
Bioactivities of Ketones Terpenes: Antifungal Effect on F. verticillioides and Repellents to Control Insect Fungal Vector, S. zeamais

Abstract
:
1. Introduction
2. Experimental Section
2.1. Chemicals

2.2. Antifungal Activity of Ketones
2.2.1. Fungal Culture
2.2.2. Medium and Culture Conditions
2.2.3. Growth Assessment
2.3. Molecular Modeling and Calculation of Molecular Parameters
2.4. Insect Growth
2.5. Repellency Bioassays
2.6. Statistical Analysis
3. Results
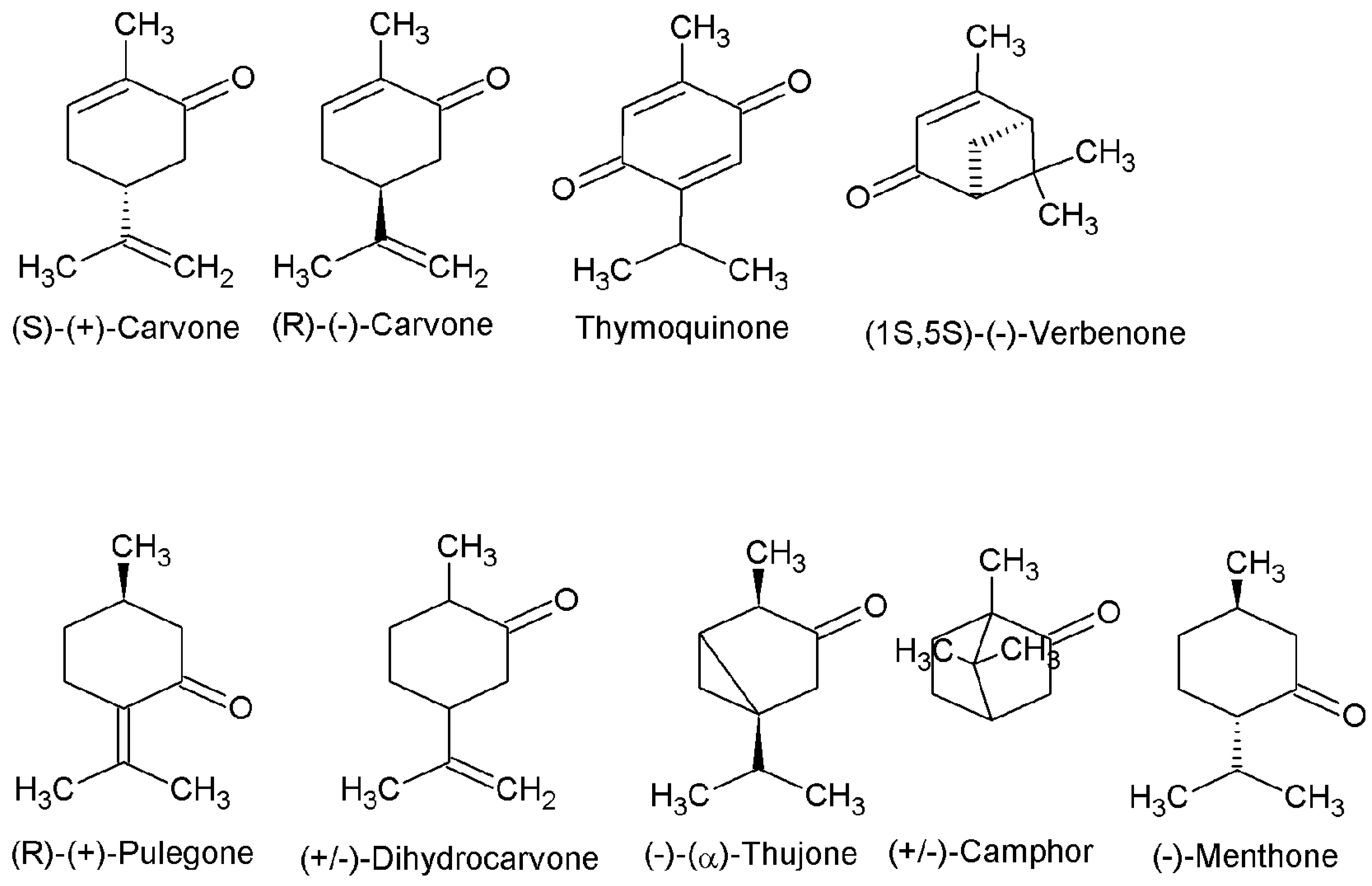
3.1. Antifungal Activity of Ketones


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | MIC A | LD25 B | Inhibition (%) ± SD C | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Concentration (mM) | |||||||||
| 0.25 | 0.5 | 0.75 | 1 | 1.5 | 2 | 3 | |||
| thymoquinone | 0.87 | 0.23 | 23.7 ± 18.1 *,d | 60.7 ± 41.1 * c | 89.7 ± 11.9 *,a | 75.2 ± 18.6 *,a | 85.3 ± 18.7 *,a | 100 ± 0 *,a | 99.4 ± 1.2 *,a |
| R-carvone | 4.56 | 1.02 | 7.6 ± 10.0 e | 13.7 ± 9.3 e | 21.0 ± 5.5 *,d | 22.3 ± 13.2 *,d | 38.4 ± 7.6 *,c | 45.3 ± 7.5 *,c | 66.2 ± 8.3 *,d |
| S-carvone | 3.84 | 1.48 | 0 f | 1.5 ± 2.9 f | 7.3 ± 8.9 f | 13.1 ± 11.3 e | 38.6 ± 12.1 *,c | 58.8 ± 7.4 *,b | 42.0 ± 11.1 *,c |
| pulegone | 6.38 | 1.57 | 1.2 ± 1.4 f | 13.8 ± 5.3 * e | 10.9 ± 11.4 e | 15.7 ± 7.0 *,e | 21.6 ± 10.0 *,d | 34.5 ± 4.5 *,c | 46.6 ± 5.8 *,c |
| dihydrocarvone | 13.57 | 4.19 | 0 f | 0 f | 0 f | 0.6 ± 1.2 f | 1.2 ± 1.4 f | 8.6 ± 4.6 e | 2.3 ± 2.7 f |
| menthone | ND | ND | 0 f | 0 f | 0 f | 0 f | 1.5 ± 2.9 f | 0 f | 0 f |
| verbenone | 4.90 | 2.43 | 0 f | 1.2 ± 2.3 f | 1.8 ± 2.2 f | 0.6 ± 1.2 f | 1.5 ± 1.8 f | 4.9 ± 6.1 f | 44.6 ± 11.7 *,c |
| α-thujone | ND | ND | 0 f | 0 f | 0.6 ± 1.2 f | 0 f | 0 f | 0 f | 0 f |
| camphor | 26.64 | 7.94 | 0 f | 0 f | 0 f | 0 f | 0 f | 0 f | 4.3 ± 5.0 f |
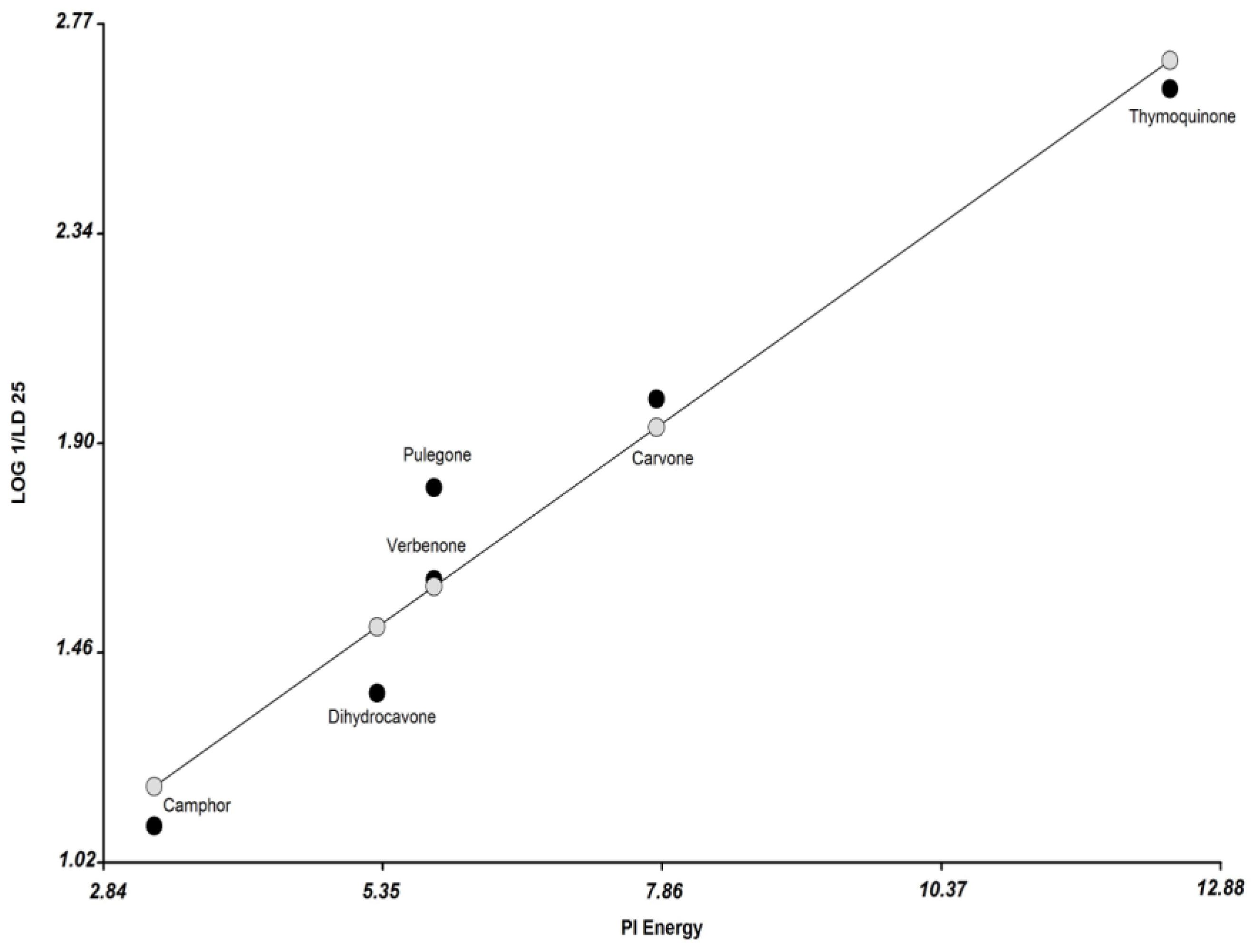
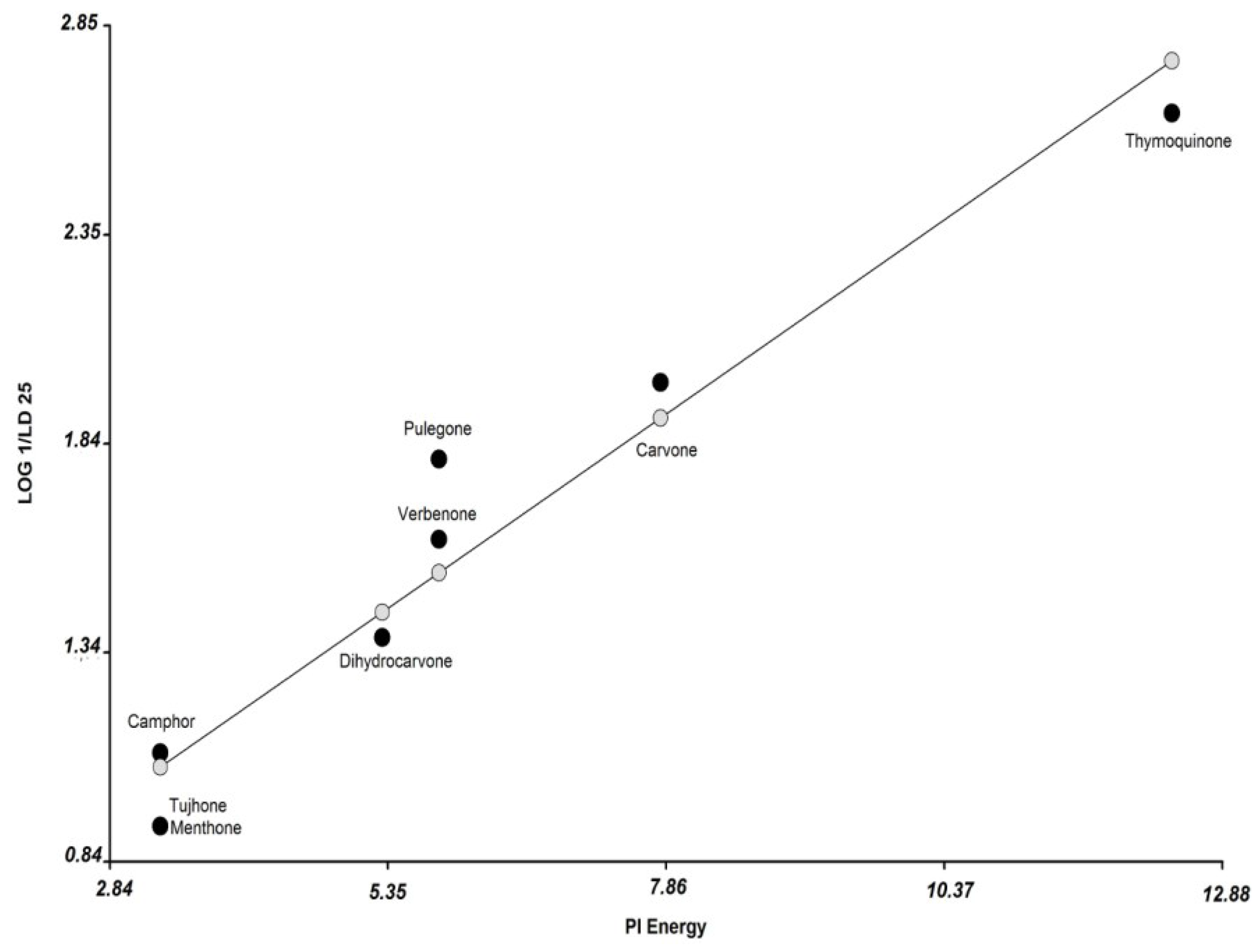
3.2. Molecular Modeling and Calculation of Molecular Parameters

| Compounds | Dreiding Energy kcal/mol | Dipolo (Debye) | Molar Refractivity | Surface Tension (dyne/cm) | Volume (A3) | Minimal Projection Area (A2) | Maximal Projection Area (A2) | Molar Volume (cm3) | Log P | Pi Energy | Boiling Point C HectoPascal | Polarizability ×10−24 cm3) | Enthalpy of Vaporization (Kj/mol) | Polar Surface Area (A2) | Solvent Accessible Surface Area | pKa |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| thymoquinone | 34.46 | 0.12 | 48.89 | 35.8 | 159 | 32.29 | 53.09 | 154.1 | 2.33 | 12.42 | 309 | 18.1 | 46.9 | 34.14 | 245.95 | −7.75 |
| carvone | 35 | 3.2 | 47.17 | 29.8 | 156.9 | 31.02 | 50.97 | 159.8 | 2.27 | 7.81 | 307 | 18 | 46.7 | 17.07 | 248.75 | −4.66 |
| pulegone | 40.89 | 2.88 | 47.13 | 29.5 | 164.5 | 31.96 | 53.06 | 164.8 | 2.56 | 5.81 | 298 | 18.2 | 46 | 17.07 | 277.53 | −4.43 |
| dihydrocarvone | 27.07 | 2.79 | 46.3 | 27.98 | 164.9 | 31.45 | 51.84 | 168.47 | 3.22 | 5.3 | 294 | 18.19 | 45.79 | 17.07 | 274.86 | −7.42 |
| menthone | 27.39 | 2.73 | 46.52 | 27.3 | 172.6 | 30.28 | 57.81 | 175.1 | 2.63 | 3.3 | 273 | 18.4 | 44.1 | 17.07 | 300.97 | −7.42 |
| verbenone | 92.48 | 3.79 | 45.37 | 29.46 | 154.3 | 34.84 | 46.84 | 151.4 | 2.14 | 5.81 | 302 | 17.48 | 46.41 | 17.07 | 253.13 | −4.73 |
| thujone | 146.8 | 2.76 | 44.54 | 35 | 160.1 | 34.23 | 47.95 | 150.8 | 1.9 | 3.3 | 266 | 17.6 | 43.7 | 17.07 | 283.79 | −7.42 |
| camphor | 51.95 | 3.02 | 44.49 | 31.6 | 161.3 | 37.48 | 42.7 | 154.9 | 2.13 | 3.3 | 275 | 17.6 | 44.4 | 17.07 | 284.25 | −7.49 |
| Compounds | Platt Index | Randic Index | Balaban Index | Harary Index | Wiener Index | Hyper Wiener Index | Wiener Polarity | Szeged Index |
|---|---|---|---|---|---|---|---|---|
| thymoquinone | 34 | 5.52 | 2.55 | 30.7 | 187 | 417 | 17 | 292 |
| Carvone * | 30 | 5.11 | 2.18 | 26.5 | 152 | 337 | 17 | 240 |
| pulegone | 30 | 5.11 | 2.22 | 26.6 | 150 | 327 | 14 | 236 |
| dihydrocarvone | 30 | 5.11 | 2.18 | 26.5 | 152 | 337 | 14 | 240 |
| menthone | 30 | 5.11 | 2.22 | 26.6 | 150 | 327 | 14 | 236 |
| verbenone | 40 | 5.03 | 2.2 | 28.58 | 132 | 252 | 16 | 272 |
| thujone | 40 | 5.08 | 2.13 | 28.05 | 139 | 283 | 15 | 169 |
| camphor | 42 | 4.98 | 2.4 | 29.58 | 123 | 219 | 19 | 174 |

3.3. Repellency Bioassays

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ranum, P.; Peña-Rosas, J.P.; García-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 2014, 1312, 105–112. [Google Scholar] [CrossRef] [PubMed]
- INDEC. Anuario Estadístico de la República Argentina, 1st ed.; Itzcovich, N., Ed.; Instituto Nacional de Estadísticas y Censos (INDEC): Buenos Aires, Argentina, 2015; p. 366. (In Spanish) [Google Scholar]
- Oerke, E.C. Crop losses to pest. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Pitt, J.I.; Taniwaki, M.H.; Cole, M.B. Mycotoxin production in major crops as influenced by growing, harvesting, storage and processing, with emphasis on the achievement of food safety objectives. Food Control 2013, 32, 205–215. [Google Scholar] [CrossRef]
- Shephard, G.S. Mycotoxins worldwide: Current issues in Africa. In Meeting the Mycotoxin Menace; Wageningen Academic: Wageningen, The Netherlands, 2004; pp. 81–88. [Google Scholar]
- Theumer, M.G.; Canepa, M.C.; Lopez, A.G.; Mary, V.S.; Dambolena, J.S.; Rubinstein, H.R. Subchronic mycotoxicoses in Wistar rats: Assessment of the in vivo and in vitro genotoxicity induced by fumonisins and aflatoxin B1, and oxidative stress biomarkers status. Toxicology 2010, 268, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.Z.; Ji, R.; Li, Y.X.; Zhang, H.Y.; Li, B.; Zhao, Y. Occurrence of fumonisin B1 in corn from the main corn-producing areas of China. Mycopathologia 2009, 167, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Chulze, S.N. Strategies to reduce mycotoxin levels in maize during storage: A review. Food Addit. Contam. 2010, 27, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.P. Coleoptera. In Integrated Management of Insects in Stored Products; Subramanyam, B.H., Hagstrum, D.W., Eds.; Marcel Dekker: New York, NY, USA, 1996; pp. 1–40. [Google Scholar]
- Ferreira-Castro, F.L.; Potenza, M.R.; Rocha, L.O.; Correa, B. Interaction between toxigenic fungi and weevils in corn grain samples. Food Control 2012, 26, 594–600. [Google Scholar] [CrossRef]
- Bell, C.H. Fumigation in the 21st century. Crop Prot. 2000, 19, 563–569. [Google Scholar] [CrossRef]
- Brent, K.J.; Hollomon, D.W. Fungicide Resistance: The Assessment of Risk; FRAC Global Crop Protection Federation: Brussels, Belgium, 1998; Volume 2, pp. 1–48. [Google Scholar]
- Margni, M.; Rossier, D.; Crettaz, P.; Jolliet, O. Life cycle impact assessment of pesticides on human health and ecosystems. Agric. Ecosyst. Environ. 2002, 93, 379–392. [Google Scholar] [CrossRef]
- Dambolena, J.S.; López, A.G.; Meriles, J.M.; Rubinstein, H.; Zygadlo, J.A. Inhibitory effect of 10 natural phenolic compounds on Fusarium verticillioides. A structure-property-activity relationship study. Food Control 2012, 28, 163–170. [Google Scholar] [CrossRef]
- Pizzolitto, R.P.; Barberis, C.L.; Dambolena, J.S.; Herrera, J.M.; Zunino, M.P.; Magnoli, C.E.; Rubinstein, H.R.; Zygadlo, J.A.; Dalcero, A.M. Inhibitory effect of natural phenolic compounds on Aspergillus parasiticus growth. J. Chem. 2015, 2015. [Google Scholar] [CrossRef]
- Herrera, J.M.; Zunino, M.P.; Massuh, Y.; Pizzollito, R.P.; Dambolena, J.S.; Gañan, N.; Zygadlo, J.A. Fumigant toxicity from five essential oils rich in ketones against Sitophilus zeamais (Motschulsky). Agriscientia 2014, 31, 35–41. [Google Scholar]
- Herrera, J.M.; Zunino, M.P.; Pizzolitto, R.P.; Dambolena, J.S.; Zygadlo, J.A. Effect of fungal volatile organic compounds on a fungus and an insect that damage stored maize. J. Stored Prod. 2015, 62, 74–80. [Google Scholar] [CrossRef]
- Leslie, J.; Summerel, B.A. Species concepts in Fusarium. In The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006; Chapter 9; pp. 88–95. [Google Scholar]
- Pizzolitto, R.P.; Dambolena, J.S.; Zunino, M.P.; Larrauri, M.; Grosso, N.R.; Nepote, V.; Dalcero, A.M.; Zygadlo, J.A. Activity of natural compounds from peanut skins on Fusarium verticillioides growth and fumonisin B1 production. Ind. Crop. Prod. 2013, 47, 286–290. [Google Scholar] [CrossRef]
- Herrera, J.M.; Zunino, M.P.; Dambolena, J.S.; Pizzolitto, R.P.; Gañan, N.A.; Lucini, E.I.; Zygadlo, J.A. Terpene ketones as natural insecticides against Sitophilus zeamais. Ind. Crop. Prod. 2015, 70, 435–442. [Google Scholar] [CrossRef]
- Food and Agricultural Organization (FAO). Recommended methods for the detection and measurement of resistance of agricultural pests to pesticides. Tentative method for adults of some major beetle pests of stored cereals with malathion or lindane—FAO Method NO. 15. FAO Plant Protect. Bull. 1974, 22, 127–137. [Google Scholar]
- Germinara, G.S.; de Cristofaro, A.; Rotundo, G. Behavioral responses of adult Sitophilus granarius to individual cereal volatiles. J. Chem. Ecol. 2008, 34, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Neri, F.; Mari, M.; Brigati, S.; Bertolini, P. Fungicidal activity of plant volatile compounds for controlling Monilinia laxa in stone fruit. Plant Dis. 2007, 91, 30–35. [Google Scholar] [CrossRef]
- Di Rienzo, J.A.; Guzmán, A.W.; Casanoves, F. A multiple-comparisons method based on the distribution of the root node distance of a binary tree. J. Agric. Biol. Envirn. Stat. 2002, 7, 129–142. [Google Scholar] [CrossRef]
- Phillips, T.W.; Jiang, X.L.; Burkholder, W.E.; Phillips, J.K.; Tran, H.Q. Behavioral responses to food volatiles by two species of stored-product Coleoptera, Sitophilus oryzae (Curculionidae) and Tribolium castaneum (Tenebrionidae). J. Chem. Ecol. 1993, 19, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C. Infostat, versión 2010, Grupo Infostat, Facultad de Ciencias Agropecuarias (FCA), Universidad Nacional de Córdoba: Córdoba, Argentina, 2010.
- Dambolena, J.S.; Zunino, M.P.; López, A.G.; Rubinstein, H.R.; Zygadlo, J.A.; Mwangi, J.W.; Thoithi, G.N.; Kibwage, I.O.; Mwalukumbi, J.M.; Kariuki, S.T. Essential oils composition of Ocimum basilicum L. and Ocimum gratissimum L. from Kenya and their inhibitory effects on growth and fumonisin production by Fusarium verticillioides. Innov. Food Sci. Emerg. Technol. 2010, 11, 410–414. [Google Scholar] [CrossRef]
- Dambolena, J.S.; López, A.G.; Cánepa, M.C.; Theumer, M.G.; Zygadlo, J.A.; Rubinstein, H.R. Inhibitory effect of cyclic terpenes (limonene, menthol, menthone and thymol) on Fusarium verticillioides MRC 826 growth and fumonisin B1 biosynthesis. Toxicon 2008, 51, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Gallucci, M.N.; Carezzano, M.E.; Oliva, M.M.; Demo, M.S.; Pizzolitto, R.P.; Zunino, M.P.; Zygadlo, J.A.; Dambolena, J.S. In vitro activity of natural phenolic compounds against fluconazole-resistant Candida species: A quantitative structure-activity relationship analysis. J. Appl. Microbiol. 2014, 116, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Samapundo, S.; de Meulenaer, B.; Osei-Nimoh, D.; Lamboni, Y.; Debevere, J.; Devlieghere, F. Can phenolic compounds be used for the protection of corn from fungal invasion and mycotoxin contamination during storage? Food Microbiol. 2007, 24, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Putz, M.V. Quantum and electrodynamic versatility of electronegativity and chemical hardness. In Quantum Frontiers of Atoms and Molecules; Nova Science Publishers, Inc.: New York, NY, USA, 2011; pp. 251–275. [Google Scholar]
- Schwobel, J.A.H.; Koleva, Y.K.; Enoch, S.J.; Bajot, F.; Hewitt, M.; Madden, J.C.; Roberts, D.W.; Schultz, T.W.; Cronin, M.T.D. Measurement and Estimation of Electrophilic Reactivity for Predictive Toxicology. Chem. Rev. 2011, 111, 2562–2596. [Google Scholar] [CrossRef] [PubMed]
- Schultz, T.W.; Yarbrough, J.W. Trends in structure-toxicity relationships for carbonyl conatining α,β-unsatured compounds. SAR QSAR Environ. Res. 2004, 15, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Lopachin, R.M.; Gavin, T. Molecular Mechanisms of Aldehyde Toxicity: A Chemical Perspective. Chem. Res. Toxicol. 2014, 27, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Semchyshyn, H.M. Reactive carbonyl species in vivo: Generation and dual biological effects. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Escher, B.; Hermes, J.M. Modes of Action in Ecotoxicology: Their Role in Body Burdens, Species Sensitivity, QSARs, and Mixture Effects. Environ. Sci. Technol. 2002, 36, 4201–4217. [Google Scholar] [CrossRef] [PubMed]
- Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Rubinstein, H.R.; Zygadlo, J.A.; Dambolena, J.S. Effect of Selected Volatiles on Two Stored Pests: The Fungus Fusarium verticillioides and the Maize Weevil Sithophilus zeamais. J. Agric. Food Chem. 2015, 63, 7743–7749. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pizzolitto, R.P.; Herrera, J.M.; Zaio, Y.P.; Dambolena, J.S.; Zunino, M.P.; Gallucci, M.N.; Zygadlo, J.A. Bioactivities of Ketones Terpenes: Antifungal Effect on F. verticillioides and Repellents to Control Insect Fungal Vector, S. zeamais. Microorganisms 2015, 3, 851-865. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040851
Pizzolitto RP, Herrera JM, Zaio YP, Dambolena JS, Zunino MP, Gallucci MN, Zygadlo JA. Bioactivities of Ketones Terpenes: Antifungal Effect on F. verticillioides and Repellents to Control Insect Fungal Vector, S. zeamais. Microorganisms. 2015; 3(4):851-865. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040851
Chicago/Turabian StylePizzolitto, Romina P., Jimena M. Herrera, Yesica P. Zaio, Jose S. Dambolena, Maria P. Zunino, Mauro N. Gallucci, and Julio A. Zygadlo. 2015. "Bioactivities of Ketones Terpenes: Antifungal Effect on F. verticillioides and Repellents to Control Insect Fungal Vector, S. zeamais" Microorganisms 3, no. 4: 851-865. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms3040851