
Dynamic Microstructure Assembly Driven by Lysinibacillus sp. LF-N1 and Penicillium oxalicum DH-1 Inoculants Corresponds to Composting Performance


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Inoculant Preparation
2.2. Composting and Sampling
2.3. Physicochemical Analysis
2.4. DNA Extraction
2.5. Bioinformatics Analyses
3. Results
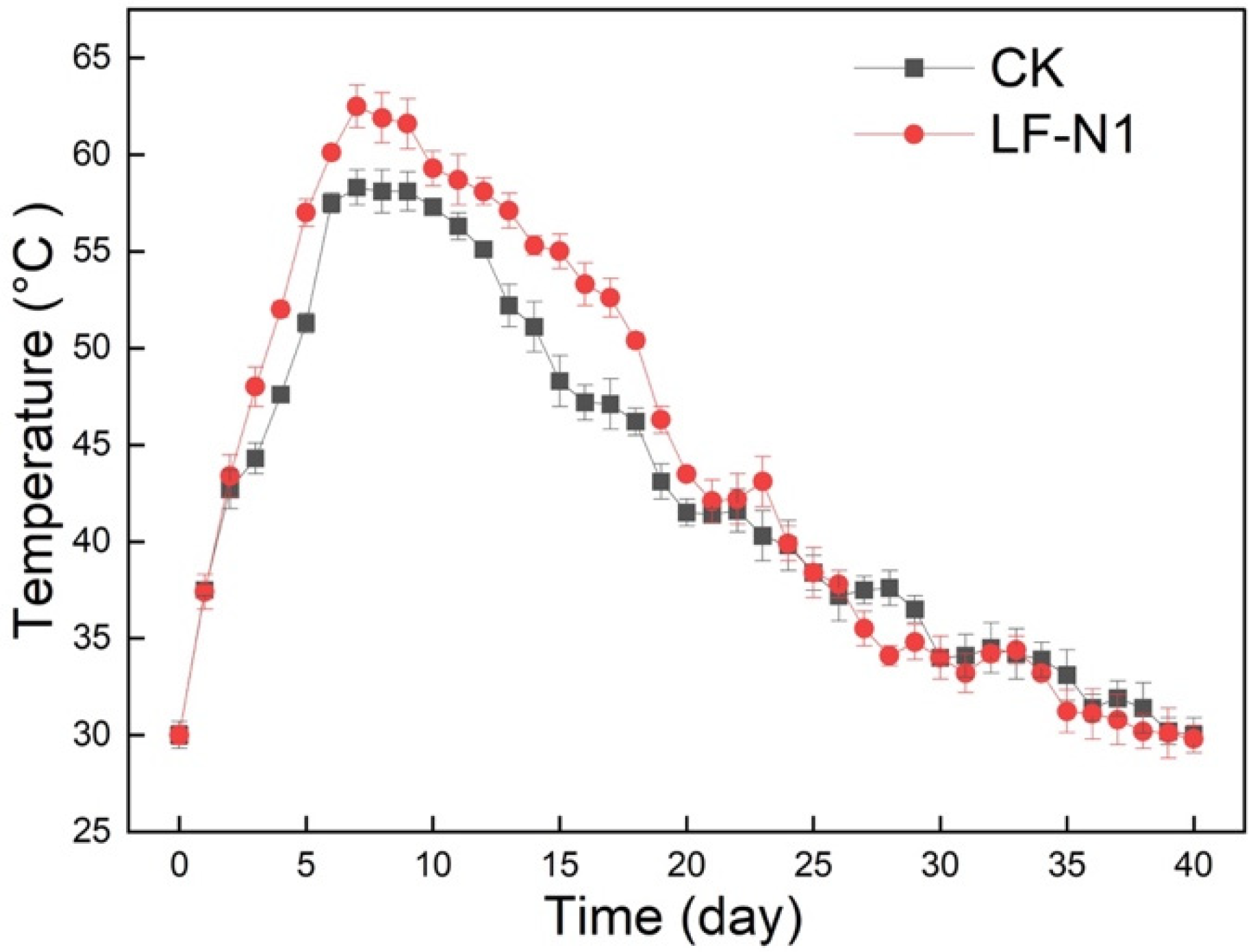
3.1. Variations in Physicochemical Properties
3.2. Degradation of Organic Matter and Lignocellulose
3.3. Similarity and Diversity of the Microbial Community
3.4. Microbial Dynamics Analysis
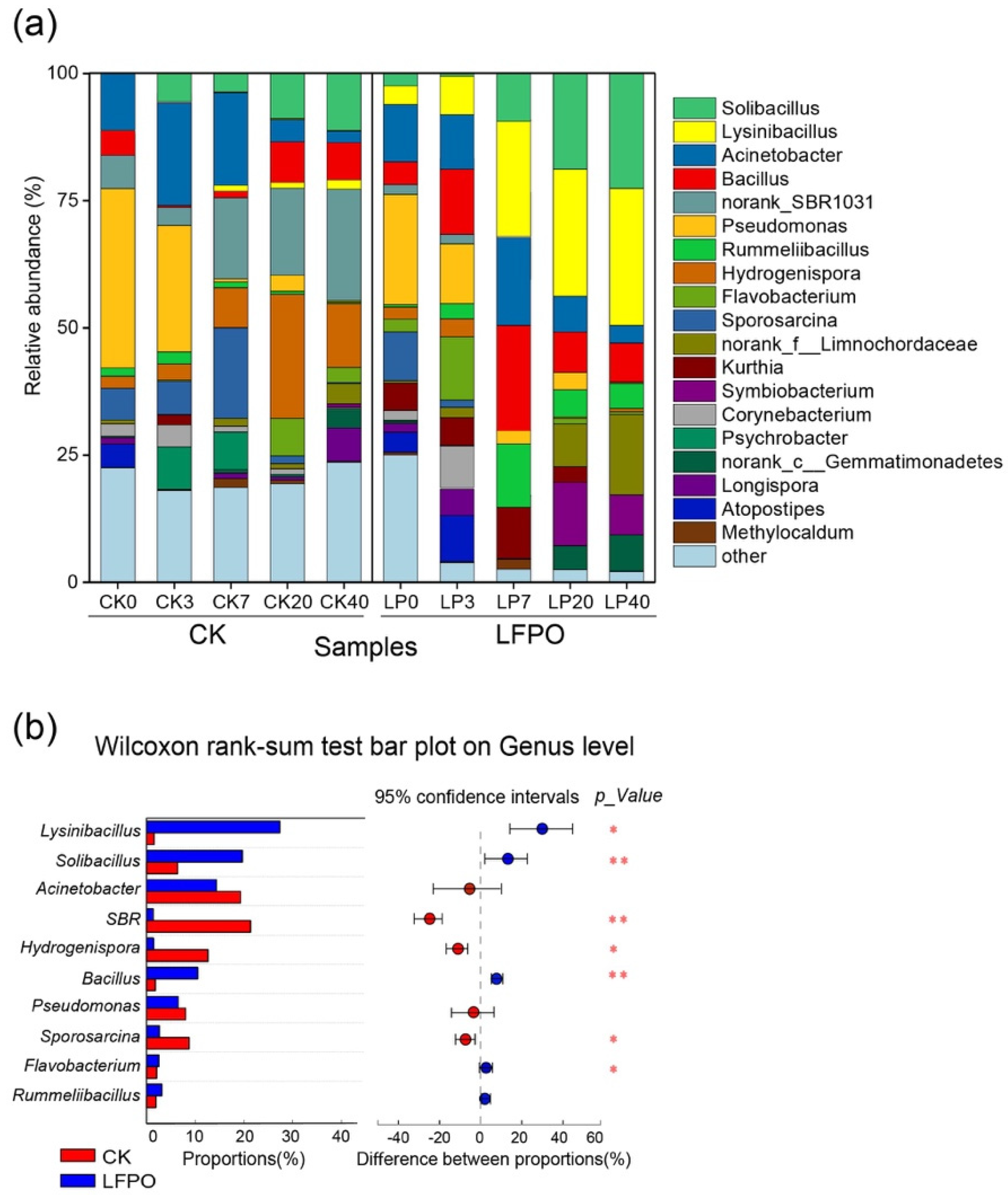
3.4.1. Bacterial Community Succession
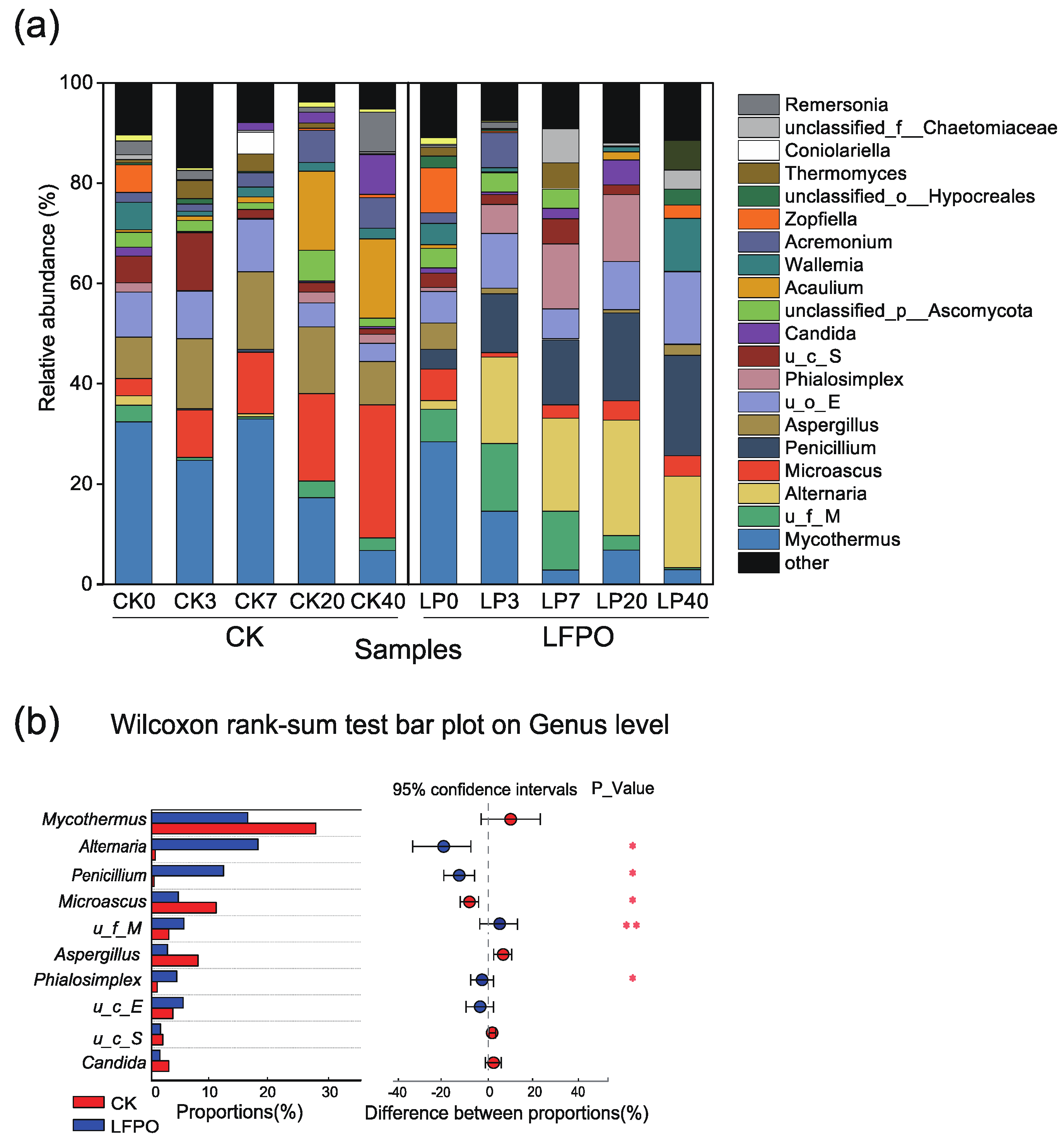
3.4.2. Fungal Community Succession
3.5. Effects on the Microbial Dynamicd and Function Prediction
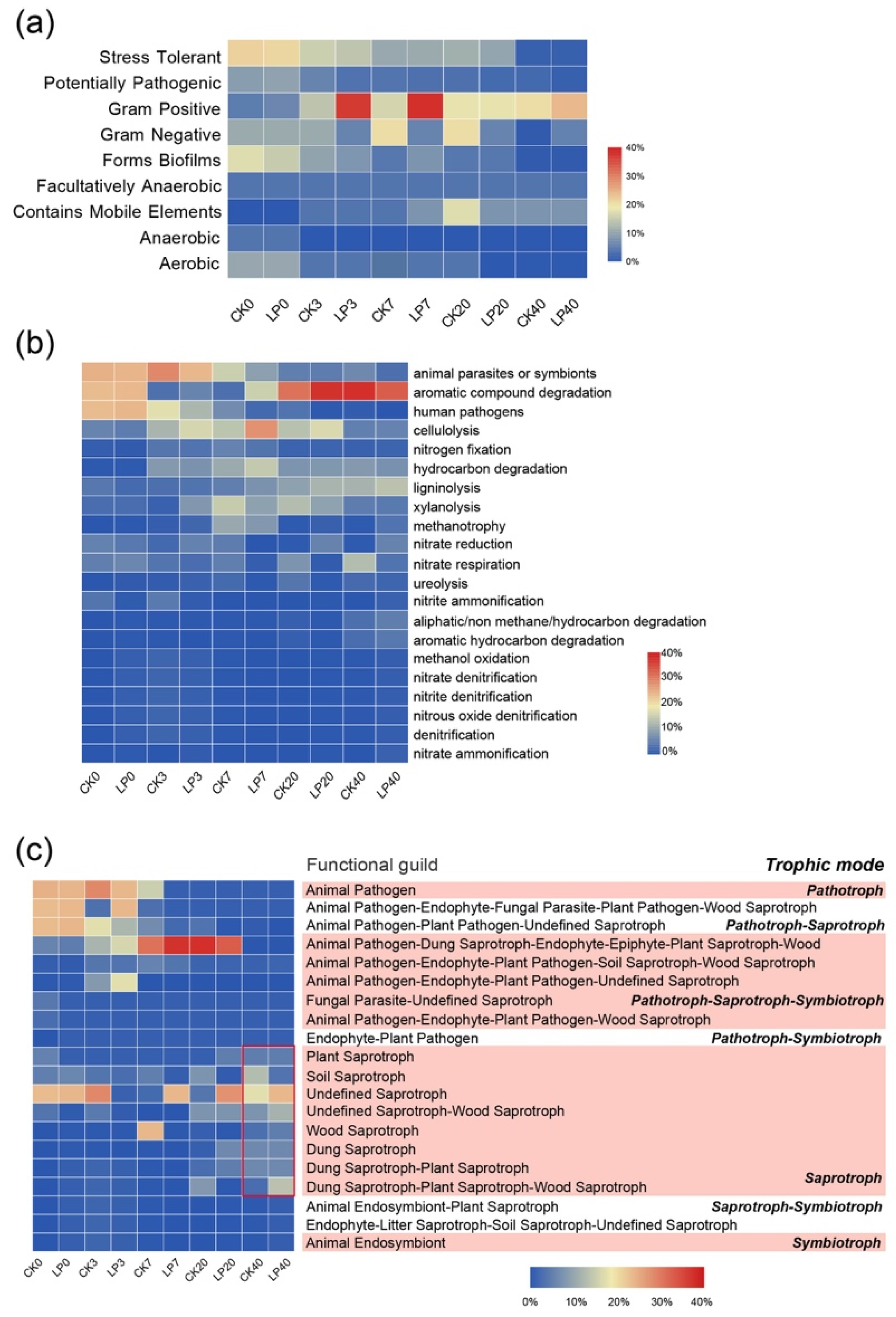
3.5.1. Effects on the Phenotype of Bacterial Communities
3.5.2. Fungal Function Prediction
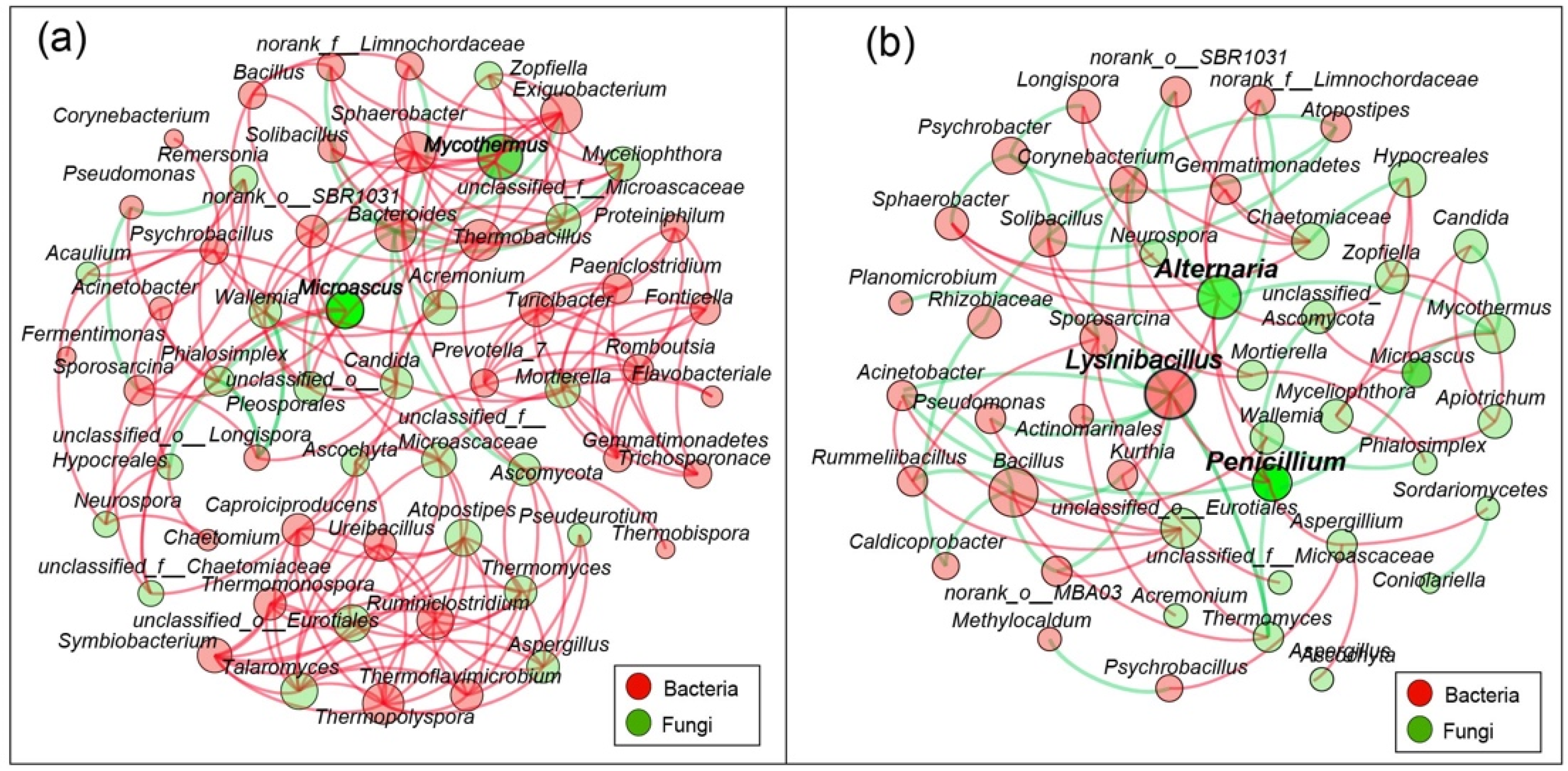
3.6. Co-Networks in Microbial Communities
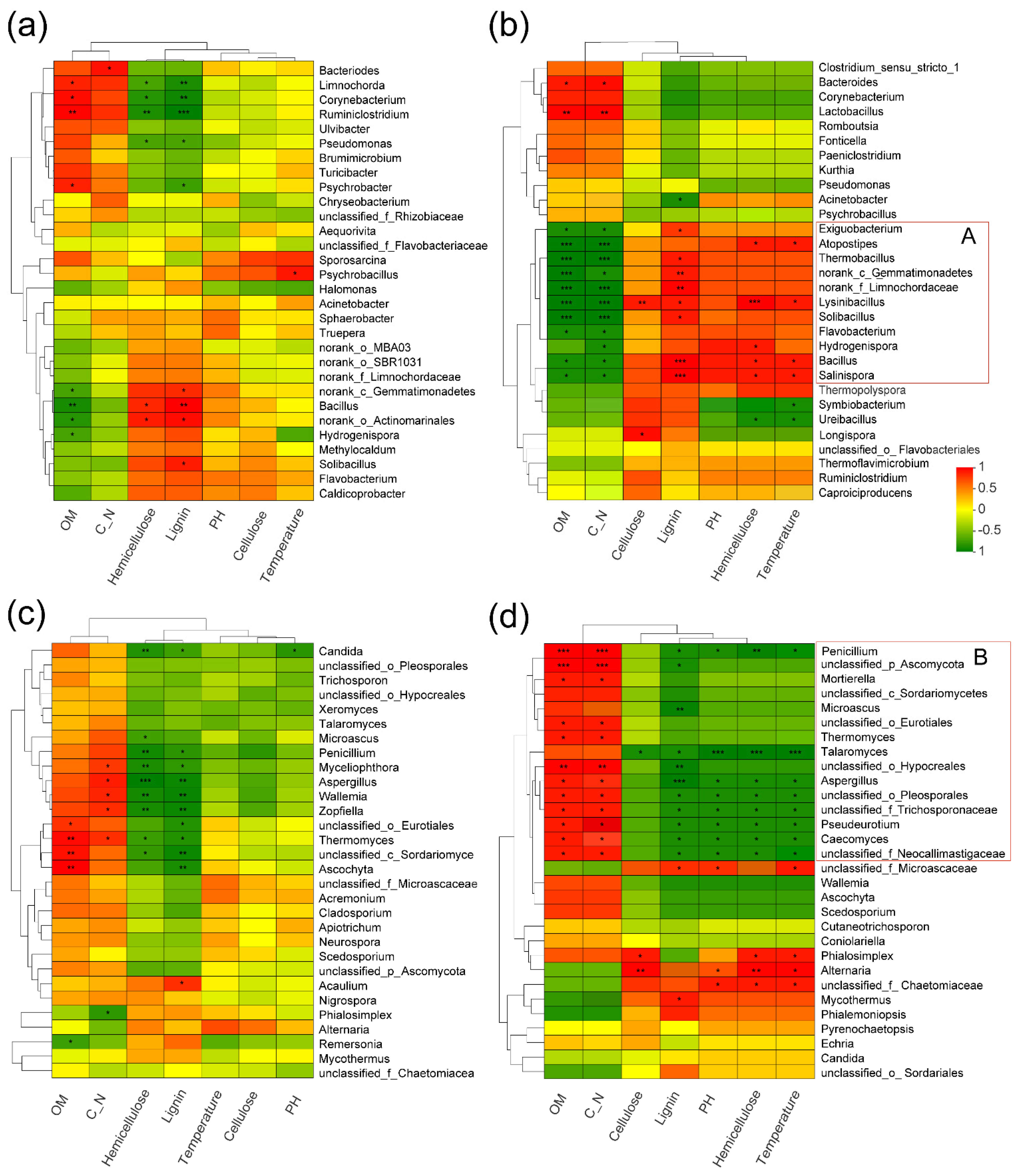
3.7. Relationships of the Microbial Structure with Physicochemical Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, S.; Fang, C.; Sun, X.; Han, L.; He, X.; Huang, G. Bacterial community succession during pig manure and wheat straw aerobic composting covered with a semi-permeable membrane under slight positive pressure. Bioresour. Technol. 2018, 259, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Qian, T.-T.; Wu, P.; Qin, Q.-Y.; Huang, Y.-N.; Wang, Y.-J.; Zhou, D.-M. Screening of wheat straw biochars for the remediation of soils polluted with Zn (II) and Cd (II). J. Hazard. Mater. 2018, 362, 311–317. [Google Scholar] [CrossRef]
- Xie, X.-Y.; Zhao, Y.; Sun, Q.-H.; Wang, X.-Q.; Cui, H.-Y.; Zhang, X.; Li, Y.-J.; Wei, Z.-M. A novel method for contributing to composting start-up at low temperature by inoculating cold-adapted microbial consortium. Bioresour. Technol. 2017, 238, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Wang, X.; Gu, J.; Zhang, S.; Yin, Y.; Li, Y.; Qian, X.; Sun, W. Effects of different swine manure to wheat straw ratios on antibiotic resistance genes and the mi-crobial community structure during anaerobic digestion. Bioresour. Technol. 2017, 231, 1–8. [Google Scholar] [CrossRef]
- Gou, C.; Wang, Y.; Zhang, X.; Lou, Y.; Gao, Y. Inoculation with a psychrotrophic-thermophilic complex microbial agent accelerates onset and pro-motes maturity of dairy manure-rice straw composting under cold climate conditions. Bioresour. Technol. 2017, 243, 339–346. [Google Scholar] [CrossRef]
- Tran, Q.N.M.; Mimoto, H.; Nakasaki, K. Inoculation of lactic acid bacterium accelerates organic matter deg-radation during composting. Int. Biodeterior. Biodegrad. 2015, 104, 377–383. [Google Scholar] [CrossRef]
- Xu, J.; Jiang, Z.; Li, M.; Li, Q. A compost-derived thermophilic microbial consortium enhances the humification process and alters the microbial diversity during composting. J. Environ. Manag. 2019, 243, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.P.; Chu, S.; Wang, B.; Zhang, D.; Zhi, Y.; Yang, X.; Zhou, P. Dynamic bacterial assembly driven by Streptomyces griseorubens JSD-1 inoculants correspond to composting performance in swine manure and rice straw co-composting. Bioresour. Technol. 2020, 313, 123692. [Google Scholar] [CrossRef]
- Jurado, M.; Estrella, F.S.; Lopez, M.J.; García, M.D.C.V.; López-González, J.A.; Moreno, J. Enhanced turnover of organic matter fractions by microbial stimulation during lignocellulosic waste composting. Bioresour. Technol. 2015, 186, 15–24. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Liu, Y.; Li, H.; Liu, H. Effect of microbiological inoculants DN-1 on lignocellulose degradation during co-composting of cattle manure with rice straw monitored by FTIR and SEM. Environ. Prog. Sustain. Energy 2015, 35, 345–351. [Google Scholar] [CrossRef]
- Zeng, G.; Yu, M.; Chen, Y.; Huang, D.; Zhang, J.; Huang, H.; Jiang, R.; Yu, Z. Effects of inoculation with Phanerochaete chrysosporium at various time points on enzyme activi-ties during agricultural waste composting. Bioresour. Technol. 2010, 101, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Wang, X.; Zhen, L.; Gu, J.; Zhang, K.; Wang, Q.; Ma, J.; Peng, H.; Lei, L.; Zhao, W. Effects of inoculating with lignocellulose-degrading consortium on cellulose-degrading genes and fungal community during co-composting of spent mushroom substrate with swine manure. Bioresour. Technol. 2019, 291, 121876. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, L.; Qin, Y.; Zou, G.; Li, Z.; Yan, X.; Wei, X.; Chen, M.; Chen, L.; Zheng, K.; et al. Long-term strain improvements accumulate mutations in regulatory elements responsible for hy-per-production of cellulolytic enzymes. Sci. Rep. 2013, 3, 1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinitsyn, A.V.G.A.P. Cellulases from Penicillium species for producing fuels from biomass. Biofuels 2012, 3, 463–477. [Google Scholar]
- Wang, H.-Y.; Fan, B.-Q.; Hu, Q.-X.; Yin, Z.-W. Effect of inoculation with Penicillium expansum on the microbial community and maturity of compost. Bioresour. Technol. 2011, 102, 11189–11193. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Bingcheng, G.; Weihong, P.; Dinghong, J.; Liyuan, X.; Zhongqian, H.; Jian, G. Screening and identification of a cellulose-decomposing strain Lysinibacillus fusiformis. Chin. J. Environ. Eng. 2013, 7, 12. [Google Scholar]
- Persinoti, G.F.; Paixão, D.A.A.; Bugg, T.D.H.; Squina, F.M. Genome Sequence of Lysinibacillus sphaericus, a Lignin-Degrading Bacterium Isolated from Municipal Solid Waste Soil. Genome Announc. 2018, 6, e00353-18. [Google Scholar] [CrossRef] [Green Version]
- Chantarasiri, A.; Boontanom, P.; Nuiplot, N.O. Isolation and characterization of Lysinibacillus sphaericus BR2308 from coastal wetland in Thailand for the biodegradation of lignin. Aquac. Aquar. Conserv. Legis. 2017, 10, 200–209. [Google Scholar]
- Zhao, Y.; Lu, Q.; Wei, Y.; Cui, H.; Zhang, X.; Wang, X.; Shan, S.; Wei, Z. Effect of actinobacteria agent inoculation methods on cellulose degradation during composting based on redundancy analysis. Bioresour. Technol. 2016, 219, 196–203. [Google Scholar] [CrossRef]
- López-González, J.A.; Estrella, F.S.; García, M.D.C.V.; Lopez, M.J.; Jurado, M.; Moreno, J. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity. Bioresour. Technol. 2015, 175, 406–416. [Google Scholar] [CrossRef]
- Duan, M.; Zhang, Y.; Zhou, B.; Qin, Z.; Wu, J.; Wang, Q.; Yin, Y. Effects of Bacillus subtilis on carbon components and microbial functional metabolism during cow manure–straw composting. Bioresour. Technol. 2020, 303, 122868. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Non-starch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of mixingNeolamarckia cadambaleaves on fermentation quality, microbial community of high moisture alfalfa and stylo silage. Microb. Biotechnol. 2019, 12, 869–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masebinu, S.O.; Akinlabi, E.T.; Muzenda, E.; Aboyade, A.O.; Mbohwa, C. Experimental and feasibility assessment of biogas production by anaerobic digestion of fruit and vegetable waste from Joburg Market. Waste Manag. 2018, 75, 236–250. [Google Scholar] [CrossRef]
- Reimer, L.C.; Vetcininova, A.; Carbasse, J.S.; Söhngen, C.; Gleim, D.; Ebeling, C.; Overmann, J. BacDivein 2019: Bacterial phenotypic data for High-throughput biodiversity analysis. Nucleic Acids Res. 2018, 47, D631–D636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Y.; Shaaban, M.; Zhu, D.; Hu, C.; Chen, Z.; Wang, Y. Evaluation of microbial inoculants pretreatment in straw and manure co-composting process enhancement. J. Clean. Prod. 2019, 239, 118078. [Google Scholar] [CrossRef]
- Zhang, D.; Luo, W.; Wang, G.; Li, G. Performance of co-composting sewage sludge and organic fraction of municipal solid waste at dif-ferent proportions. Bioresour. Technol. 2018, 250, 853–859. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Y.; Liu, J.; Yang, X.; Ren, Y.; Miao, H.; Li, Y. Exploring the mechanisms of organic matter degradation and methane emission during sewage sludge composting with added vesuvianite: Insights into the prediction of microbial metabolic function and enzy-matic activity. Bioresour. Technol. 2019, 286, 121397. [Google Scholar] [CrossRef]
- Du, G.; Feng, W.; Cai, H.; Ma, Z.; Liu, X.; Yuan, C.; Shi, J.; Zhang, B. Exogenous enzyme amendment accelerates maturity and changes microflora succession in horse and wildlife animal manure co-composting. Environ. Sci. Pollut. Res. 2021, 28, 21610–21620. [Google Scholar] [CrossRef]
- Glasser, W.G. About Making Lignin Great Again—Some Lessons From the Past. Front. Chem. 2019, 7, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Wu, D.; Wei, D.; Zhao, Y.; Wu, J.; Xie, X.; Zhang, R.; Wei, Z. Improved lignocellulose-degrading performance during straw composting from diverse sources with actinomycetes inoculation by regulating the key enzyme activities. Bioresour. Technol. 2018, 271, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Dhaouadi, Y.; Stoodley, P.; Ren, D. Sensing the unreachable: Challenges and opportunities in biofilm detection. Curr. Opin. Biotechnol. 2019, 64, 79–84. [Google Scholar] [CrossRef]
- Wall, G.; Montelongo-Jauregui, D.; Bonifacio, B.V.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Duan, H.; Ji, M.; Xie, Y.; Shi, J.; Liu, L.; Zhang, B.; Sun, J. Exploring the Microbial Dynamics of Organic Matter Degradation and Humification during Co-Composting of Cow Manure and Bedding Material Waste. Sustainability 2021, 13, 13035. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | Cow Manure | Wheat Straw |
|---|---|---|
| Moisture (%) | 61.6 ± 1.9 | 8.7 ± 1.7 |
| pH | 7.92 ± 0.07 | 6.63 ± 0.05 |
| OM (Organic matter, g·kg−1)) | 785.4 ± 2.3 | 653 ± 2.3 |
| TOC (Total organic carbon, g·kg−1) | 312 ± 1.5 | 443 ± 2.3 |
| TKN (Total Kjeldahl nitrogen, g·kg−1) | 14.7 ± 0.02 | 7.9 ± 0.01 |
| C/N (Carbon: nitrogen ratio) | 29.1 ± 0.3 | 24.2 ± 0.5 |
| Cellulose (%) | 33.9 ± 1.3 | 39.4 ± 1.8 |
| Hemicellulose (%) | 26.5 ± 1.6 | 28.4 ± 1.2 |
| Lignin (%) | 13.5 ± 0.2 | 4.6 ± 0.03 |
| Items | Time (Day) | SEM | Significance | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 7 | 20 | 40 | TR | TI | TR × TI | ||
| pH | |||||||||
| CK | 7.67 ± 0.01 Ea | 8.36 ± 0.01 Aa | 8.21 ± 0.01 Ba | 7.89 ± 0.01 Db | 8.04 ± 0.06 Ca | 0.004 | * | *** | *** |
| LFPO | 7.68 ± 0.02 Ea | 8.31 ± 0.03 Aa | 8.18 ± 0.01 Ba | 8.03 ± 0.01 Ca | 7.87 ± 0.01 Db | ||||
| TOC (g·kg−1) | |||||||||
| CK | 374.6 ± 10.59 Aa | 349.3 ± 4.16 ABa | 336.0 ± 6.25 ABa | 286.1 ± 58.0 Ba | 282.3 ± 10.4 Ba | 3.675 | 0.134 | *** | 0.673 |
| LFPO | 373.1 ± 13.1 Aa | 343.6 ± 9.60 Ba | 321.7 ± 4.04 Ba | 282.7 ± 9.61 Ca | 269.6 ± 5.58 Cb | ||||
| TN (g·kg−1) | |||||||||
| CK | 14.6 ± 0.51 Aa | 13.9 ± 0.21 Aa | 15.4 ± 0.34 Aa | 14.6 ± 2.86 Aa | 16.1 ± 0.29 Aa | 0.182 | 0.331 | * | 0.759 |
| LFPO | 14.5 ± 0.72 BCa | 14.3 ± 0.66 Ca | 16.3 ± 0.44 Aa | 15.6 ± 0.34 ABCa | 16.8 ± 0.49 ABa | ||||
| C/N | |||||||||
| CK | 25.6 ± 0.25 Aa | 24.9 ± 0.11 Aa | 21.7 ± 0.32 Ba | 19.5 ± 0.17 Ca | 18.7 ± 0.36 Da | 0.075 | *** | *** | * |
| LFPO | 25.5 ± 0.41 Aa | 23.9 ± 0.61 Bb | 19.7 ± 0.35 Cb | 18.1 ± 0.26 Db | 16.4 ± 0.78 Eb | ||||
| GI | |||||||||
| CK | 32.3 ± 1.15 Ea | 44.7 ± 0.45 Db | 73.5 ± 0.56 Cb | 101.5 ± 2.76 Bb | 125.4 ± 5.50 Ab | 0.773 | 0.331 | *** | *** |
| LFPO | 33.1 ± 1.05 a | 55.8 ± 0.95 a | 88.2 ± 1.05 a | 136.0 ± 6.45 a | 177.1 ± 9.72 a | ||||
| Items | Time (Day) | SEM | Significance | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 3 | 7 | 20 | 40 | TR | TI | TR × TI | ||
| OM (organic matter, g·kg−1) | |||||||||
| CK | 782.3 ± 17.6 Aa | 736.1 ± 6.0 Ba | 679.2 ± 4.6 Ca | 543.4 ± 10.5 Da | 521.6 ± 3.0 Da | 0.225 | * | *** | *** |
| LFPO | 778.7 ± 17.4 Aa | 665.7 ± 10.7 Bb | 527.3 ± 15.5 Cb | 460.9 ± 16.1 Db | 420.1 ± 10.5 Eb | ||||
| Cellulose (g·kg−1) | |||||||||
| CK | 363.3 ± 15.5 Aa | 353.6 ± 13.2 ABa | 319.6 ± 14.1 Ba | 244.3 ± 10.4 Ca | 193.6 ± 11.0 Da | 2.203 | *** | *** | *** |
| LFPO | 364.0 ± 8.54 Aa | 321.3 ± 14.0 Bb | 235.0 ± 11.5 Cb | 154.3 ± 13.0 Db | 125.0 ± 6.0 Db | ||||
| Hemicellulose (g·kg−1) | |||||||||
| CK | 275.6 ± 17.6 Aa | 232.3 ± 8.50 Ba | 172.5 ± 10.0 Ca | 123.7 ± 10.2 Da | 93.6 ± 10.5 Da | 1.927 | *** | *** | ** |
| LFPO | 278.3 ± 16.5 Aa | 192.3 ± 11.0 Bb | 124.7 ± 3.51 Cb | 71.6 ± 2.31 Db | 53.8 ± 3.02 Db | ||||
| Lignin (g·kg−1) | |||||||||
| CK | 104.6 ± 2.51 Aa | 98.0 ± 6.00 Aa | 89.6 ± 2.08 Ba | 69.0 ± 3.00 Ca | 51.0 ± 8.54 Da | 1.018 | * | *** | 0.787 |
| LFPO | 104.0 ± 2.65 Aa | 89.3 ± 7.64 ABa | 84.3 ± 10.1 Ba | 64.6 ± 2.51 Ca | 44.3 ± 3.21 Da | ||||
| Community | Treatment | Time (Day) | Coverage | Chao 1 | Ace | Shannon | Simpson |
|---|---|---|---|---|---|---|---|
| Richness | Diversity | ||||||
| Bacterial community | CK | 0 | 1.0 | 783.29 ± 3.8 | 892.26 ± 41.8 | 3.20 ± 0.01 | 0.28 ± 0.01 |
| 3 | 1.0 | 620.12 ± 21.3 | 716.45 ± 31.81 | 1.54 ± 0.02 | 0.33 ± 0.01 | ||
| 7 | 1.0 | 404.09 ± 31.1 | 389.50 ± 21.3 | 2.13 ± 0.06 | 0.30 ± 0.01 | ||
| 20 | 1.0 | 476.01 ± 12.7 | 485.06 ± 24.7 | 2.96 ± 0.07 | 0.18 ± 0.01 | ||
| 40 | 1.0 | 597.19 ± 14.3 | 659.16 ± 41.2 | 2.89 ± 0.21 | 0.12 ± 0.08 | ||
| LFPO | 0 | 1.0 | 749.29 ± 4.26 | 878.35 ± 34.6 | 3.30 ± 0.01 | 0.25 ± 0.12 | |
| 3 | 1.0 | 647.17 ± 6.89 | 759.72 ± 12.9 | 1.51 ± 0.07 | 0.43 ± 0.17 | ||
| 7 | 1.0 | 456.72 ± 9.12 | 518.17 ± 14.7 | 1.37 ± 0.05 | 0.41 ± 0.05 | ||
| 20 | 1.0 | 596.57 ± 13.2 | 657.00 ± 31.5 | 2.38 ± 0.21 | 0.25 ± 0.01 | ||
| 40 | 1.0 | 652.08 ± 21.1 | 730.58 ± 23.4 | 2.33 ± 0.01 | 0.26 ± 0.01 | ||
| Fungal community | CK | 0 | 1.0 | 323.25 ± 11.4 | 283.07 ± 25.6 | 4.21 ± 0.22 | 0.03 ± 0.01 |
| 3 | 1.0 | 223.44 ± 21.8 | 188.42 ± 32.5 | 3.19 ± 0.09 | 0.17 ± 0.01 | ||
| 7 | 1.0 | 123.74 ± 22.1 | 172.69 ± 26.5 | 3.88 ± 0.03 | 0.14 ± 0.02 | ||
| 20 | 1.0 | 197.89 ± 11.6 | 192.20 ± 24.3 | 3.01 ± 0.02 | 0.08 ± 0.03 | ||
| 40 | 1.0 | 200.75 ± 27.8 | 219.31 ± 12.5 | 3.13 ± 0.07 | 0.11 ± 0.01 | ||
| LFPO | 0 | 1.0 | 303.05 ± 17.6 | 246.86 ± 21.6 | 4.40 ± 0.04 | 0.11 ± 0.01 | |
| 3 | 1.0 | 293.73 ± 24.3 | 221.06 ± 14.3 | 3.87 ± 0.14 | 0.20 ± 0.07 | ||
| 7 | 1.0 | 173.67 ± 3.87 | 185.17 ± 24.3 | 2.88 ± 0.03 | 0.12 ± 0.01 | ||
| 20 | 1.0 | 227.89 ± 9.12 | 203.87 ± 14.5 | 3.41 ± 0.02 | 0.18 ± 0.01 | ||
| 40 | 1.0 | 226.25 ± 21.8 | 226.99 ± 25.9 | 3.85 ± 0.03 | 0.21 ± 0.01 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, H.; Fu, C.; Du, G.; Xie, S.; Liu, M.; Zhang, B.; Shi, J.; Sun, J. Dynamic Microstructure Assembly Driven by Lysinibacillus sp. LF-N1 and Penicillium oxalicum DH-1 Inoculants Corresponds to Composting Performance. Microorganisms 2022, 10, 709. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040709
Duan H, Fu C, Du G, Xie S, Liu M, Zhang B, Shi J, Sun J. Dynamic Microstructure Assembly Driven by Lysinibacillus sp. LF-N1 and Penicillium oxalicum DH-1 Inoculants Corresponds to Composting Performance. Microorganisms. 2022; 10(4):709. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040709
Chicago/Turabian StyleDuan, Haiyan, Cong Fu, Guilin Du, Shiqiu Xie, Min Liu, Baoguo Zhang, Jiping Shi, and Junsong Sun. 2022. "Dynamic Microstructure Assembly Driven by Lysinibacillus sp. LF-N1 and Penicillium oxalicum DH-1 Inoculants Corresponds to Composting Performance" Microorganisms 10, no. 4: 709. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040709