Failure of Staphylococcus aureus to Acquire Direct and Cross Tolerance after Habituation to Cinnamon Essential Oil


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Essential Oil
2.2. Target Bacterium
2.3. Determination of Minimum Inhibitory Concentration (MIC)
2.4. Induction of Bacterial Direct and Cross Tolerance
2.5. Statistical Analysis
3. Results and Discussion
3.1. CEO Exhibited Antimicrobial Activity against S. aureus
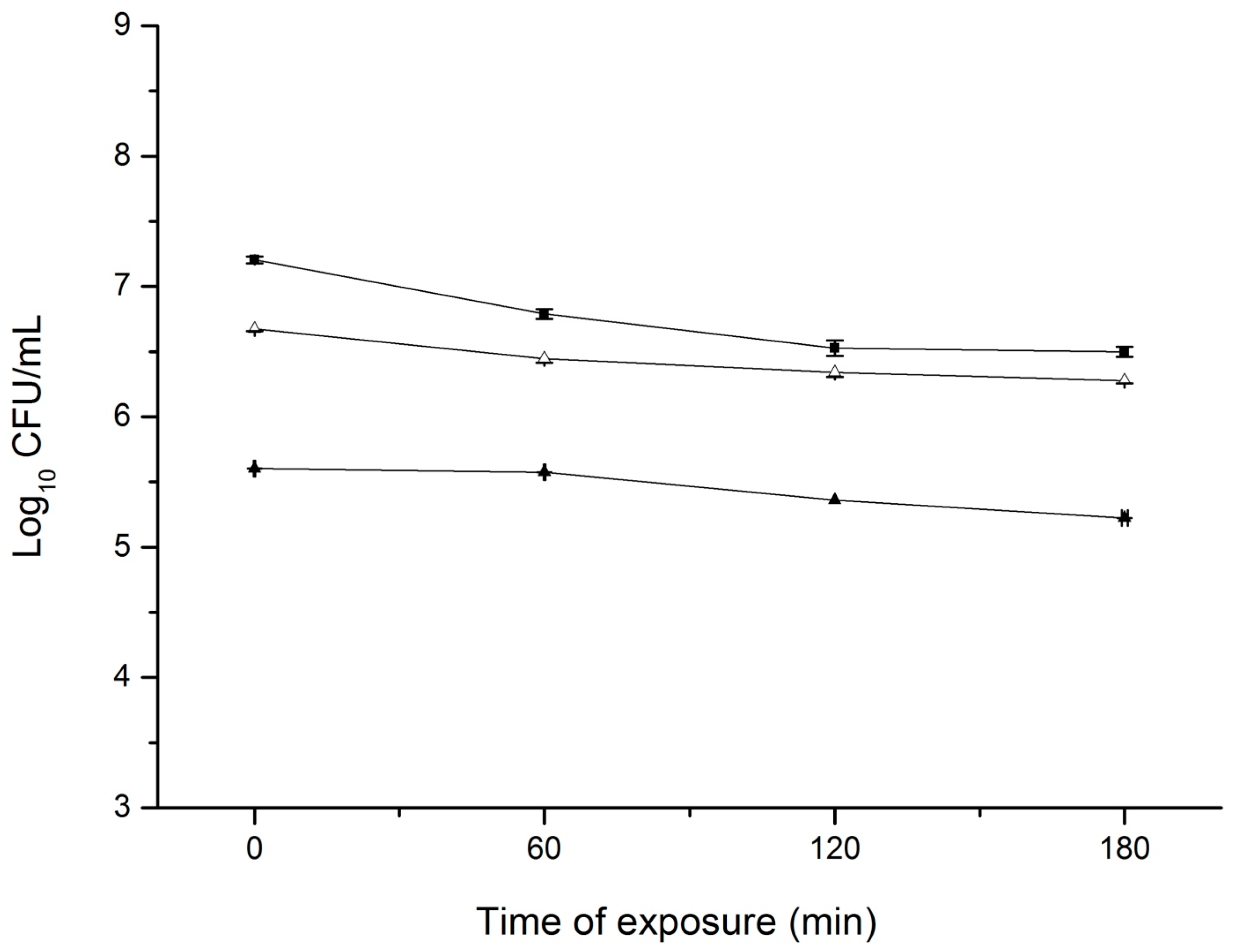
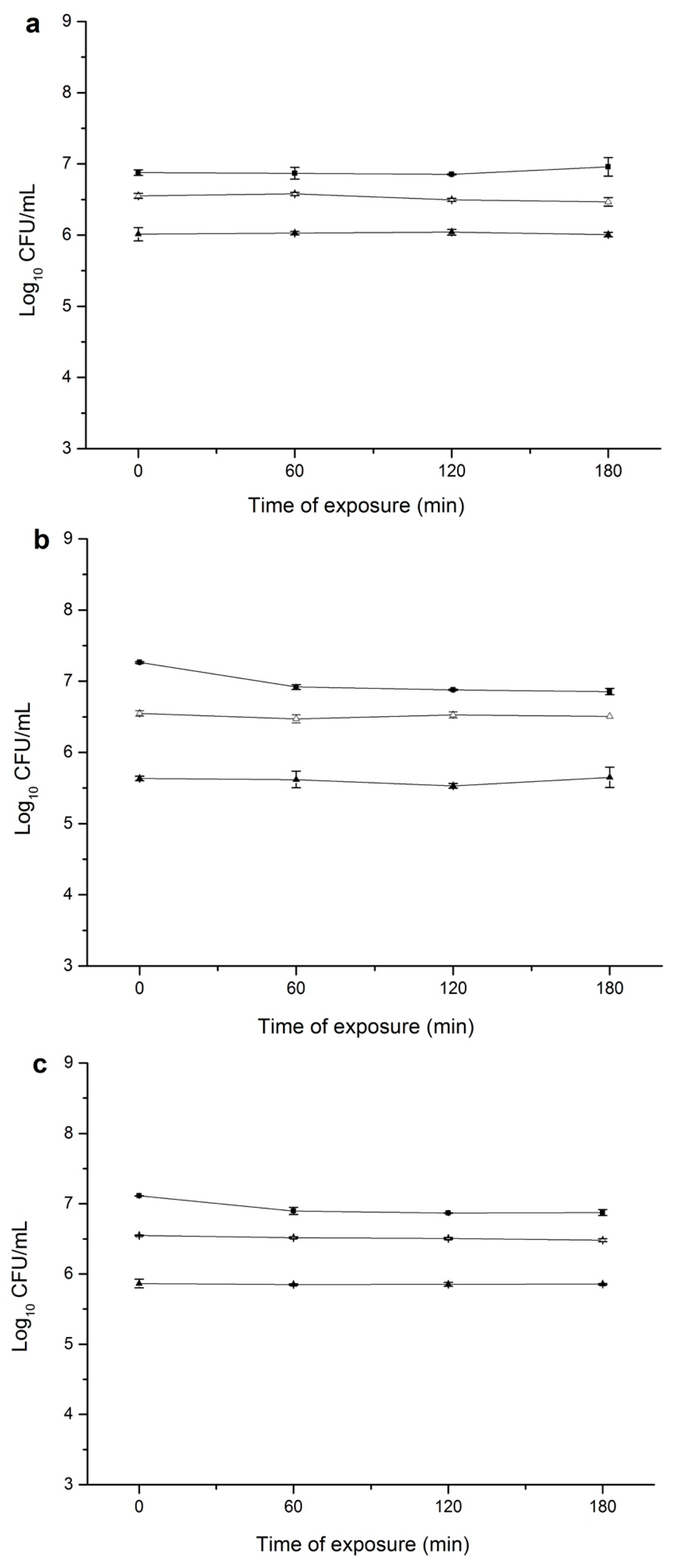
3.2. Overnight Exposure of S. aureus to CEO Did Not Induce Direct and Cross Tolerance
3.3. Repeated Exposure of S. aureus to Increasing Amounts of CEO Did Not Induce Direct Tolerance
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Viçosa, G.N.; Botta, C.; Ferrocino, I.; Bertolino, M.; Ventura, M.; Nero, L.A.; Cocolin, L. Staphylococcus aureus undergoes major transcriptional reorganization during growth with Enterococcus faecalis in milk. Food Microbiol. 2018, 73, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; Deleo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Bikels-Goshen, T.; Landau, E.; Saguy, S.; Shapira, R. Staphylococcal strains adapted to epigallocathechin gallate (EGCG) show reduced susceptibility to vancomycin, oxacillin and ampicillin, increased heat tolerance, and altered cell morphology. Int. J. Food Microbiol. 2010, 138, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Cebrián, G.; Sagarzazu, N.; Pagán, R.; Condón, S.; Mañas, P. Development of stress resistance in Staphylococcus aureus after exposure to sublethal environmental conditions. Int. J. Food Microbiol. 2010, 140, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Jayasena, D.D.; Jo, C. Essential oils as potential antimicrobial agents in meat and meat products: A review. Trends Food Sci. Technol. 2013, 34, 96–108. [Google Scholar] [CrossRef]
- Souza, E.L.; Almeida, E.T.; Guedes, J.P. The potential of the incorporation of essential oils and their individual constituents to improve microbial safety in juices: A review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 753–772. [Google Scholar] [CrossRef]
- He, S.; Wang, Y.; Sun, Y.; Chen, S.; Zhang, Y.; Ying, M. Antimicrobial activity and preliminary characterization of κ-carrageenan films containing cinnamon essential oil. Adv. J. Food Sci. Technol. 2015, 9, 523–528. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Wang, Y.; Jiang, P.; Quek, S. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus. Food Control 2016, 59, 282–289. [Google Scholar] [CrossRef]
- Cui, H.; Li, W.; Li, C.; Vittayapadung, S.; Lin, L. Liposome containing cinnamon oil with antibacterial activity against methicillin-resistant Staphylococcus aureus biofilm. Biofouling 2016, 32, 215–225. [Google Scholar] [CrossRef]
- He, S.; Yang, Q.; Ren, X.; Zi, J.; Lu, S.; Wang, S.; Zhang, Y.; Wang, Y. Antimicrobial efficiency of chitosan solutions and coatings incorporated with clove oil and/or ethylenediaminetetraacetate. J. Food Saf. 2014, 34, 345–352. [Google Scholar] [CrossRef]
- Wang, Y.; Xia, Y.; Zhang, P.; Ye, L.; Wu, L.; He, S. Physical characterization and pork packaging application of chitosan films incorporated with combined essential oils of cinnamon and ginger. Food Bioprocess Technol. 2017, 10, 503–511. [Google Scholar] [CrossRef]
- Souza, E.L. The effects of sublethal doses of essential oils and their constituents on antimicrobial susceptibility and antibiotic resistance among food-related bacteria: A review. Trends Food Sci. Technol. 2016, 56, 1–12. [Google Scholar] [CrossRef]
- He, S.; Zhou, X.; Shi, C.; Shi, X. Ethanol adaptation induces direct protection and cross-protection against freezing stress in Salmonella enterica serovar Enteritidis. J. Appl. Microbiol. 2016, 120, 697–704. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Ren, X.; Lu, Y.; Zhang, Y.; Wang, Y.; Sun, L. Microemulsification of clove essential oil improves its in vitro and in vivo control of Penicillium digitatum. Food Control 2016, 65, 106–111. [Google Scholar] [CrossRef]
- Gomes-Neto, N.J.; Luz, I.S.; Tavares, A.G.; Honorio, V.G.; Magnani, M.; Souza, E.L. Rosmarinus officinalis L. essential oil and its majority compound 1,8-cineole at sublethal amounts induce no direct and cross protection in S. aureus ATCC 6538. Foodborne Pathog. Dis. 2012, 9, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Neto, N.J.; Luz, I.S.; Honório, W.G.; Tavares, A.G.; Souza, E.L. Rosmarinus officinalis L. essential oil and the related compound 1, 8-cineole do not induce direct or cross-protection in Listeria monocytogenes ATCC 7644 cultivated in meat broth. Can. J. Microbiol. 2012, 58, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Neto, N.J.; da Silva Luz, I.; Honório, V.G.; Conceição, M.L.; Souza, E.L. Pseudomonas aeruginosa cells adapted to Rosmarinus officinalis L. essential oil and 1, 8-cineole acquire no direct and cross protection in a meat-based broth. Food Res. Int. 2012, 49, 143–146. [Google Scholar] [CrossRef]
- Gomes-Neto, N.J.; Luz, I.S.; Franco, O.L.; Magnani, M.; Souza, E.L. Tolerance evaluation in Salmonella enterica serovar Typhimurium challenged with sublethal amounts of Rosmarinus officinalis L. essential oil or 1, 8-cineole in meat model. Int. J. Food Sci. Technol. 2014, 49, 1912–1917. [Google Scholar] [CrossRef]
- Luz, I.S.; Neto, N.J.G.; Tavares, A.G.; Nunes, P.C.; Magnani, M.; Souza, E.L. Evidence for lack of acquisition of tolerance in Salmonella enterica serovar Typhimurium ATCC 14028 after exposure to subinhibitory amounts of Origanum vulgare L. essential oil and carvacrol. Appl. Environ. Microbiol. 2012, 78, 5021–5024. [Google Scholar] [CrossRef] [PubMed]
- Luz, I.D.S.; Neto, N.J.G.; Tavares, A.G.; Magnani, M.; Souza, E.L. Exposure of Listeria monocytogenes to sublethal amounts of Origanum vulgare L. essential oil or carvacrol in a food-based medium does not induce direct or cross protection. Food Res. Int. 2012, 48, 667–672. [Google Scholar] [CrossRef]
- Luz, I.S.; Neto, N.J.G.; Tavares, A.G.; Nunes, P.C.; Magnani, M.; de Souza, E.L. Lack of induction of direct protection or cross-protection in Staphylococcus aureus by sublethal concentrations of Origanum vulgare L. essential oil and carvacrol in a meat-based medium. Arch. Microbiol. 2013, 195, 587–593. [Google Scholar]
- Luz, I.S.; Gomes-Neto, N.J.; Magnani, M.; de Souza, E.L. Assessment of tolerance induction by Origanum vulgare L. essential oil or carvacrol in Pseudomonas aeruginosa cultivated in a meat-based broth and in a meat model. Food Sci. Technol. Int. 2014, 21, 571–580. [Google Scholar]
- Tavares, A.G.; Monte, D.F.M.D.; Albuquerque, A.D.R.; Sampaio, F.C.; Magnani, M.; Siqueira Júnior, J.P.D.; Souza, E.L. Habituation of enterotoxigenic Staphylococcus aureus to Origanum vulgare L. essential oil does not induce direct-tolerance and cross-tolerance to salts and organic acids. Braz. J. Microbiol. 2015, 46, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F. Effect of External Factors on Antimicrobial Activity of Cinnamon Oil. Master’s Thesis, Shanghai Institute of Technology, Shanghai, China, 25 May 2016. [Google Scholar]
- Wang, Y.; Bian, W.; Ren, X.; Song, X.; He, S. Microencapsulation of clove essential oil improves its antifungal activity against Penicillium digitatum in vitro and green mould on Navel oranges. J. Hortic. Sci. Biotechnol. 2018, 93, 159–166. [Google Scholar] [CrossRef]
- Begley, M.; Hill, C. Stress adaptation in foodborne pathogens. Annu. Rev. Food Sci. Technol. 2015, 6, 191–210. [Google Scholar] [CrossRef] [PubMed]
- To, M.S.; Favrin, S.; Romanova, N.; Griffiths, M.W. Postadaptational resistance to benzalkonium chloride and subsequent physicochemical modifications of Listeria monocytogenes. Appl. Environ. Microbiol. 2002, 68, 5258–5264. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods-a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Lu, F.; Ding, Y.C.; Ye, X.Q.; Ding, Y.T. Antibacterial effect of cinnamon oil combined with thyme or clove oil. Agr. Sci. China 2011, 10, 1482–1487. [Google Scholar] [CrossRef]
- Herman, A.; Bochenek, J.; Herman, A.P. Effect of cinnamon and lavender oils on FtsZ gene expression in the Staphylococcus aureus ATCC 29213. Appl. Biochem. Microbiol. 2013, 49, 481–484. [Google Scholar] [CrossRef]
- Bui, L.M.G. Staphylococcus aureus: Stress Response and Its Roles in Pathogenesis. Ph.D. Thesis, The University of Adelaide, Adelaide, South Australia, Australia, July 2015. [Google Scholar]
- Azizkhani, M.; Parsaeimehr, M. Effects of Cinnamomum zeylanicum and Ocimum basilicum essential oils on the growth of Staphylococcus aureus ATCC 29213 and gene expression of enterotoxins A, C and E. J. Essent. Oil Res. 2015, 27, 506–513. [Google Scholar] [CrossRef]
- He, S.; Cui, Y.; Qin, X.; Zhang, F.; Shi, C.; Paoli, G.C.; Shi, X. Influence of ethanol adaptation on Salmonella enterica serovar Enteritidis survival in acidic environments and expression of acid tolerance-related genes. Food Microbiol. 2018, 72, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Sihto, H.M. Stress response in Staphylococcus aureus: Regulatory Mechanisms Influencing Enterotoxin Gene Expression under Stress Relevant to Food Production and Preservation. Ph.D. Thesis, University of Bern, Bern, Swiss, 2016. [Google Scholar]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Effects of Melaleuca alternifolia (Tea tree) essential oil and the major monoterpene component terpinen-4-ol on the development of single-and multistep antibiotic resistance and antimicrobial susceptibility. Antimicrob. Agents Chemother. 2012, 56, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Haft, R.J.; Keating, D.H.; Schwaegler, T.; Schwalbach, M.S.; Vinokur, J.; Tremaine, M.; Peters, J.M.; Kotlajich, M.V.; Pohlmann, E.L.; Ong, I.M.; et al. Correcting direct effects of ethanol on translation and transcription machinery confers ethanol tolerance in bacteria. Proc. Natl. Acad. Sci. USA 2014, 111, E2576–E2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinouchi, T.; Tamaoka, K.; Furusawa, C.; Ono, N.; Suzuki, S.; Hirasawa, T.; Yomo, T.; Shimizu, H. Transcriptome analysis of parallel-evolved Escherichia coli strains under ethanol stress. BMC Genom. 2010, 11, 579. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, N.A.; Hammer, K.A.; Riley, T.V.; Van Belkum, A.; Carson, C.F. Effect of habituation to tea tree (Melaleuca alternifolia) oil on the subsequent susceptibility of Staphylococcus spp. to antimicrobials, triclosan, tea tree oil, terpinen-4-ol and carvacrol. Int. J. Antimicrob. Agents 2013, 41, 343–351. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Sun, Y.; Zhang, Q.; Yang, X.; Zheng, F.; He, S.; Wang, Y. Failure of Staphylococcus aureus to Acquire Direct and Cross Tolerance after Habituation to Cinnamon Essential Oil. Microorganisms 2019, 7, 18. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010018
Song X, Sun Y, Zhang Q, Yang X, Zheng F, He S, Wang Y. Failure of Staphylococcus aureus to Acquire Direct and Cross Tolerance after Habituation to Cinnamon Essential Oil. Microorganisms. 2019; 7(1):18. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010018
Chicago/Turabian StyleSong, Xiaoqiu, Yue Sun, Qian Zhang, Xiaobo Yang, Feng Zheng, Shoukui He, and Yifei Wang. 2019. "Failure of Staphylococcus aureus to Acquire Direct and Cross Tolerance after Habituation to Cinnamon Essential Oil" Microorganisms 7, no. 1: 18. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010018