 32, 255–278. [Google Scholar] [CrossRef] [Green Version]
- Atanaskovic, I.; Kleanthous, C. Tools and approaches for dissecting protein bacteriocin import in Gram-negative bacteria. Front. Microbiol. 2019, 10, 646. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic. Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Charkowski, A.O. The changing face of bacterial soft-rot diseases. Annu. Rev. Phytopathol. 2018, 56, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Marquez-Villavicencio, M.D.P.; Groves, R.L.; Charkowski, A.O. Soft Rot disease severity is affected by potato physiology and Pectobacterium taxa. Plant. Dis. 2011, 95, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derilo, R.C.; Wang, J.W.; Lagitnay, R.B.J.; Wu, H.P.; Chen, K.I.; Chuang, D.Y. Identification and characterization of the bacteriocin Carocin S3 from the multiple bacteriocin strain of Pectobacterium carotovorum subsp. carotovorum. BMC Microbiol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Chuang, D.Y.; Chien, Y.C.; Wu, H.P. Cloning and expression of the Erwinia carotovora subsp. carotovora gene encoding low-molecular weight bacteriocin Carocin S1. J. Bacteriol. 2007, 189, 620–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.-C.; Wu, J.-L.; Wu, H.-P.; Tzeng, K.-C.; Chuang, D.-Y. Cloning, purification, and functional characterization of Carocin S2, a ribonuclease bacteriocin produced by Pectobacterium carotovorum. BMC Microbiol. 2011, 11, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, E.; Park, T.-H.; Kim, M.-I.; Lee, S.; Ryu, S.; Oh, C.-S.; Rhee, S.; Kim, D.-H.; Part, B.-S.; Hue, S. Characterization of a new bacteriocin, Carocin D, from Pectobacterium carotovorum subsp. carotovorum Pcc21. Appl. Environ. Microbiol. 2010, 76, 7541–7549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinter, R.; Milner, J.; Walker, D. Ferredoxin containing bacteriocins suggest a novel mechanism of iron uptake in Pectobacterium spp. PLoS ONE 2012, 7, e33033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooney, W.M.; Chai, R.; Milner, J.J.; Walker, D. Bacteriocins targeting Gram-negative phytopathogenic bacteria: Plantibiotics of the future. Front. Microbiol. 2020, 11, 575981. [Google Scholar] [CrossRef]
- Behrens, H.M.; Lowe, E.D.; Gault, J.; Housden, N.G.; Kaminska, R.; Weber, T.M.; Catriona, M.A.; Mislin, G.L.A.; Schalk, I.J.; Walker, D. Pyocin S5 import into Pseudomonas aeruginosa reveals a generic mode of bacteriocin transport. mBio 2020, 11, e03230-19. [Google Scholar] [CrossRef] [Green Version]
- Michel-Briand, Y.; Baysse, C. The pyocins of Pseudomonas aeruginosa. Biochimie 2002, 84, 499–510. [Google Scholar] [CrossRef]
- Cascales, E.; Buchanan, S.K.; Duché, D.; Kleanthous, C.; Lloubès, R.; Postle, K.; Riley, M.; Slatin, S.; Cavarad, D. Colicin biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [Green Version]
- Walker, D.; Mosbahi, K.; Vankemmelbeke, M.; James, R.; Kleanthous, C. The role of electrostatics in colicin nuclease domain translocation into bacterial cells. J. Biol. Chem. 2007, 282, 31389–31397. [Google Scholar] [CrossRef] [Green Version]
- Jakes, K.S.; Zinder, N.D. Highly purified colicin E3 contains immunity protein. Proc. Natl. Acad. Sci. USA 1974, 71, 3380–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakes, K.; Zinder, N.D.; Boon, T. Purification and properties of colicin E3 immunity protein. J. Biol. Chem. 1974, 249, 438–444. [Google Scholar] [CrossRef]
- Vankemmelbeke, M.; Zhang, Y.; Moore, G.R.; Kleanthous, C.; Penfold, C.N.; James, R. Energy-dependent immunity protein release during tol-dependent nuclease colicin translocation. J. Biol Chem. 2009, 284, 18932–18941. [Google Scholar] [CrossRef] [Green Version]
- Addgene: Handling Plasmids from Addgene—Purifying Plasmid DNA. Addgene.org. Available online: https://www.addgene.org/protocols/purify-plasmid-dna/ (accessed on 5 March 2020).
- Sambrook, J.; Russel, D.W. Molecular Cloning: A laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Liu, Y.G.; Whittier, R.F. Thermal asymmetric interlaced PCR: Automable amplification and sequencing of insert and fragments from P1 and YAC clones for chromosome walking. Genomics 1995, 25, 674–681. [Google Scholar] [CrossRef]
- Chiu, J.; March, P.E.; Lee, R.; Tillett, D. Site-directed, ligase-independent mutagenesis (SLIM): A single-tube methodology approaching 100% efficiency in 4 h. Nucleic Acids Res. 2004, 32, e174. [Google Scholar] [CrossRef]
- Fyfe, J.A.; Harris, G.; Govan, J.R. Revised pyocin typing method for Pseudomonas aeruginosa. J. Clin. Microbiol. 1984, 20, 47–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, S.; Ross, R.P.; Hill, C. Bacteriolytic activity caused by the presence of a novel lactococcal plasmid encoding lactococcins A, B, and M. Appl. Environ. Microbiol. 1995, 61, 2995–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Kelly, S.; Price, N. The use of circular dichroism in the investigation of protein structure and function. Curr. Protein Pept. Sci. 2000, 1, 349–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to study proteins by circular dichroism. Biochim. Biophys. Acta Proteins Proteom. 2005, 1751, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Ogawa, T.; Uozumi, T.; Watanabe, K.; Masaki, H. A cytotoxic ribonuclease which specifically cleaves four isoaccepting arginine tRNAs at their anticodon loops. Proc. Natl. Acad. Sci. USA 2000, 97, 8278–8283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, H.; Ogawa, T. The modes of action of colicins E5 and D, and related cytotoxic tRNases. Biochimie 2002, 84, 433–438. [Google Scholar] [CrossRef]
- Graille, M.; Mora, L.; Buckingham, R.H.; van Tilbeurgh, H.; de Zamaroczy, M. Structural inhibition of the colicin D tRNase by the tRNA-mimicking immunity protein. EMBO J. 2004, 23, 1474–1482. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Relevant Characteristics | Source of Reference |
|---|---|---|
| Pectobacterium carotovorum subsp. carotovorum | ||
| F-rif-18 | Pcc, Rifr, wild-type | Laboratory stock |
| SP33 | Wild type, indicator | Laboratory stock |
| Escherichia coli | ||
| DH5α | supE44ΔlacU169(ψ80 lacZΔM15)hsdR17 recA1 endA1 gyrA96 thi-1 relA1 | Laboratory stock |
| BL21 (DE3) | hsdS gal(λcIts857 ind1 Sam7 nin5 lacUV5-T7 gene 1) | Laboratory stock |
| Plasmid | ||
| pGEM T-Easy | Ampr; lacZ cloning vector | Promega |
| pET30b | Kanr; expression vector with the C-terminal His-tag | Novagen |
| pET32a | Ampr; expression vector with the N-terminal His-tag | Novagen |
| pEN2K | caroS2K subcloned into pET32a | This study |
| pEX2K | Derived from pEN2K; deleted series of Tag-element in front of expressed caroS2K | This study |
| pEC2I | Caros2I subcloned intro pET30b | This study |
| pEX2I | Derived from pEC2I; deleted series of Tag-element in front of expressed caroS2I | This study |
| pEH2K | Derived from pEC2I; adding (His)6-Tag adjacent to caroS2I | This study |
| pES2TKD400 | N-terminally truncated Carocin S2 at Gln400 | This study |
| pES2TKD600 | N-terminally truncated Carocin S2 at Arg600 | This study |
| pES2TKD677 | N-terminally truncated Carocin S2 at deleted Gly677 | This study |
| pES2TKD691 | N-terminally truncated Carocin S2 at deleted Lys691 | This study |
| pES2TKD692 | N-terminally truncated Carocin S2 at deleted Lys692 | This study |
| pES2TKD693 | N-terminally truncated Carocin S2 at deleted Lys693 | This study |
| Primers | Sequence (5′-3′) |
|---|---|
| CaroS2I_C-taq_for | CATATGATGAGTAATAAACT |
| CaroS2I_C-taq_rev | CTCGAGAAGAAGTTTGAA |
| X2I_forT | GGAAAAATTCAAACTTCTTTGAGATCCGGCTGCT |
| X2I_forS | TGAGATCCGGCTGCT |
| X2I_revT | AAGAAGTTTGAATTTTTCCAACGTGGCTTTTATTTC |
| X2I_revS | AACGTGGCTTTTATTTC |
| CarocinS2K_for2 | CGGTCAGGATCCATGATTAAGTAC |
| CarocinS2I_rev2 | GCGCCAAAGCTTCAAGAGATATCA |
| 5IHT32a2KI_forT | GAAGGAGATATACATATGATTAAGTACCGTTTATA |
| 5IHTGT2KI_forS | ATGATTAAGTACCGTTTATA |
| 5IHT32a3KI_revT | ATGTATATCTCCTTCTTAAAGTTAAACAAAATTATTTC |
| 5IHT32a4KI_revS | TTAAAGTTAAACAAAATTATTTC |
| C2KIH_forT | CACCACCACCACCACCACTGATATCTCAAGCTTGCG |
| C2KIH_forS | TGATATCTCAAGCTTGCG |
| C2KIH_revT | GTGGTGGTGGTGGTGGTGAAGAAGTTTGAATTTTTCC |
| C2KIH_revS | AAGAAGTTTGAATTTTTCC |
| S2TKD667_revT | CATATGTATATCTCCTTCTTAAAGTTAAACAAAATTATTTC |
| 5IHT32a4KI_revS | TTAAAGTTAAACAAAATTATTTC |
| S2TKD400_forT | GAAGGAGATATACATATGCAGGCTTATTTCAGAGC |
| S2TKD400_forS | CAGGCTTATTTCAGAGC |
| S2TKD450_forT | GAAGGAGATATACATATGATAAAACGCAACAGGGT |
| S2TKD450_forS | ATAAAACGCAACAGGGT |
| S2TKD500_forT | GAAGGAGATATACATATGAAGTCACAAGGGATGATTGG |
| S2TKD500_forS | AAGTCACAAGGGATGATTGG |
| S2TKD600_forT | GAAGGAGATATACATATGCGCCTTGTACTGGAAAACC |
| S2TKD600_forS | CGCCTTGTACTGGAAAACC |
| S2TKD677_forT | GAAGGAGATATACATATGGATCCCTTGGATTCAGATCGG |
| S2TKD677_forS | GATCCCTTGGATTCAGATCGG |
| S2TKD691_forT | GAAGGAGATATACATATGAAAAAGTATCTTAAACATGCC |
| S2TKD691_forT | AAAAAGTATCTTAAACATGCC |
| S2TKD692_forT | GAAGGAGATATACATATGAAGTATCTTAAACATGCC |
| S2TKD692_forS | AAGTATCTTAAACATGCC |
| S2TKD693_forT | GAAGGAGATATACATATGTATCTTAAACATGCCAAAG |
| S2TKD693_forS | TATCTTAAACATGCCAAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, P.-C.; Lagitnay, R.B.J.S.; Derilo, R.C.; Wu, J.-L.; Chuang, Y.; Lin, J.-D.; Chuang, D.-Y. Unraveling the Uncharacterized Domain of Carocin S2: A Ribonuclease Pectobacterium carotovorum subsp. carotovorum Bacteriocin. Microorganisms 2022, 10, 359. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020359
Chung P-C, Lagitnay RBJS, Derilo RC, Wu J-L, Chuang Y, Lin J-D, Chuang D-Y. Unraveling the Uncharacterized Domain of Carocin S2: A Ribonuclease Pectobacterium carotovorum subsp. carotovorum Bacteriocin. Microorganisms. 2022; 10(2):359. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020359
Chicago/Turabian StyleChung, Ping-Chen, Ruchi Briam James S. Lagitnay, Reymund C. Derilo, Jian-Li Wu, Yutin Chuang, Jia-De Lin, and Duen-Yau Chuang. 2022. "Unraveling the Uncharacterized Domain of Carocin S2: A Ribonuclease Pectobacterium carotovorum subsp. carotovorum Bacteriocin" Microorganisms 10, no. 2: 359. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10020359