
Genetic Diversity of Babesia canis Strains in Dogs in Lithuania

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Extraction
2.2. PCR-RFLP Analysis
2.3. Sequence Analysis
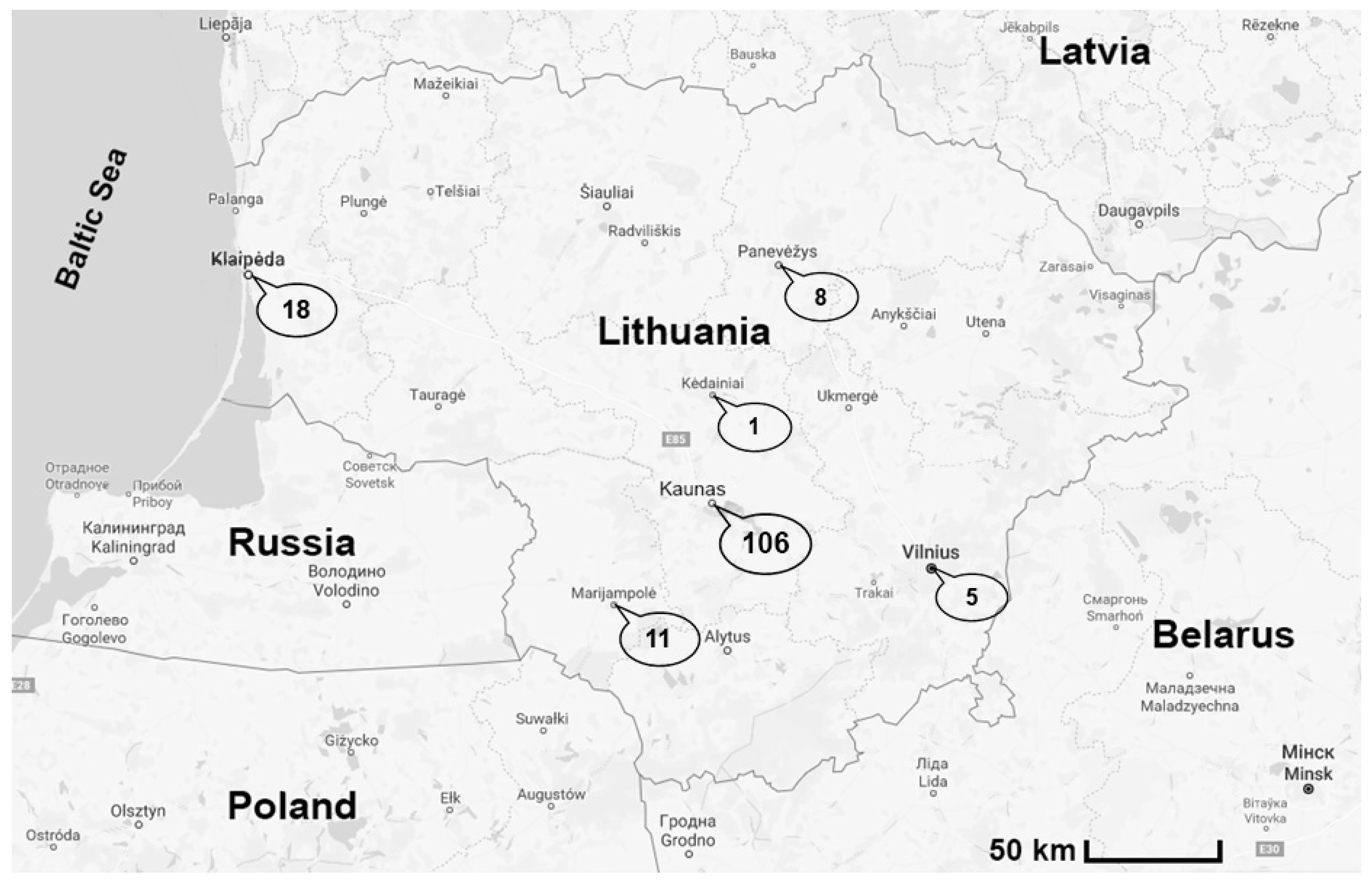
3. Results
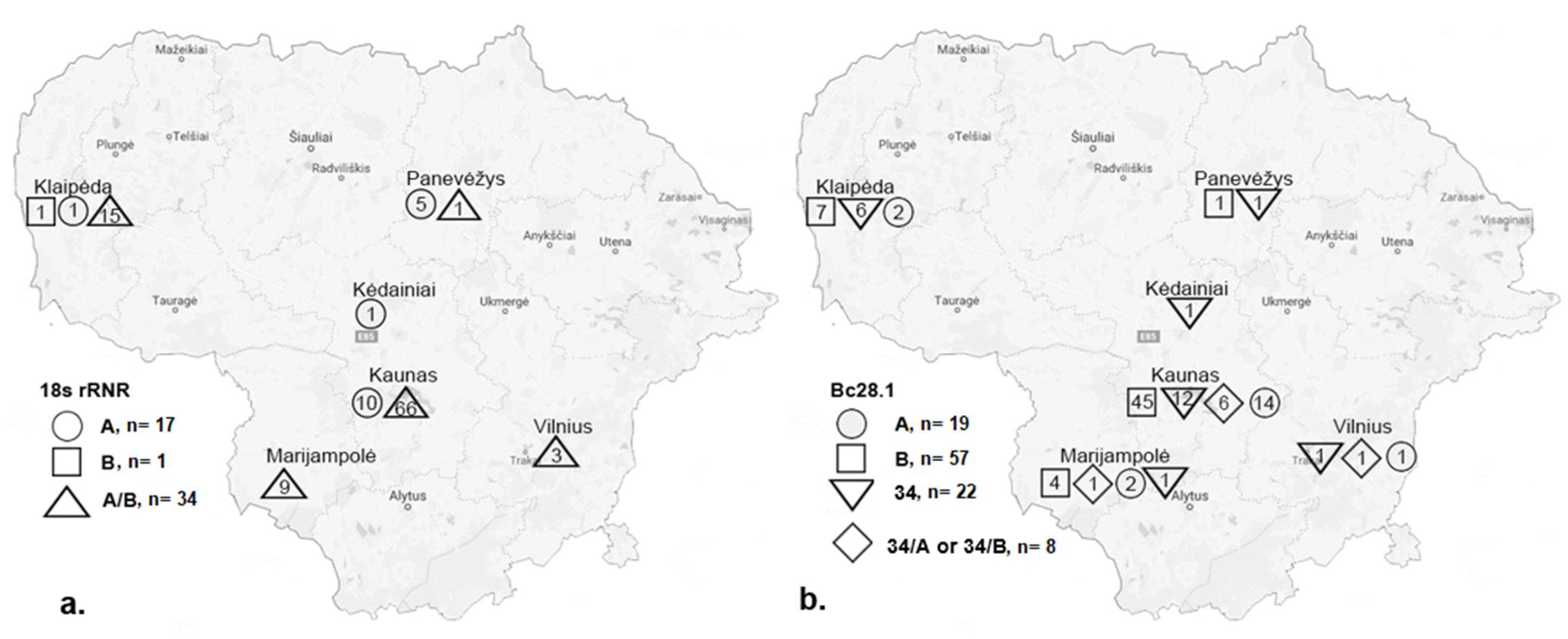
3.1. Babesia canis 18S rRNA Genotypes
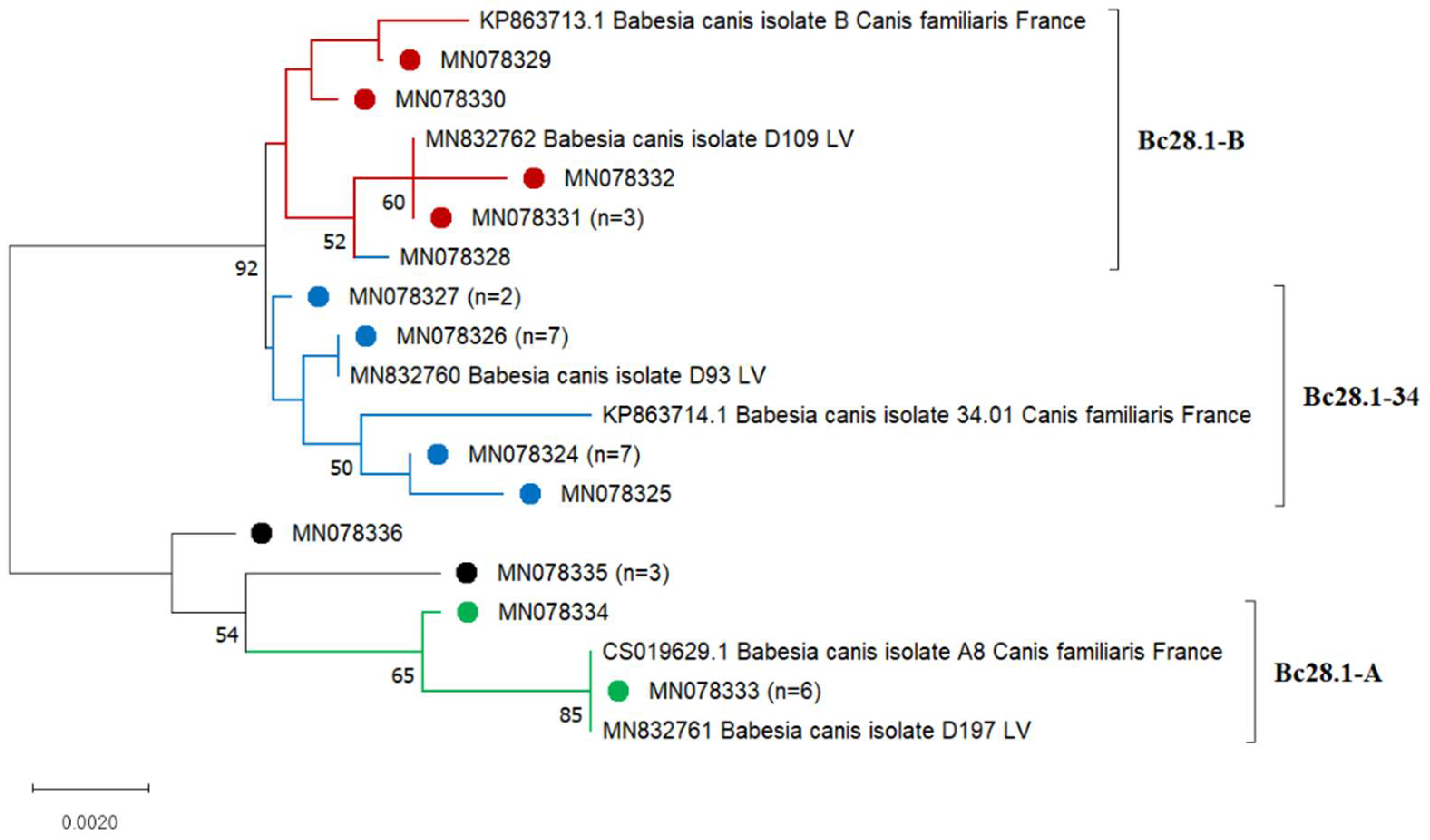
3.2. Babesia canis Bc28.1 Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Irwin, P.J. Canine babesiosis: From molecular taxonomy to control. Parasit. Vectors 2009, 2 (Suppl. S1), S4. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gallego, L.; Sainz, Á.; Roura, X.; Estrada-Peña, A.; Miró, G. A review of canine babesiosis: The European perspective. Parasit. Vectors 2016, 9, 336. [Google Scholar] [CrossRef] [PubMed]
- Matijatko, V.; Torti, M.; Schetters, T.P. Canine babesiosis in Europe: How many diseases? Trends Parasitol. 2012, 28, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Bajer, A.; Beck, A.; Beck, R.; Behnke, J.M.; Dwużnik-Szarek, D.; Eichenberger, R.M.; Farkas, R.; Fuehrer, H.P.; Heddergott, M.; Jokelainen, P.; et al. Babesiosis in Southeastern, Central and Northeastern Europe: An Emerging and Re-Emerging Tick-Borne Disease of Humans and Animals. Microorganisms 2022, 10, 945. [Google Scholar] [CrossRef]
- Berzina, I.; Capligina, V.; Baumanis, V.; Ranka, R.; Cirule, D.; Matise, I. Autochthonous canine babesiosis caused by Babesia canis canis in Latvia. Vet. Parasitol. 2013, 196, 515–518. [Google Scholar] [CrossRef]
- Paulauskas, A.; Radzijevskaja, J.; Karvelienė, B.; Grigonis, A.; Aleksandravičienė, A.; Zamokas, G.; Babickaitė, L.; Sabūnas, V.; Petkevičius, S. Detection and molecular characterization of canine babesiosis causative agent Babesia canis in naturally infected dog in Lithuania. Vet. Parasitol. 2014, 205, 702–706. [Google Scholar] [CrossRef]
- Seleznova, M.; Kivrane, A.; Namina, A.; Krumins, R.; Aleinikova, D.; Lazovska, M.; Akopjana, S.; Capligina, V.; Ranka, R. Babesiosis in Latvian domestic dogs, 2016–2019. Ticks Tick Borne Dis. 2020, 11, 101459. [Google Scholar] [CrossRef]
- Schäfer, I.; Helm, C.; Marsboom, C.; Hendrickx, G.; Kohn, B.; Krücken, J.; Samson-Himmelstjerna, G.; Müller, E. Infections with Babesia spp. in dogs living in Germany (2007–2020). J. Vet. Intern. Med. 2021, 35, 3199. [Google Scholar]
- Rubel, F.; Brugger, K.; Pfeffer, M.; Chitimia-Dobler, L.; Didyk, Y.M.; Leverenz, S.; Dautel, H.; Kahl, O. Geographical distribution of Dermacentor marginatus and Dermacentor reticulatus in Europe. Ticks Tick Borne Dis. 2016, 7, 224–233. [Google Scholar] [CrossRef]
- Drehmann, M.; Springer, A.; Lindau, A.; Fachet, K.; Mai, S.; Thoma, D.; Schneider, C.R.; Chitimia-Dobler, L.; Bröker, M.; Dobler, G.; et al. The spatial distribution of Dermacentor ticks (Ixodidae) in Germany—Evidence of a continuing spread of Dermacentor reticulatus. Front. Vet. Sci. 2020, 7, 578220. [Google Scholar] [CrossRef] [PubMed]
- Dwużnik-Szarek, D.; Mierzejewska, E.J.; Rodo, A.; Goździk, K.; Behnke-Borowczyk, J.; Kiewra, D.; Kartawik, N.; Bajer, A. Monitoring the expansion of Dermacentor reticulatus and occurrence of canine babesiosis in Poland in 2016–2018. Parasit. Vectors 2021, 14, 267. [Google Scholar] [CrossRef]
- Paulauskas, A.; Radzijevskaja, J.; Mardosaitė-Busaitienė, D.; Aleksandravičienė, A.; Galdikas, M.; Krikštolaitis, R. New localities of Dermacentor reticulatus ticks in the Baltic countries. Ticks Tick Borne Dis. 2015, 6, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Capligina, V.; Seleznova, M.; Akopjana, S.; Freimane, L.; Lazovska, M.; Krumins, R.; Kivrane, A.; Namina, A.; Aleinikova, D.; Kimsis, J.; et al. Large-scale countrywide screening for tick-borne pathogens in field-collected ticks in Latvia during 2017–2019. Parasit. Vectors 2020, 13, 351. [Google Scholar] [CrossRef] [PubMed]
- Radzijevskaja, J.; Mardosaitė-Busaitienė, D.; Aleksandravičienė, A.; Paulauskas, A. Investigation of Babesia spp. in sympatric populations of Dermacentor reticulatus and Ixodes ricinus ticks in Lithuania and Latvia. Ticks Tick Borne Dis. 2018, 9, 270–274. [Google Scholar] [CrossRef]
- Sabūnas, V. Prevalence of Vector-Borne Pathogens in Dogs. Ph.D. Thesis, Vytautas Magnus University, Kaunas, Lithuania, 2019. [Google Scholar]
- Carcy, B.; Randazzo, S.; Depoix, D.; Adaszek, L.; Cardoso, L.; Baneth, G.; Gorenflot, A.; Schetters, T.P. Classification of Babesia canis strains in Europe based on polymorphism of the Bc28.1-gene from the Babesia canis Bc28 multigene family. Vet. Parasitol. 2015, 211, 111–123. [Google Scholar] [CrossRef]
- Adaszek, Ł.; Winiarczyk, S. Molecular characterization of Babesia canis canis isolates from naturally infected dogs in Poland. Vet. Parasitol. 2008, 152, 235–241. [Google Scholar] [CrossRef]
- Yang, Y.S.; Murciano, B.; Moubri, K.; Cibrelus, P.; Schetters, T.; Gorenflot, A.; Delbecq, S.; Roumestand, C. Structural and functional characterization of Bc28.1, major erythrocyte-binding protein from Babesia canis merozoite surface. J. Biol. Chem. 2012, 287, 9495–9508. [Google Scholar] [CrossRef]
- Zhou, M.; Cao, S.; Luo, Y.; Liu, M.; Wang, G.; Moumouni, P.F.A.; Jirapattharasate, C.; Iguchi, A.; Vudriko, P.; Terkawi, M.A.; et al. Molecular identification and antigenic characterization of a merozoite surface antigen and a secreted antigen of Babesia canis (BcMSA1 and BcSA1). Parasit. Vectors 2016, 9, 257. [Google Scholar] [CrossRef]
- Eichenberger, R.M.; Riond, B.; Willi, B.; Hofmann-Lehmann, R.; Deplazes, P. Prognostic markers in acute Babesia canis infections. J. Vet. Intern. Med. 2016, 30, 174–182. [Google Scholar] [CrossRef]
- Beck, R.; Vojta, L.; Mrljak, V.; Marinculić, A.; Beck, A.; Živičnjak, T.; Cacciò, S.M. Diversity of Babesia and Theileria species in symptomatic and asymptomatic dogs in Croatia. Int. J. Parasitol. 2009, 39, 843–848. [Google Scholar] [CrossRef]
- Schaarschmidt, D.; Gilli, U.; Gottstein, B.; Marreros, N.; Kuhnert, P.; Daeppen, J.A.; Rosenberg, G.; Hirt, D.; Frey, C.F. Questing Dermacentor reticulatus harbouring Babesia canis DNA associated with outbreaks of canine babesiosis in the Swiss Midlands. Ticks Tick Borne Dis. 2013, 4, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Łyp, P.; Adaszek, Ł.; Furmaga, B.; Winiarczyk, S. Identification of new 18S rRNA strains of Babesia canis isolated from dogs with subclinical babesiosis. Pol. J. Vet. Sci. 2015, 18, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Hrazdilová, K.; Myśliwy, I.; Hildebrand, J.; Buńkowska-Gawlik, K.; Janaczyk, B.; Perec-Matysiak, A.; Modrý, D. Paralogs vs. genotypes? Variability of Babesia canis assessed by 18S rDNA and two mitochondrial markers. Vet. Parasitol. 2019, 266, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Helm, C.S.; Weingart, C.; Ramünke, S.; Schäfer, I.; Müller, E.; von Samson-Himmelstjerna, G.; Kohn, B.; Krücken, J. High genetic diversity of Babesia canis (Piana & Galli-Valerio, 1895) in a recent local outbreak in Berlin/Brandenburg, Germany. Transbound. Emerg. Dis. 2022. [Google Scholar] [CrossRef]
- Adaszek, Ł.; Winiarczyk, S.; Skrzypczak, M. The clinical course of babesiosis in 76 dogs infected with protozoan parasites Babesia canis canis. Pol. J. Vet. Sci. 2009, 12, 81–87. [Google Scholar]
- Uilenberg, G.; Franssen, F.F.; Perié, N.M.; Spanjer, A.A. Three groups of Babesia canis distinguished and a proposal for nomenclature. Vet. Q. 1989, 11, 33–40. [Google Scholar] [CrossRef]
- Kivrane, A.; Namina, A.; Seleznova, M.; Akopjana, S.; Capligina, V.; Ranka, R. Development of a real-time PCR method for rapid diagnosis of canine babesiosis and anaplasmosis. Parasit. Vectors 2021, 14, 266. [Google Scholar] [CrossRef]
- Paulauskas, A.; Galdikas, M.; Galdikaitė-Brazienė, E.; Stanko, M.; Kahl, O.; Karbowiak, G.; Radzijevskaja, J. Microsatellite-based genetic diversity of Dermacentor reticulatus in Europe. Infect. Genet. Evol. 2018, 66, 200–209. [Google Scholar] [CrossRef]
- Kloch, A.; Mierzejewska, E.J.; Karbowiak, G.; Slivinska, K.; Alsarraf, M.; Rodo, A.; Kowalec, M.; Dwużnik, D.; Didyk, Y.M.; Bajer, A. Origins of recently emerged foci of the tick Dermacentor reticulatus in central Europe inferred from molecular markers. Vet. Parasitol. 2017, 237, 63–69. [Google Scholar] [CrossRef]
- De Tommasi, A.S.; Otranto, D.; Dantas-Torres, F.; Capelli, G.; Breitschwerdt, E.B.; De Caprariis, D. Are vector-borne pathogen co-infections complicating the clinical presentation in dogs? Parasit. Vectors 2013, 6, 97. [Google Scholar] [CrossRef] [PubMed]
- Radzijevskaja, J.; Tamoliūnaitė, D.; Sabunas, V.; Aleksandravičienė, A.; Paulauskas, A. Prevalence and co-infection of mosquitoand tick-borne pathogens in domestic dogs suspected for canine babesiosis in Lithuania. Biologija 2020, 66, 2. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genotype | GenBank Accession Number (Sample Number) | Nucleotide Positions | ||
|---|---|---|---|---|
| 80 | 92 | 93 | ||
| A | MN078319 (n = 11) | T | G | A |
| MN078322 (n = 2) | Y | . | . | |
| B | MN078320 (n = 1) | . | A | G |
| A/B | MN078321 (n = 15) | . | R | R |
| MN078323 (n = 1) | Y | R | R | |
| Geootype | GenBank ace.No(Number of Samplcs) | Nucleotide Positions a,b | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 76 | 82 | 93 | 148 | 150 | 246 | 327 | 328 | 339 | 420 | 433 | 468 | 477 | 518 | 525 | 526 | 527 | 573 | 585 | 611 | ||
| A | CS019629 | G(D) | A(K) | G | G(E) | A | C | A(E) | G(D) | A(K) | C | C(L) | A(K) | T | C(T) | G | G | A | C | G | G(S) |
| MN078333 (n = 6) | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | |
| MN078334 (n = 1) | . | . | . | . | . | . | C(D) | A(N) | . | . | . | . | . | . | . | . | . | . | . | . | |
| B | KP863713 | G | C(Q) | G | G | A | T | C | A | T(N) | A | C | T(N) | C | C | G | G | A | A | G | G |
| MN078329 (n = 1) | . | . | . | . | . | . | . | . | . | . | . | . | . | A(N) | - | - | - | . | . | . | |
| MN078330 (n = 1) | . | . | . | . | . | . | . | . | . | . | . | . | . | A | - | - | - | C | . | . | |
| MN078331 (n = 3) | . | . | . | A(K) | . | . | . | . | . | . | . | . | . | A | - | - | - | C | . | . | |
| MN078332 (n = 1) | . | . | . | A | . | . | . | . | . | . | . | . | . | A | - | - | - | C | A | . | |
| 34 | KP863714 | A(N) | C | A | G | G | T | C | A | T(N) | A | G(V) | T | C | A | - | - | - | A | G | G |
| MN078324 (n = 7) | G | . | . | . | . | . | . | . | . | C | . | . | . | - | - | - | C | . | . | ||
| MN078325 (n = 1) | G | . | . | . | . | . | . | . | . | C | . | . | . | - | - | - | C | . | A(N) | ||
| MN078326 (n = 7) | G | . | G | . | . | . | . | . | . | C | . | . | . | - | - | - | . | . | . | ||
| MN078327 (n = 2) | G | . | G | . | . | . | . | . | . | C | . | . | . | - | - | - | C | . | . | ||
| MN078328 (n = 1) | G | . | G | A | . | . | . | . | . | C | . | . | . | - | - | - | C | . | . | ||
| 34/A | MN078335 (n = 3) | G | C | G | G | G | C | A | G | A | C | C | A | T | A | - | - | - | A | G | G |
| MN078336 (n = 1) | . | . | . | . | . | . | . | . | . | . | . | T | . | . | - | - | - | C | . | . | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radzijevskaja, J.; Mardosaitė-Busaitienė, D.; Aleksandravičienė, A.; Karvelienė, B.; Razgūnaitė, M.; Stadalienė, I.; Paulauskas, A. Genetic Diversity of Babesia canis Strains in Dogs in Lithuania. Microorganisms 2022, 10, 1446. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071446
Radzijevskaja J, Mardosaitė-Busaitienė D, Aleksandravičienė A, Karvelienė B, Razgūnaitė M, Stadalienė I, Paulauskas A. Genetic Diversity of Babesia canis Strains in Dogs in Lithuania. Microorganisms. 2022; 10(7):1446. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071446
Chicago/Turabian StyleRadzijevskaja, Jana, Dalytė Mardosaitė-Busaitienė, Asta Aleksandravičienė, Birutė Karvelienė, Miglė Razgūnaitė, Inga Stadalienė, and Algimantas Paulauskas. 2022. "Genetic Diversity of Babesia canis Strains in Dogs in Lithuania" Microorganisms 10, no. 7: 1446. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071446