
Molecular Identification of Babesia spp. and Anaplasma marginale in Water Buffaloes in Veracruz and Tabasco, Mexico: A Retrospective Study

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
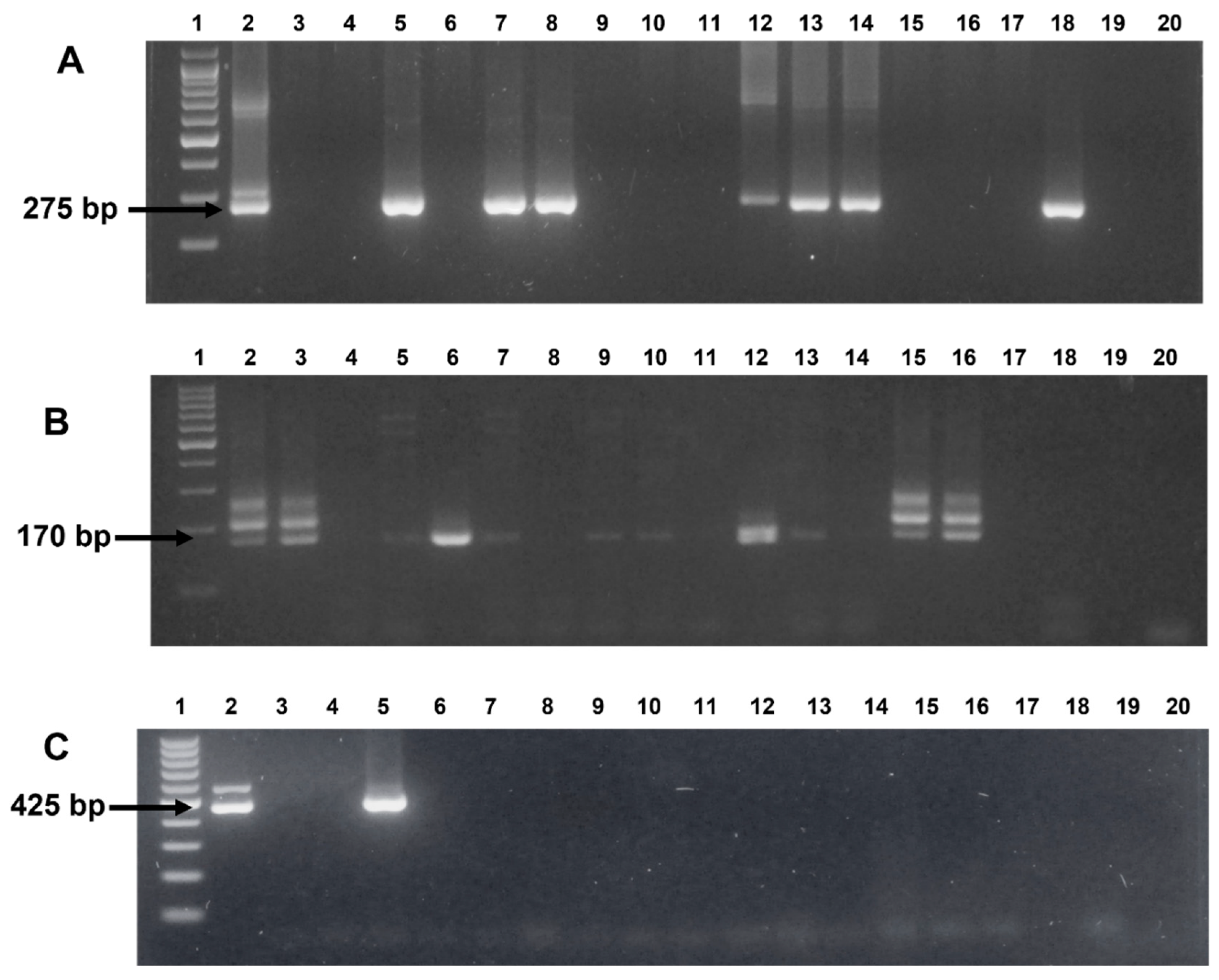
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- SIAP. Población Ganadera|Servicio de Información Agroalimentaria y Pesquera|Gobierno|gob.mx. Available online: www.gob.mx (accessed on 30 June 2022).
- Rojas-Martinez, C.; Loza-Rubio, E.; Rodriguez-Camarillo, S.D.; Figueroa-Millan, J.V.; Aguilar-Romero, F.; Lagunes-Quintanilla, R.E.; Morales-Alvarez, J.F.; Santillan-Flores, M.A.; Socci-Escatell, G.B.; Alvarez-Martinez, J.A. Background and perspectives of certain priority diseases affecting cattle farming in Mexico. Rev. Mex. Cienc. Pecu. 2021, 12, 111–148. [Google Scholar] [CrossRef]
- Urdaneta, F. Mejoramiento de la eficiencia productiva de los sistemas de ganaderia doble proposito (Taurus-Indicus). Arch. Lat. Prod Anim. 2009, 17, 109–120. [Google Scholar]
- Rodriguez-Vivas, R.I.; Grisi, L.; Perez de Leon, A.A.; Silva-Villelad, H.; Torres-Acosta, J.F.J.; Fragoso-Sanchez, H.; Romero-Salas, D.; Rosario-Cruz, R.; Saldierna, F.; Garcia-Carrasco, D. Potential economic impact assessment for cattle parasites in Mexico. Review. Rev. Mex. Cienc. Pecu. 2017, 8, 61–74. [Google Scholar] [CrossRef]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitol 2004, 129, S3–S14. [Google Scholar] [CrossRef]
- Marcelino, I.; de Almeida, A.M.; Ventosa, M.; Pruneau, L.; Meyer, D.F.; Martinez, D.; Lefrançois, T.; Vachiéry, N.; Coelho, A.V. Tick-borne diseases in cattle: Applications of proteomics to develop new generation vaccines. J. Proteom. 2012, 75, 4232–4350. [Google Scholar] [CrossRef]
- Ganzinelli, S.; Rodriguez, A.E.; Schnittger, L.; Florin-Christensen, M. Babesia in domestic ruminants. In Parasitic Protozoa of Farm Animals and Pets, 1st ed.; Florin-Christensen, M., Schnittger, L., Eds.; Springer International Publishing AG: Cham, Switzerland, 2018; pp. 215–239. [Google Scholar] [CrossRef]
- Bhat, S.A.; Singh, N.K.; Singh, H.; Rath, S.S. Molecular prevalence of Babesia bigemina in Rhipicephalus microplus ticks infesting cross-bred cattle of Punjab, India. Parasite Epidemiol. Control 2017, 2, 85–90. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Rojas, C.; Figueroa, J.V. Diagnostic tools for the identification of Babesia sp. in persistently infected cattle. Pathogens 2019, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Ocampo, R.; Vega y Murguia, C.A.; Oviedo-Ortega, O.N.; Rojas-Ramirez, E.E.; Garcia-Ortiz, M.A.; Preciado de la Torre, J.F.; Rosario-Cruz, R.; Dominguez-Garcia, D.I.; Rodriguez-Camarillo, S.D. Diversidad genetica de la region variable de los genes msp1a y msp4 en cepas de Anaplasma marginale de Mexico. Rev. Mex. Cienc. Pecu. 2012, 3, 373–387. [Google Scholar]
- Guswanto, A.; Allamanda, P.; Mariamah, E.S.; Sodirun, S.; Wibowo, P.E.; Indrayani, L.; Nugroho, R.H.; Wirata, I.K.; Jannah, N.; Dias, L.P.; et al. Molecular and serological detection of bovine babesiosis in Indonesia. Parasit. Vectors 2017, 10, 550. [Google Scholar] [CrossRef] [Green Version]
- Terkawi, M.A.; Huyen, N.X.; Shinuoa, C.; Inpankaew, T.; Maklon, K.; Aboulaila, M.; Ueno, A.; Goo, Y.K.; Yokoyama, N.; Jittapalapong, S.; et al. Molecular and serological prevalence of Babesia bovis and Babesia bigemina in water buffaloes in the northeast region of Thailand. Vet. Parasitol. 2011, 178, 201–207. [Google Scholar] [CrossRef]
- Obregon, D.; Cabezas-Cruz, A.; Armas, Y.; Silva, J.B.; Fonseca, A.H.; Andre, M.R.; Alfonso, P.; Oliveira, M.C.S.; Machado, R.Z.; Corona-González, B. High co-infection rates of Babesia bovis, Babesia bigemina, and Anaplasma marginale in water buffalo in Western Cuba. Parasitol. Res. 2019, 118, 955–967. [Google Scholar] [CrossRef]
- Aubry, P.; Geale, D.W. A review of bovine anaplasmosis. Transbound. Emerg. Dis. 2011, 58, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Millan, J.V.; Castañeda-Arriola, R.O.; Lira-Amaya, J.J.; Vargas-Uriostegui, P.; Alvarez-Martinez, J.A.; Rojas-Martinez, C. ELISA test using Babesia sp. recombinant antigens for epidemiological diagnosis of bovine babesiosis in Mexico. In ELISA: Advances in Research and Application, 1st ed.; Angeles, J.M., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2022; pp. 49–67. [Google Scholar]
- Figueroa, J.V.; Chieves, L.P.; Johson, G.S.; Buening, G.M. Polymerase chain reaction-based assay for the detection of Babesia bigemina, Babesia bovis and Anaplasma marginale DNA in bovine blood. Vet. Parasitol. 1993, 50, 69–81. [Google Scholar] [CrossRef]
- Figueroa, J.V.; Chieves, L.P.; Johnson, G.S.; Buening, G.M. Detection of Babesia bigemina-infected carriers by polymerase reaction amplification. J. Clin. Microbiol. 1992, 30, 2576–2582. [Google Scholar] [CrossRef] [Green Version]
- Torioni de Echaide, S.; Knowles, D.P.; McGuire, T.C.; Palmer, G.H.; Suarez, C.E.; McElwain, T.F. Detection of cattle naturally infected with Anaplasma marginale in a region of endemicity by nested PCR and a competitive enzyme-linked immunosorbent assay using recombinant major surface protein 5. J. Clin. Microbiol. 1998, 36, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, E.C.; Romero, S.D.; Garcia, V.Z.; Cruz, R.A.; Peniche, C.A.; Ibarra, P.N.; Aguilar, D.M.; Perez, D.L.A.; Dubey, J.P. Seroprevalence of Toxoplasma gondii infection in water buffaloes (Bubalus bubalis) in Veracruz State, Mexico, and its association with climatic factors. BMC Vet. Res. 2014, 10, 232. [Google Scholar]
- Salguero-Romero, J.L.; Romero-Salas, D.; Cruz-Romero, A.; Alonso-Diaz, M.A.; Aguilar-Dominguez, M.; Sanchez-Montes, S.; Lammoglia-Villagómez, M.A.; Chaparro-Gutierrez, J.J.; Nogueira-Domingues, L.; Perez-de Leon, A.A.; et al. Serosurveillance and risk factors for Neospora caninum infection in water buffaloes (B. bubalis) from central and southern Mexico. Vet. Parasitol. Reg. Stud. Rep. 2021, 26, 100631. [Google Scholar] [CrossRef] [PubMed]
- Corona, B.; Obregon, D.; Martinez, S.; Espinosa, I.; Fonseca, A.H.; Roque, A. Deteccion por PCR de Anaplasma marginale en bufalos de la region occidental de Cuba. Rev. Salud. Anim. 2012, 34, 11–18. [Google Scholar]
- Obregon, D.; De Sena, O.M.C.; Tizioto, P.C.; Funnes, M.E.; Martinez, S.; Roquel, E.; Fonseca, A.H.; Corona, B. Diagnostico de Babesia bovis en bufalos de la region occidental de Cuba a traves de un ensayo de nPCR. Rev. Salud. Anim. 2012, 34, 101–108. [Google Scholar]
- Viana, R.V.; Wallis, C.L. Good Clinical Laboratory Practice (GCLP) for Molecular Based Tests Used in Diagnostic Laboratories. In Wide Spectra of Quality Control Akyar, 1st ed.; InTech: London, UK, 2011; Available online: http://www.intechopen.com/books/wide-spectra-of-quality-control/goodclinical-laboratory-practice-gclp-for-molecular-based-tests-used-in-diagnostic-laboratories (accessed on 7 June 2022).
- Genis, A.D.; Perez, J.; Mosqueda, J.J.; Alvarez, A.; Camacho, M.; Muñoz, M.D.L.; Rojas, C.; Figueroa, J.V. Using msa-2b as a molecular marker for genotyping Mexican isolates of Babesia bovis. Infect. Genet. Evol. 2009, 9, 1102–1107. [Google Scholar] [CrossRef]
- Suarez, C.E.; Palmer, G.H.; Hotzel, I.; McElwain, T.F. Structure, sequence, and transcriptional analysis of the Babesia bovis rap-1 multigene locus. Mol. Biochem. Parasitol. 1998, 93, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, T.; Tattiyapong, M.; Fukushi, S.; Hayashida, K.; Kothalawala, H.; Silva, S.S.P.; Vimalakumar, S.C.; Kanagaratnam, R.; Meewewa, A.S.; Suthaharan, K.; et al. Genetic characterization of Babesia and Theileria parasites in water buffaloes in Sri Lanka. Vet. Parasitol. 2014, 200, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.G.; Henriques, G.; Sanchez, C.; Marques, P.X.; Suarez, C.E.; Oliva, A. First survey for Babesia bovis and Babesia bigemina infection in cattle from Central and Southern regions of Portugal using serological and DNA detection methods. Vet. Parasitol. 2009, 166, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Zhyldyz, A.; Sivakumar, T.; Igarashi, I.; Gunasekara, E.; Kothalawala, H.; Silva, S.S.P.; Yokoyama, N. Epidemiological survey of Anaplasma marginale in cattle and buffalo in Sri Lanka. J. Vet. Med. Sci. 2019, 81, 1601–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, J.V.; Alvarez, J.A.; Canto, G.J.; Ramos, J.A.; Mosqueda, J.J.; Buening, G.M. Comparative sensitivity of two tests for the diagnosis of multiple hemoparasite infection of cattle. Ann. N. Y. Acad. Sci. 1996, 23, 117–127. [Google Scholar] [CrossRef]
- Oliveira-Sequeira, T.C.; Oliveira, M.C.; Araujo, J.P., Jr.; Amarante, A.F. PCR-based detection of Babesia bovis and Babesia bigemina in their natural host Boophilus microplus and cattle. Int. J. Parasitol. 2005, 35, 105–111. [Google Scholar] [CrossRef] [Green Version]
- OIE World Organization for Animal Health (WOAH). Bovine Babesiosis; OIE WOAH Terrestrial Manual: Paris, France, 2021; Chapter 3.4.2; Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.04.02_BABESIOSIS.pdf (accessed on 28 June 2022).
- Sivakumar, T.; Altangerel, K.; Battsetseg, B.; Battur, B.; Aboulaila, M.; Munkhjargal, T.; Yoshinari, T.; Yokoyama, N.; Igarashi, I. Genetic detection of Babesia bigemina from Mongolian cattle using apical membrane antigen-1 gene based PCR assay. Vet. Parasitol. 2012, 187, 17–22. [Google Scholar] [CrossRef]
- OIE World Organization for Animal Health. Bovine Anaplasmosis; OIE WOAH Terrestrial Manual: Paris, France, 2018; Chapter 3.4.1; Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/3.04.01_BOVINE_ANAPLASMOSIS.pdf (accessed on 28 June 2022).
- Parodi, P.; Corbellini, L.G.; Leotti, V.B.; Rivero, R.; Miraballes, C.; Riet-Correa, F.; Venzal, J.M.; Armúa-Fernández, M.T. Validation of a multiplex PCR assay to detect Babesia spp. and Anaplasma marginale in cattle in Uruguay in the absence of a gold standard test. J. Vet. Diagn. Invest. 2021, 33, 73–79. [Google Scholar] [CrossRef]
- Obregon, D.; Corona-Gonzalez, B.; Diaz-Sanchez, A.A.; Armas, Y.; Roque, E.; de Sena Oliveira, M.C.; Cabezas-Cruz, A. Efficient transovarial transmission of Babesia spp. in Rhipicephalus microplus ticks fed on water buffalo (Bubalus bubalis). Pathogens 2020, 9, 280. [Google Scholar] [CrossRef]
- Romero-Salas, D.; Mira, A.; Mosqueda, J.; Garcia-Vazquez, Z.; Hidalgo-Ruiz, M.; Vela, N.A.O.; de León, A.A.; Florin-Christensen, M.; Schnittger, L. Molecular and serological detection of Babesia bovis and Babesia bigemina-infection in bovines and water buffaloes raised jointly in an endemic field. Vet. Parasitol. 2016, 217, 101–107. [Google Scholar] [CrossRef] [Green Version]
- da Silva, J.B.; Andre, M.R.; da Fonseca, A.H.; Albuquerque-Lopez, C.T.; da Silva-Lima, D.H.; Tavares de Andrade, S.J.; Oliveira, C.M.; Barbosa, J.D. Molecular and serological prevalence of Babesia bovis and Babesia bigemina in water buffaloes in the north region of Brazil. Vet. Parasitol. 2013, 197, 678–681. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.S.; Kandil, O.M.; Nasr, S.M.; Hendawy, S.; Habeeb, S.; Mabrouk, D.; Silva, M.G.; Suarez, C.E. Serological and molecular diagnostic surveys combined with examining hematological profiles suggests increased levels of infection and hematological response of cattle to babesiosis infections compared to native buffaloes in Egypt. Parasit. Vectors 2015, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AL-Hosary, A.; Raileanu, C.; Tauchmann, O.; Fischer, S.; Nijhof, A.M.; Silaghi, C. Epidemiology and genotyping of Anaplasma marginale and co-infection with piroplasms and other Anaplasmataceae in cattle and buffaloes from Egypt. Parasit. Vectors 2020, 13, 495. [Google Scholar] [CrossRef] [PubMed]
- Obregon, D.; Corona, B.G.; de la Fuente, J.; Cabezas-Cruz, A.; Goncalves, L.R.; Matos, C.A.; Armas, Y.; Hinojosa, Y.; Alfonso, P.; Oliveira, M.C.S.; et al. Molecular evidence of the reservoir competence of water buffalo (Bubalus bubalis) for Anaplasma marginale in Cuba. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Galon, E.M.S.; Adjou Moumouni, P.F.; Ybañez, R.H.D.; Ringo, A.E.; Efstratiou, A.; Lee, S.H.; Salces, C.B.; Maurillo, B.C.A.; Boldbaatar, D.; Ybañez, A.P.; et al. First molecular detection and characterization of tick-borne pathogens in water buffaloes in Bohol, Philippines. Ticks Tick Borne Dis. 2019, 10, 815–821. [Google Scholar] [CrossRef]
- Otgonsuren, O.; Sivakumar, T.; Amgalanbaatar, T.; Enkhtaivan, B.; Narantsatsral, S.; Tuvshintulga, B.; Zoljargal, M.; Munkhgerel, D.; Davkharbayar, B.; Baatarjargal, P.; et al. Molecular epidemiological survey of Babesia bovis, Babesia bigemina, and Babesia sp. Mymensingh infections in Mongolian cattle. Parasitol. Inter. 2020, 77, 102107. [Google Scholar] [CrossRef]
- Mtshali, P.S.; Tsotetsi, A.M.; Thekisoe, M.M.O.; Mtshali, M.S. Nested PCR detection and phylogenetic analysis of Babesia bovis and Babesia bigemina in cattle from Peri-urban localities in Gauteng Province, South Africa. J. Vet. Med. Sci. 2014, 76, 145–150. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Species | Assay | Primer Sequences (5′ > 3′) | Product Size (bp) | Reference |
|---|---|---|---|---|
| B. bovis | PCR | CGAGGAAGGAACTACCGATG | 354 | [16] |
| GGAGCTTCAACGTACGAGGT | ||||
| nPCR | TGGCTACCATGAACTACAAGACTTA | 275 | [16] | |
| GAGCAGAACCTTCTTCACCAT | ||||
| B. bigemina | PCR | CATCTAATTTCTCTCCATACCCC | 278 | [17] |
| CCTCGGCTTCAACTCTGATGCC | ||||
| nPCR | CGCAAGCCCAGCACGCCCCGGT | 170 | [17] | |
| CCGACCTGGATAGGCTGTGATG | ||||
| A. marginale | PCR | ACCTTCTGCTGTTCGTTGC | 628 | [18] |
| TGTACCACTGCCATGCTTAAG | ||||
| nPCR | CATAGCCTCCGCGTCTTT | 425 | [18] | |
| CTTAAACAGCTCCTCGCCTT |
| Production Unit | B. bovis | B. bigemina | A. marginale | |||
|---|---|---|---|---|---|---|
| + | % | + | % | + | % | |
| Veracruz 1, n = 61 | 23 | (37.7) | 6 | (9.8) | 1 | (1.6) |
| Veracruz 2, n = 63 | 13 | (20.6) | 7 | (11.1) | 4 | (6.3) |
| Tabasco 1, n = 58 | 35 | (60.3) | 8 | (13.7) | 0 | (0) |
| Tabasco 2, n = 58 | 21 | (41.1) | 12 | (23.5) | 1 | (1.9) |
| Total = 233 | 92 | (39.5) | 33 | (14.2) | 6 | (2.6) |
| Sample ID | Parasite Sample Source | Accession Number (GenBank) | Highest DNA Sequence Identity (%) | Accession Number (GenBank) |
|---|---|---|---|---|
| B. bovis | ||||
| +Control | Bovine blood | OP174624 | 275/275 (100) | AF030056.2 |
| 4.1 | Veracruz | ON862140 | 273/275 (99) | AF030056.2 |
| 17.1 | “ | ON862141 | 274/275 (99) | JF279443.1 |
| 105.1 | “ | ON862142 | 275/275 (100) | AB845432.1 |
| 33.1 | “ | ON862143 | 275/275 (100) | AB845432.1 |
| 50.5 | “ | ON862144 | 275/275 (100) | AB845432.1 |
| 83.5 | “ | ON862145 | 275/275 (100) | AB845432.1 |
| 25.1 | Tabasco | ON862146 | 274/275 (99) | JF279443.1 |
| 28.3 | “ | ON862147 | 274/275 (99) | AB845432.1 |
| 33.1 | “ | ON862148 | 274/275 (99) | JF279443.1 |
| 62.1 | “ | ON862149 | 275/275 (100) | AB845432.1 |
| 75.1 | “ | ON862150 | 275/275 (100) | JF279443.1 |
| 94.2 | “ | ON862151 | 275/275 (100) | AB845432.1 |
| B. bigemina | ||||
| +Control | Bovine blood | OP174630 | 171/171 (100) | S45366.1 |
| 57.1 | Veracruz | ON862152 | 168/171 (98) | S45366.1 |
| 68.1 | “ | ON862153 | 168/171 (98) | S45366.1 |
| 73.1 | “ | ON862154 | 168/171 (98) | S45366.1 |
| 30.1 | “ | ON862155 | 165/170 (97) | S45366.1 |
| 100.1 | “ | ON862156 | 168/171 (98) | S45366.1 |
| 6.1 | Tabasco | ON862157 | 165/171 (96) | S45366.1 |
| 23.1 | “ | ON862158 | 168/173 (97) | S45366.1 |
| 27.1 | “ | ON862159 | 168/171 (98) | S45366.1 |
| 71.1 | “ | ON862160 | 167/171 (98) | S45366.1 |
| 75.1 | “ | ON862161 | 80/95 (88) | FJ939724.1 |
| 88.1 | “ | ON862162 | 167/171 (98) | S45366.1 |
| A. marginale | ||||
| +Control | Bovine blood | OP174628 | 425/425 (100) | LC467711.1 |
| 218.3 | Veracruz | ON862163 | 408/414 (99) | LC467711.1 |
| 236.120 | “ | ON862164 | 415/423 (98) | LC467711.1 |
| 220.117 | “ | ON862165 | 423/426 (99) | LC467711.1 |
| 235.32 | “ | ON862166 | 303/314 (96) | LC467711.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lira-Amaya, J.J.; Santamaria-Espinosa, R.M.; Castañeda-Arriola, R.O.; Martínez-García, G.; Polanco-Martínez, D.J.; Rojas-Martínez, C.; Alvarez-Martínez, J.Á.; Figueroa-Millán, J.V. Molecular Identification of Babesia spp. and Anaplasma marginale in Water Buffaloes in Veracruz and Tabasco, Mexico: A Retrospective Study. Microorganisms 2022, 10, 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091702
Lira-Amaya JJ, Santamaria-Espinosa RM, Castañeda-Arriola RO, Martínez-García G, Polanco-Martínez DJ, Rojas-Martínez C, Alvarez-Martínez JÁ, Figueroa-Millán JV. Molecular Identification of Babesia spp. and Anaplasma marginale in Water Buffaloes in Veracruz and Tabasco, Mexico: A Retrospective Study. Microorganisms. 2022; 10(9):1702. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091702
Chicago/Turabian StyleLira-Amaya, José Juan, Rebeca Montserrat Santamaria-Espinosa, Roberto O. Castañeda-Arriola, Grecia Martínez-García, Diego J. Polanco-Martínez, Carmen Rojas-Martínez, Jesús Ántonio Alvarez-Martínez, and Julio V. Figueroa-Millán. 2022. "Molecular Identification of Babesia spp. and Anaplasma marginale in Water Buffaloes in Veracruz and Tabasco, Mexico: A Retrospective Study" Microorganisms 10, no. 9: 1702. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091702