
Biodegradation of Quinoline by a Newly Isolated Salt-Tolerating Bacterium Rhodococcus gordoniae Strain JH145

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Enrichment and Isolation of Bacteria
2.3. Identification of Strain JH145
2.4. Characteristics of Quinoline Biodegradation by Strain JH145
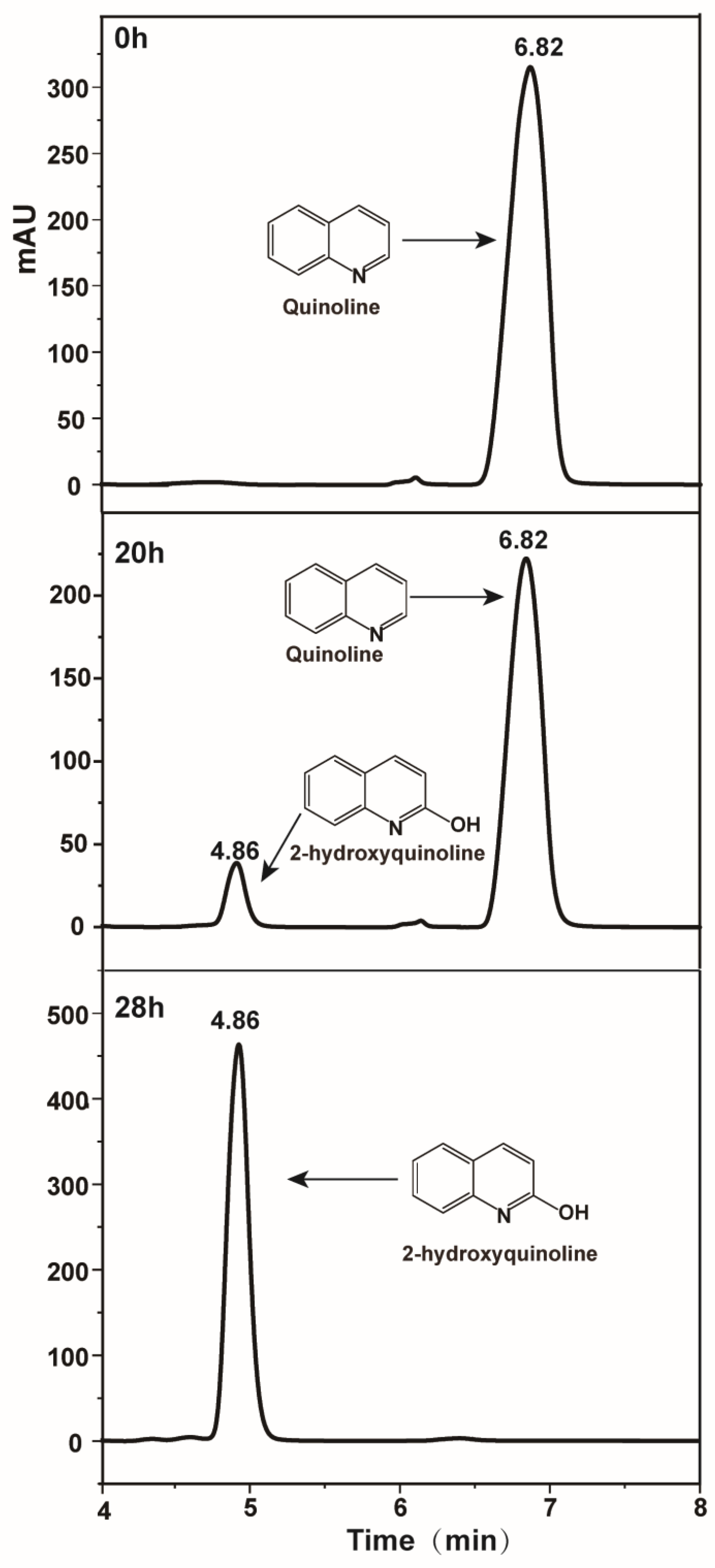
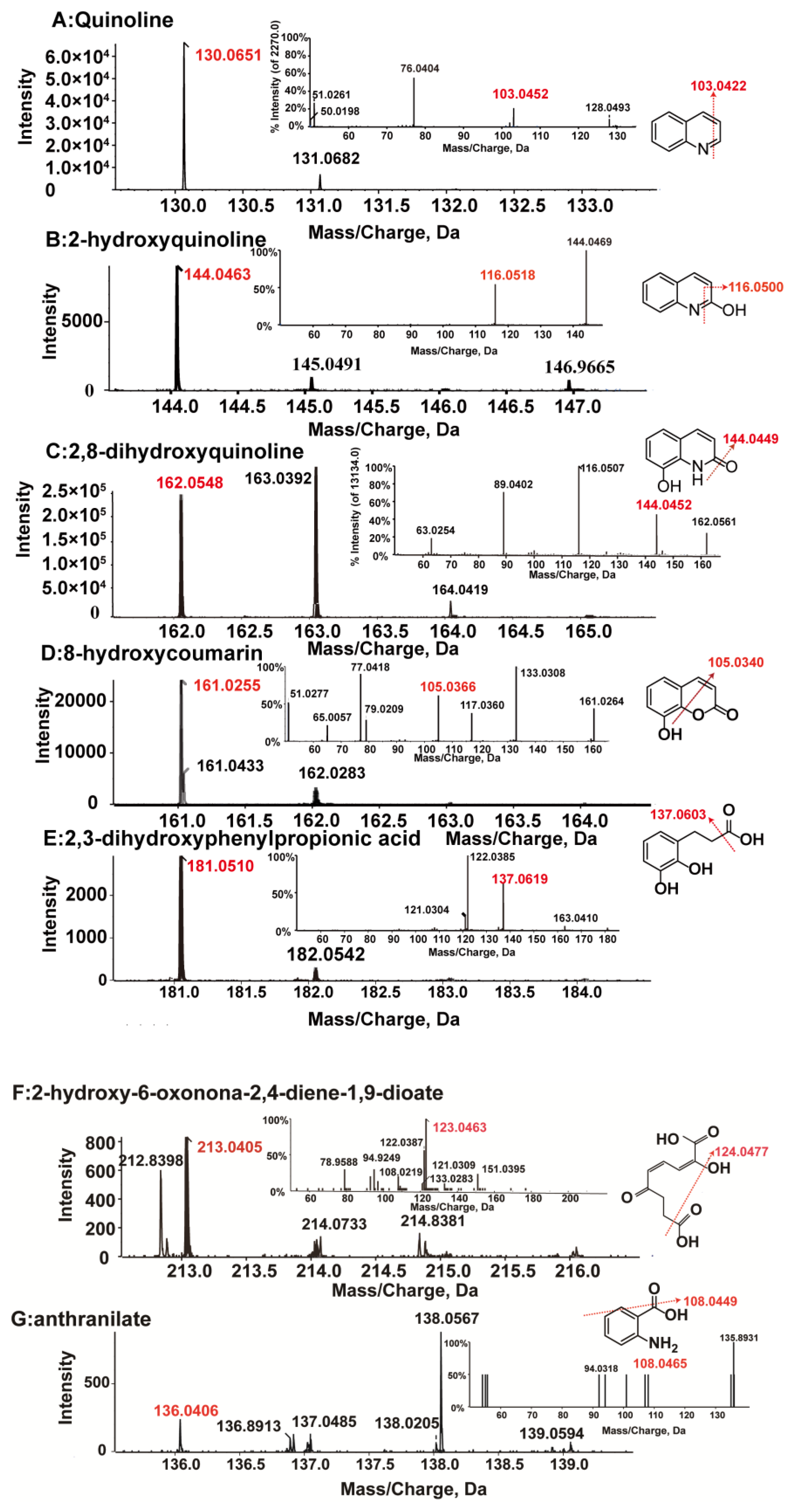
2.5. Detection of Quinoline Metabolites and Analysis Method
3. Results and Discussion
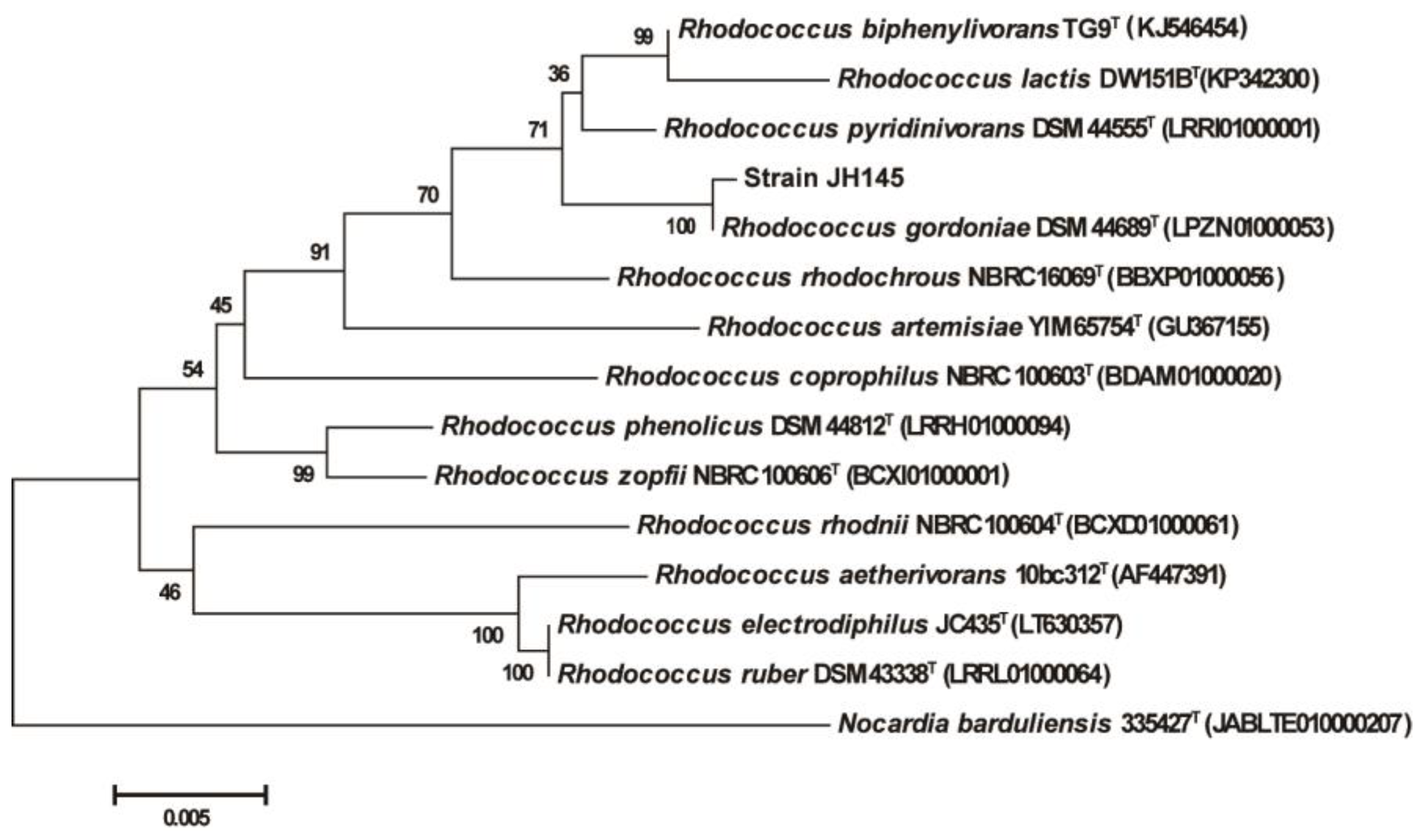
3.1. Identification of Quinoline Degrading Bacteria
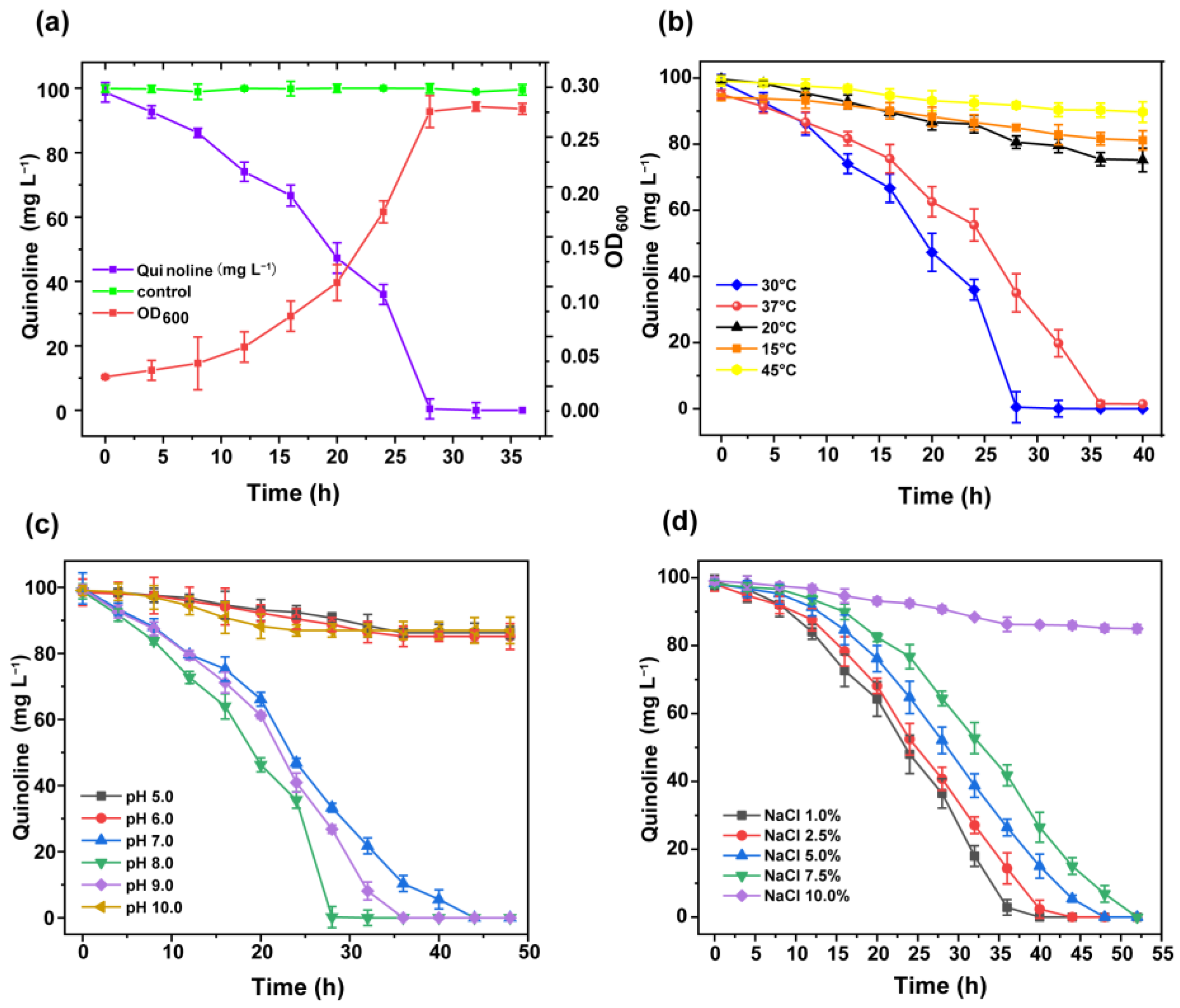
3.2. Growth Status and Degradation Characteristics of Strain JH145
3.2.1. Study on the Growth Profile of Quinoline-Degrading Strain JH145
3.2.2. Study on the Degradability of Quinoline by Temperature
3.2.3. Study on the Degradability of Quinoline by pH
3.2.4. Study on Salt Tolerance of Strain JH145
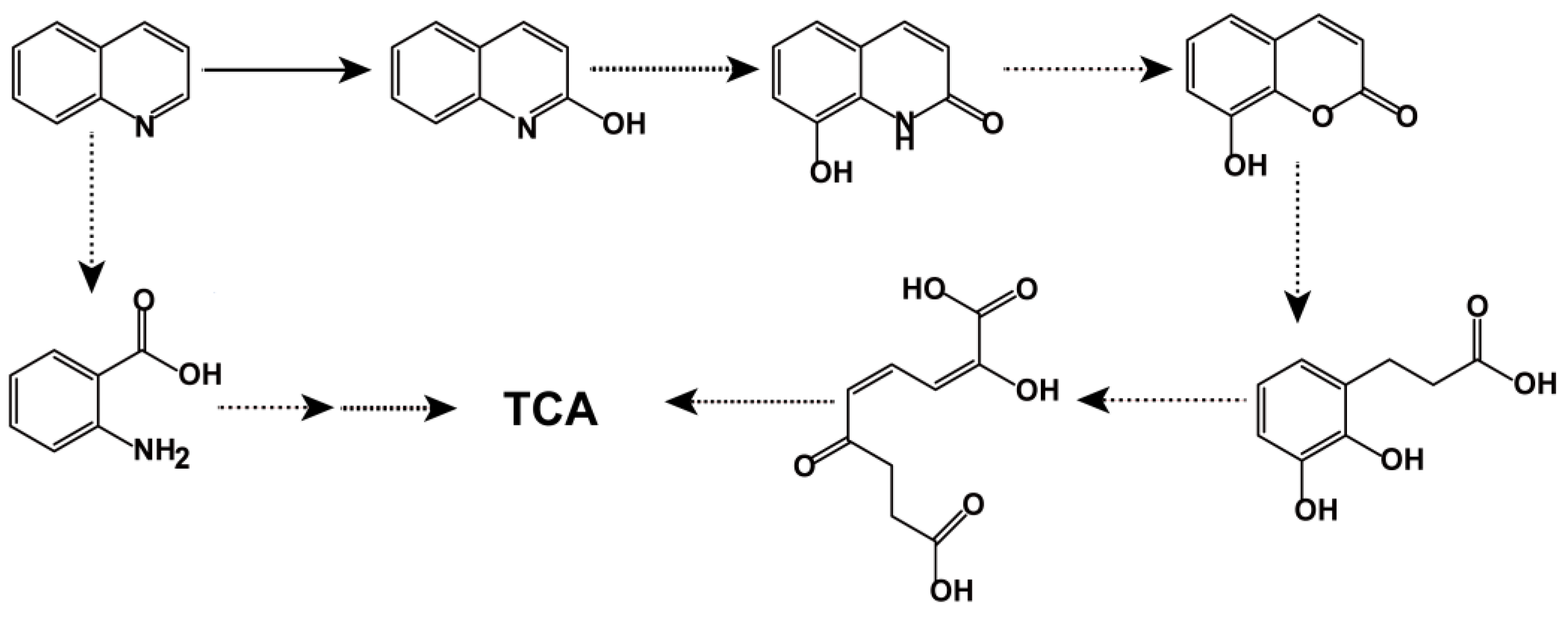
3.3. Identification of Metabolites and Proposal of Quinoline Aerobic Catabolism Pathway
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Felczak, A.; Zawadzka, K.; Lisowska, K. Efficient biodegradation of quinolone-Factors determining the process. Int. Biodeter. Biodegr. 2014, 96, 127–134. [Google Scholar] [CrossRef]
- Shi, J.; Han, H.; Xu, C. A novel enhanced anaerobic biodegradation method using biochar and Fe (OH)3@biochar for the removal of nitrogen heterocyclic compounds from coal gasification wastewater. Sci. Total Environ. 2019, 697, 134052. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liu, R.; Liu, C.; Chen, L. Bioaugmentation with isolated strains for the removal of toxic and refractory organics from coking wastewater in a membrane bioreactor. Biodegradation 2015, 26, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Chen, Q.; Parales, R.E.; Lu, Z.; Hong, Q.; He, J.; Qiu, J.; Jiang, J. Bacterial catabolism of nicotine: Catabolic strains, pathways and modules. Environ. Res. 2020, 183, 109258. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Meng, N.; Li, Y.; Wang, J. Occurrence, impacts, and microbial transformation of 3-methylindole (skatole): A critical review. J. Hazard. Mater. 2021, 416, 126181. [Google Scholar] [CrossRef]
- Shaikhulova, S.; Fakhrullina, G.; Nigamatzyanova, L.; Akhatova, F.; Fakhrullin, R. Worms eat oil: Alcanivorax borkumensis hydrocarbonoclastic bacteria colonise Caenorhabditis elegans nematodes intestines as a first step towards oil spills zooremediation. Sci. Total Environ. 2021, 761, 143209. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, S.; Li, B. Removal of Toxic Organic Pollutants from Coke Plant Wastewater by UV-Fenton. J. Residuals Sci. Technol. 2016, 13, S143–S148. [Google Scholar] [CrossRef] [Green Version]
- Rameshraja, D.; Srivastava, V.C.; Kushwaha, J.P.; Mall, I.D. Quinoline adsorption onto granular activated carbon and bagasse fly ash. Chem. Eng. J. 2012, 181–182, 343–351. [Google Scholar] [CrossRef]
- Zhu, H.; Ma, W.; Han, H.; Xu, C.; Han, Y.; Ma, W. Degradation characteristics of two typical N-heterocycles in ozone process: Efficacy, kinetics, pathways, toxicity and its application to real biologically pretreated coal gasification wastewater. Chemosphere 2018, 209, 319–327. [Google Scholar] [CrossRef]
- Singh, L.; Rekha, P.; Chand, S. Comparative evaluation of synthesis routes of Cu/zeolite Y catalysts for catalytic wet peroxide oxidation of quinoline in fixed-bed reactor. J. Environ. Manag. 2018, 215, 1–12. [Google Scholar] [CrossRef]
- Bai, Q.; Yang, L.; Li, R.; Chen, B.; Zhang, L.; Zhang, Y.; Rittmann, B.E. Accelerating Quinoline Biodegradation and Oxidation with Endogenous Electron Donors. Environ. Sci. Technol. 2015, 49, 11536–11542. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Yu, S.; Wang, J. Degradation of pyridine and quinoline in aqueous solution by gamma radiation. Radiat. Phys. Chem. 2018, 144, 322–328. [Google Scholar] [CrossRef]
- Yi, R.; Yi, C.; Du, D.; Zhang, Q.; Yu, H.; Yang, L. Research on quinoline degradation in drinking water by a large volume strong ionization dielectric barrier discharge reaction system. Plasma Sci. Technol. 2021, 23, 085505. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Y. State of the art of biological processes for coal gasification wastewater treatment. Biotechnol. Adv. 2016, 34, 1442. [Google Scholar] [CrossRef]
- Oberoi, A.S.; Philip, L.; Bhallamudi, S.M. Biodegradation of Various Aromatic Compounds by Enriched Bacterial Cultures: Part B-Nitrogen-, Sulfur-, and Oxygen-Containing Heterocyclic Aromatic Compounds. Appl. Biochem. Biot. 2015, 176, 1746–1769. [Google Scholar] [CrossRef]
- Zhuang, H.; Han, H.; Xu, P.; Hou, B.; Jia, S.; Wang, D.; Li, K. Biodegradation of quinoline by Streptomyces sp N01 immobilized on bamboo carbon supported Fe3O4 nanoparticles. Biochem. Eng. J. 2015, 99, 44–47. [Google Scholar] [CrossRef]
- Wu, B.; Wang, J.; Hu, Z.; Yuan, S.; Wang, W. Anaerobic biotransformation and potential impact of quinoline in an anaerobic methanogenic reactor treating synthetic coal gasification wastewater and response of microbial community. J. Hazard. Mater. 2020, 384, 121404. [Google Scholar] [CrossRef]
- Wang, J.; Han, L.; Shi, H.; Qian, Y. Biodegradation of quinoline by gel immobilized Burkholderia sp. Chemosphere 2001, 44, 1041–1046. [Google Scholar] [CrossRef]
- Wang, J.; Quan, X.; Han, L.; Qian, Y.; Hegemann, W. Microbial degradation of quinoline by immobilized cells of Burkholderia pickettii. Water Res. 2002, 36, 2288–2296. [Google Scholar] [CrossRef]
- Tuo, B.; Yan, J.; Fan, B.; Yang, Z.; Liu, J. Biodegradation characteristics and bioaugmentation potential of a novel quinoline-degrading strain of Bacillus sp. isolated from petroleum-contaminated soil. Bioresour. Technol. 2012, 107, 55–60. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, M.; Cheng, F.; Geng, Q. Biodegradation characterization and immobilized strains’ potential for quinoline degradation by Brevundimonas sp. K4 isolated from activated sludge of coking wastewater. Biosci. Biotechnol. Biochem. 2015, 79, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Dinamarca, M.A.; Eyzaguirre, J.; Baeza, P.; Aballay, P.; Canales, C.; Ojeda, J. A new functional biofilm biocatalyst for the simultaneous removal of dibenzothiophene and quinoline using Rhodococcus rhodochrous and curli amyloid overproducer mutants derived from Cobetia sp. strain MM1IDA2H-1. Biotechnol. Rep. 2018, 20, e286. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Chen, F.; Fu, J.; Sheng, G.; Sun, G. Microbial metabolism of quinoline by Comamonas sp. World J. Microb. Biot. 2004, 20, 539–543. [Google Scholar] [CrossRef]
- Sugaya, K.; Nakayama, O.; Hinata, N.; Kamekura, K.; Ito, A.; Yamagiwa, K.; Ohkawa, A. Biodegradation of quinoline in crude oil. J. Chem. Technol. Biot. 2001, 76, 603–611. [Google Scholar] [CrossRef]
- Bai, Y.; Sun, Q.; Zhao, C.; Wen, D.; Tang, X. Quinoline biodegradation and its nitrogen transformation pathway by a Pseudomonas sp strain. Biodegradation 2010, 21, 335–344. [Google Scholar] [CrossRef]
- Qiao, L.; Wang, J. Biodegradation characteristics of quinoline by Pseudomonas putida. Bioresour. Technol. 2010, 101, 7683–7686. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, J.; Zhang, Y.; Wang, Y.; Tong, L.; Hong, Q.; He, J. Biodegradation of Picolinic Acid by a Newly Isolated Bacterium Alcaligenes faecalis Strain JQ135. Curr. Microbiol. 2017, 74, 508–514. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Bao, Y.; Zhang, K.; Qiu, J.; He, Q.; Zhu, J.; He, J. Enhanced degradation of dicamba by an anaerobic sludge acclimated from river sediment. Sci. Total Environ. 2021, 777, 145931. [Google Scholar] [CrossRef]
- Zhu, S.; Liu, D.; Fan, L.; Ni, J. Degradation of quinoline by Rhodococcus sp. QL2 isolated from activated sludge. J. Hazard. Mater. 2008, 160, 289–294. [Google Scholar] [CrossRef]
- Oren, A. Thermodynamic limits to microbial life at high salt concentrations. Environ. Microbiol. 2011, 13, 1908–1923. [Google Scholar] [CrossRef]
- Luo, Y.; Yue, X.; Wei, P.; Zhou, A.; Kong, X.; Alimzhanova, S. A state-of-the-art review of quinoline degradation and technical bottlenecks. Sci. Total Environ. 2020, 747, 141136. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, J.; Feng, Y.C.; Bollag, J. Microbial metabolism of pyridine, quinoline, acridine, and their derivatives under aerobic and anaerobic conditions. Microbiol. Rev. 1996, 60, 483. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, G.; Bauder, R.; Speer, M.; Rommel, T.O.; Lingens, F. Microbial-metabolism of quinoline and related-compounds II. degradation of quinoline by Pseudomonas fluorescens 3, Pseudomonas-putida 86 and Rhodococcus spec b1. Biol. Chem. Hoppe-Seyler 1989, 370, 1183–1189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, L.; Wu, M.; Tang, Q.; Song, Z.; Zhou, H.; Bao, Y.; Liu, L.; Qu, Y. Comparative characterization and functional genomic analysis of two Comamonas sp. strains for biodegradation of quinoline. J. Chem. Technol. Biot. 2020, 95, 2017–2026. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Zhang, F.; Xu, S.; Yang, P.; Wang, X.; Zhang, X.; Hong, Q.; Qiu, J.; Chu, C.; He, J. Biodegradation of Quinoline by a Newly Isolated Salt-Tolerating Bacterium Rhodococcus gordoniae Strain JH145. Microorganisms 2022, 10, 797. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040797
Jiang Y, Zhang F, Xu S, Yang P, Wang X, Zhang X, Hong Q, Qiu J, Chu C, He J. Biodegradation of Quinoline by a Newly Isolated Salt-Tolerating Bacterium Rhodococcus gordoniae Strain JH145. Microorganisms. 2022; 10(4):797. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040797
Chicago/Turabian StyleJiang, Yinhu, Fuyin Zhang, Siqiong Xu, Pan Yang, Xiao Wang, Xuan Zhang, Qing Hong, Jiguo Qiu, Cuiwei Chu, and Jian He. 2022. "Biodegradation of Quinoline by a Newly Isolated Salt-Tolerating Bacterium Rhodococcus gordoniae Strain JH145" Microorganisms 10, no. 4: 797. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10040797