Effect of Myclobutanil Pesticide on the Physiological Behavior of Two Newly Isolated Saccharomyces cerevisiae Strains during Very-High-Gravity Alcoholic Fermentation

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
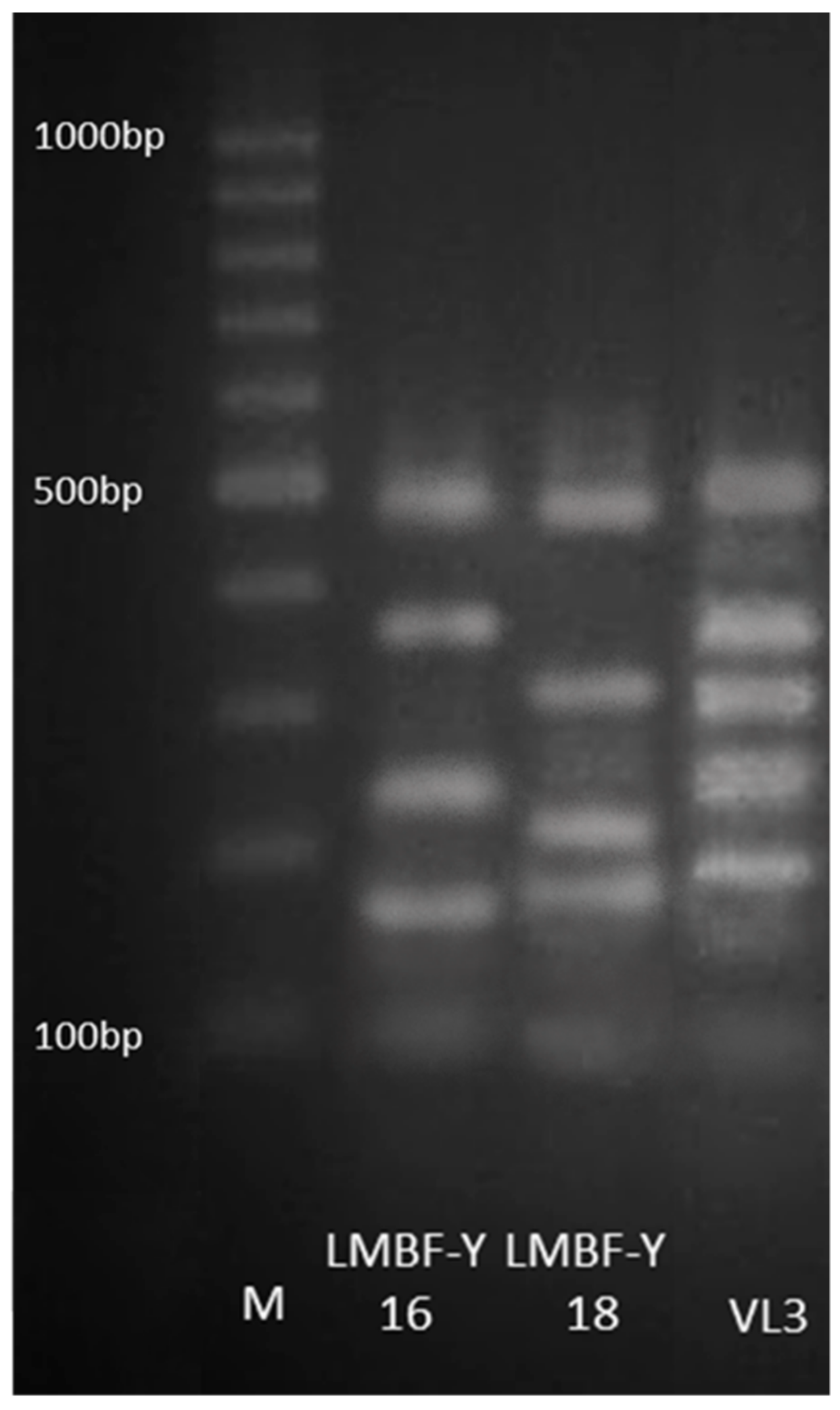
2.1. Yeast Isolation and Molecular Identification
2.2. Yeast Strains and Culture Conditions
2.3. Analytical Methods
2.4. Repeatability
3. Results and Discussion
3.1. Yeast Molecular Identification
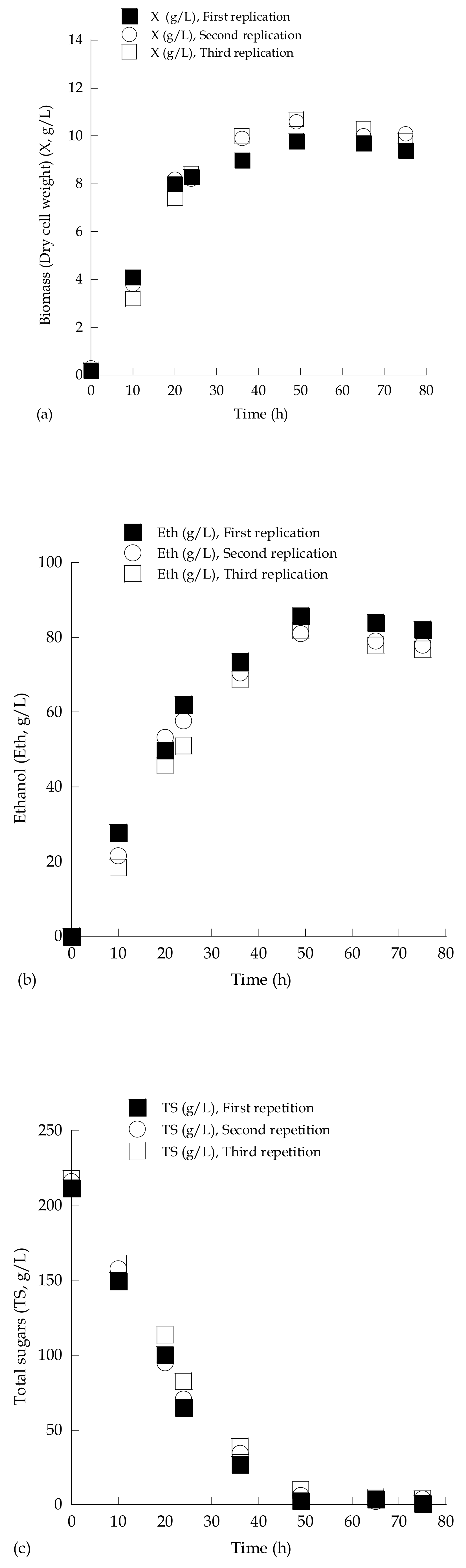
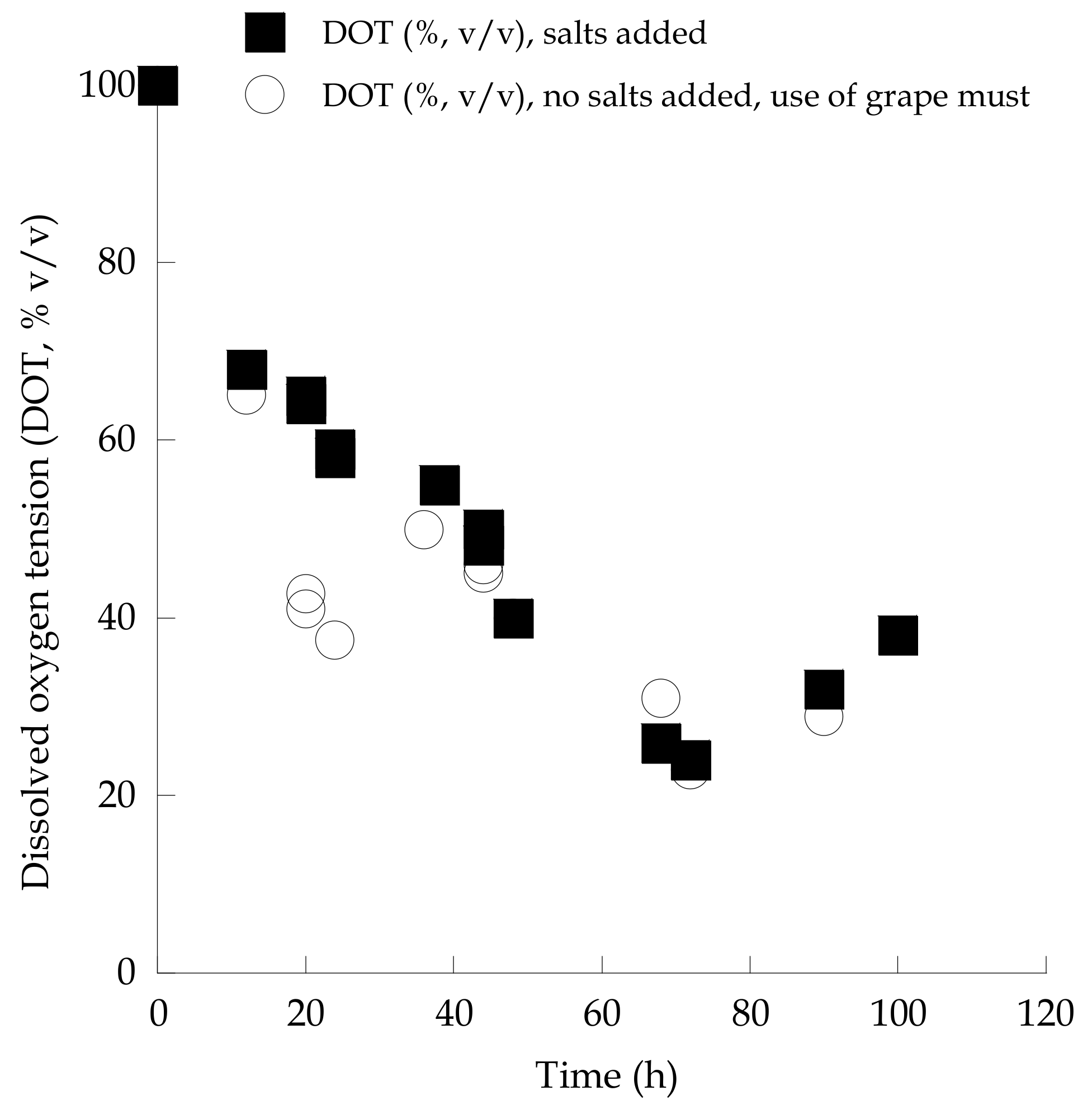
3.2. Medium and Shake-Flask Culture Optimization
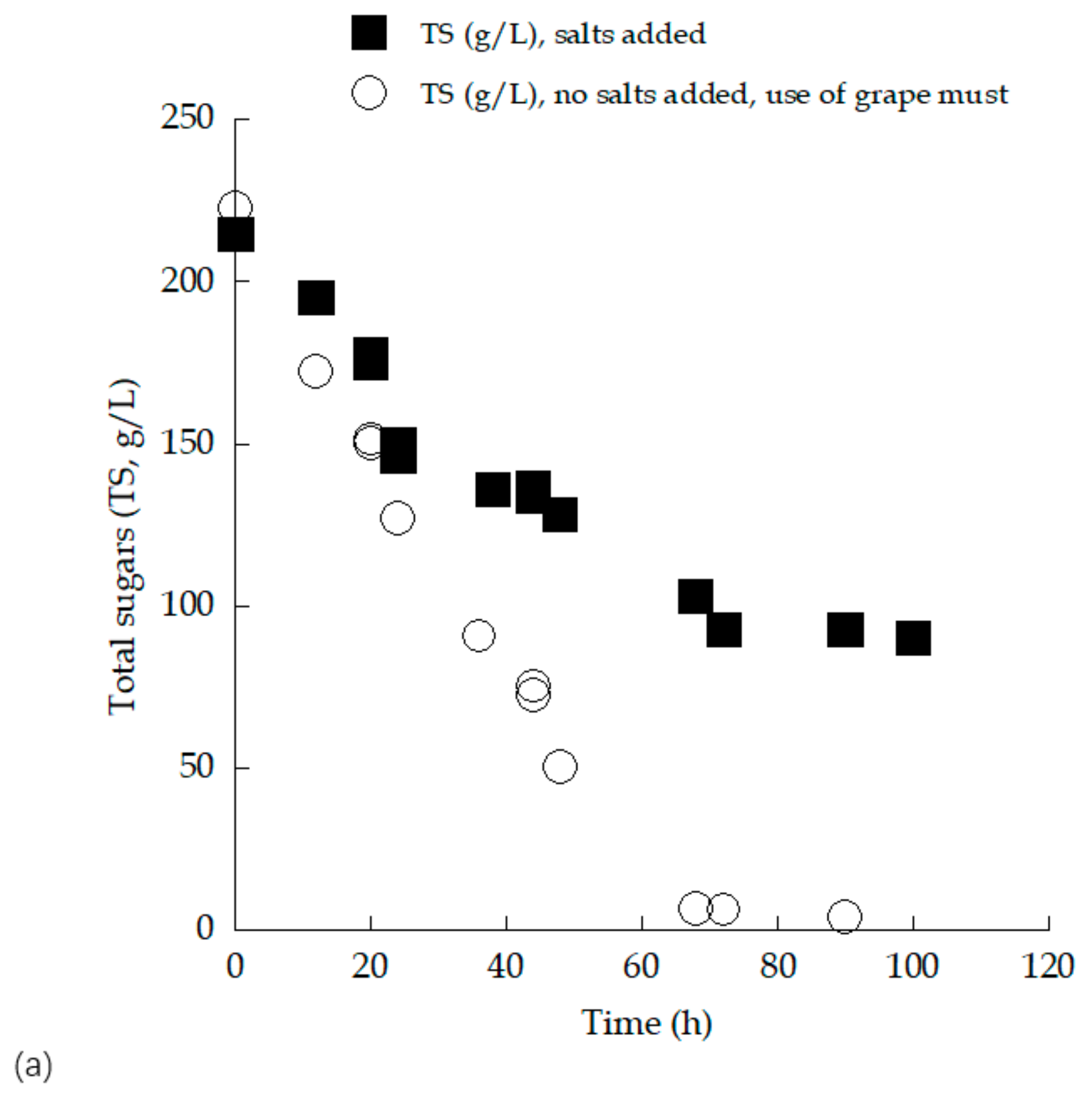
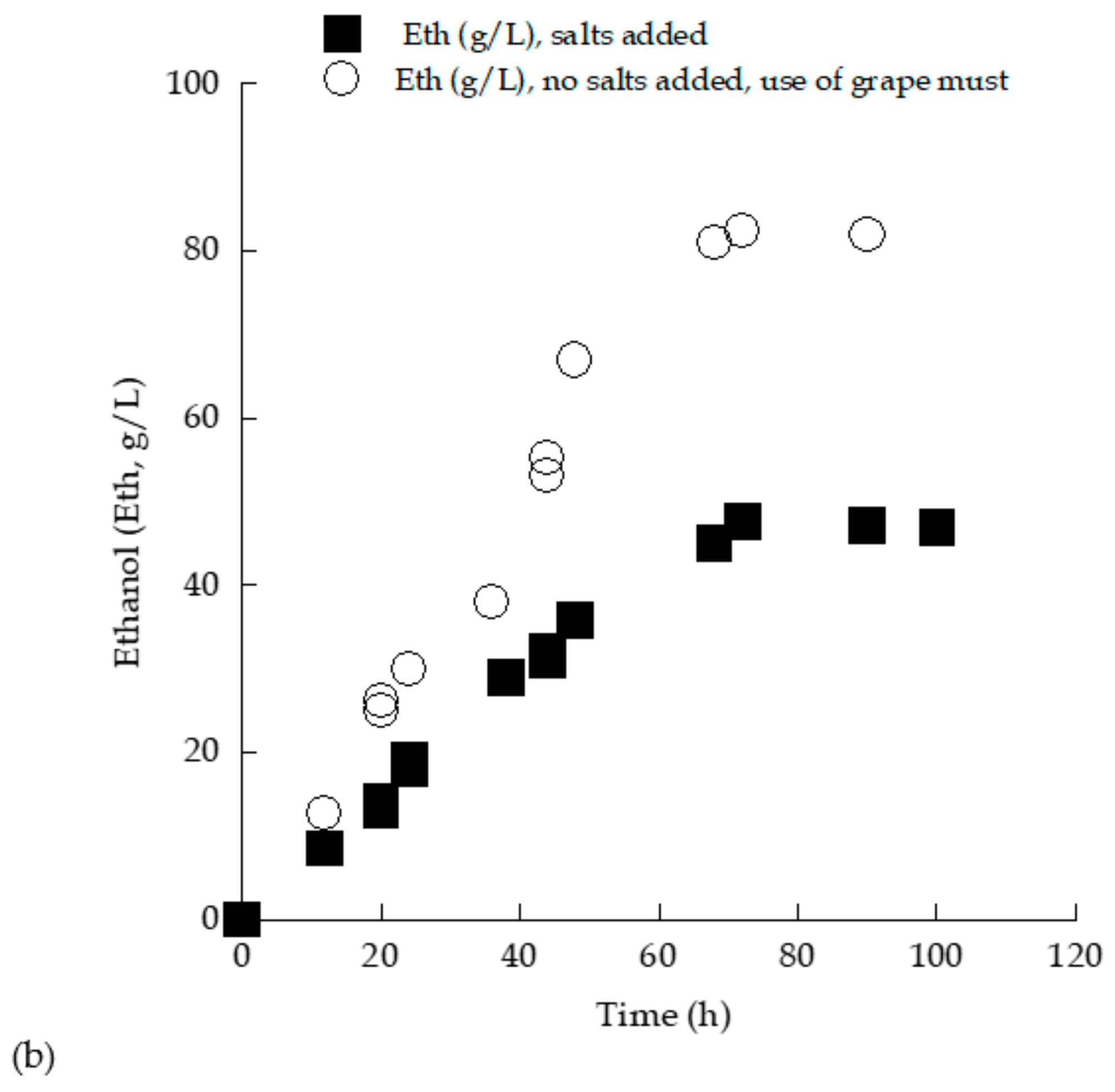
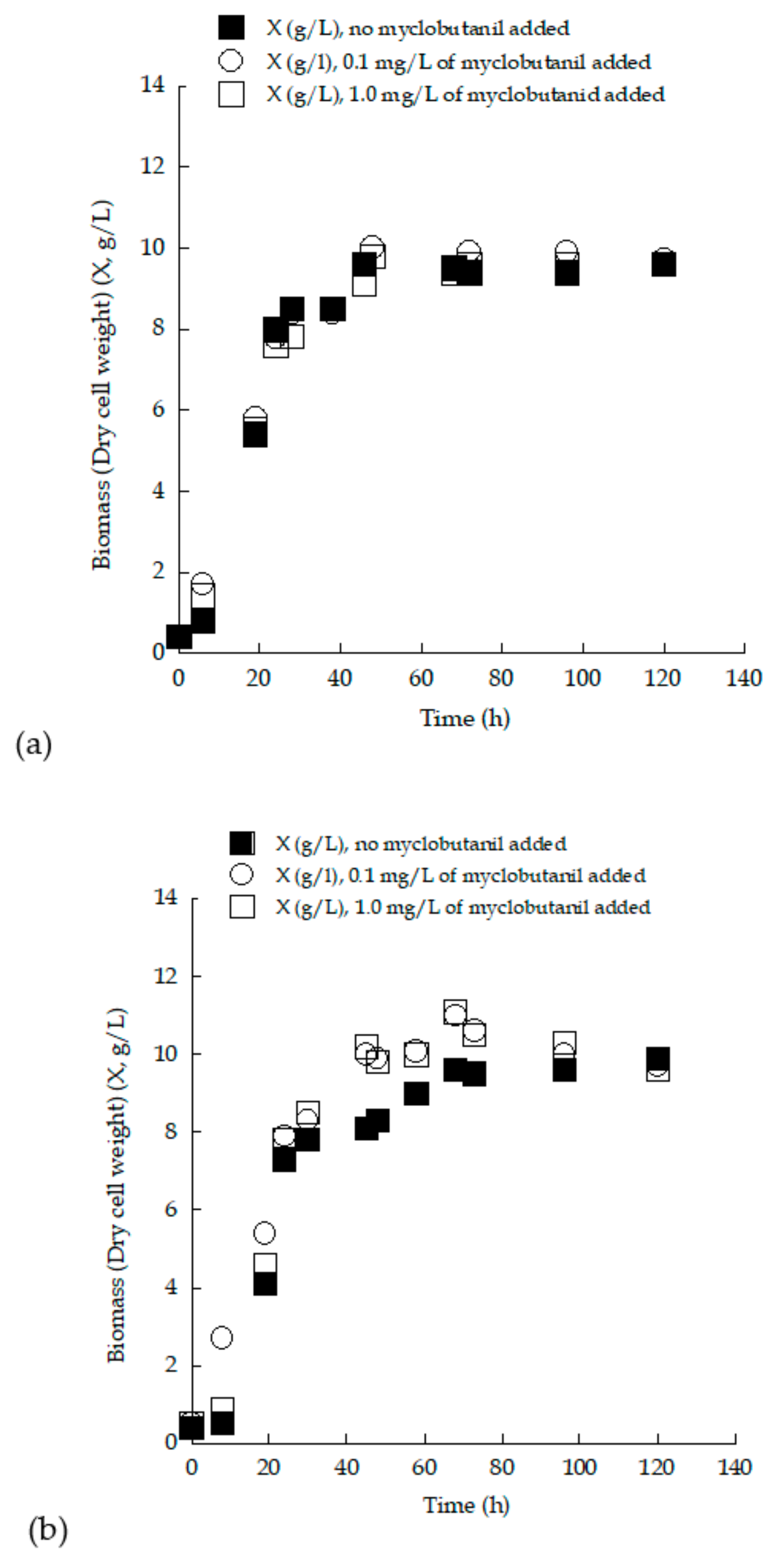
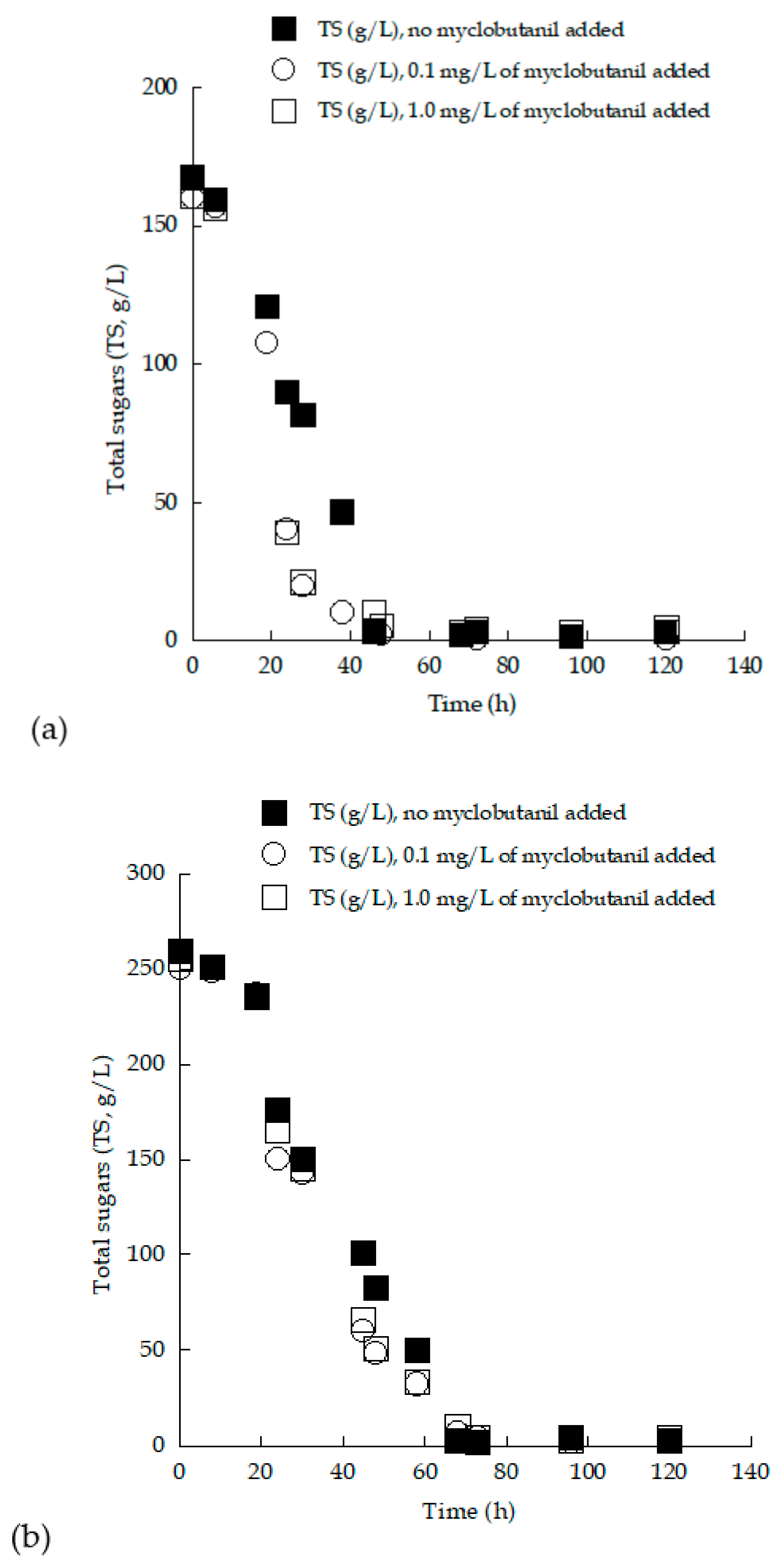
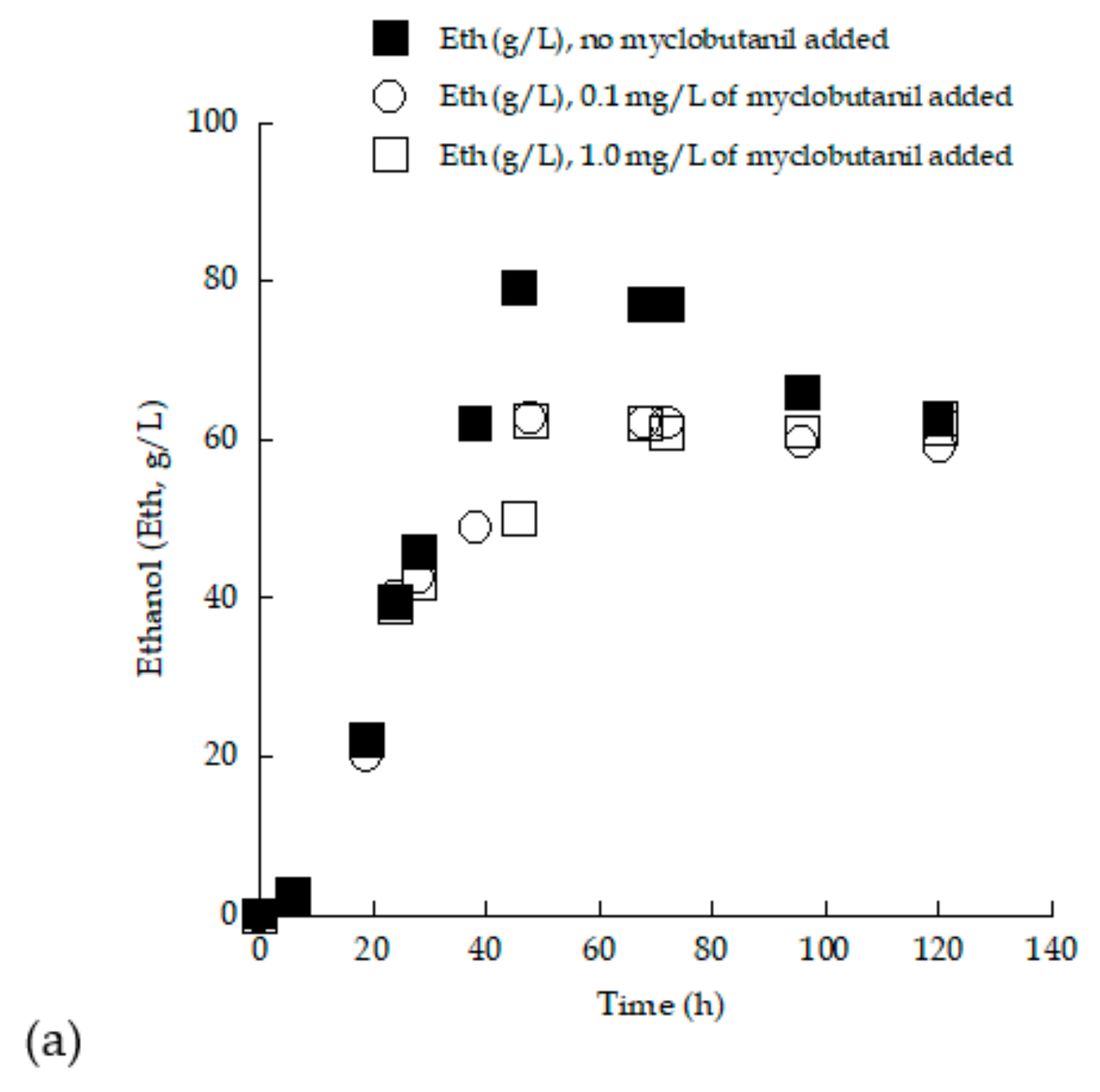
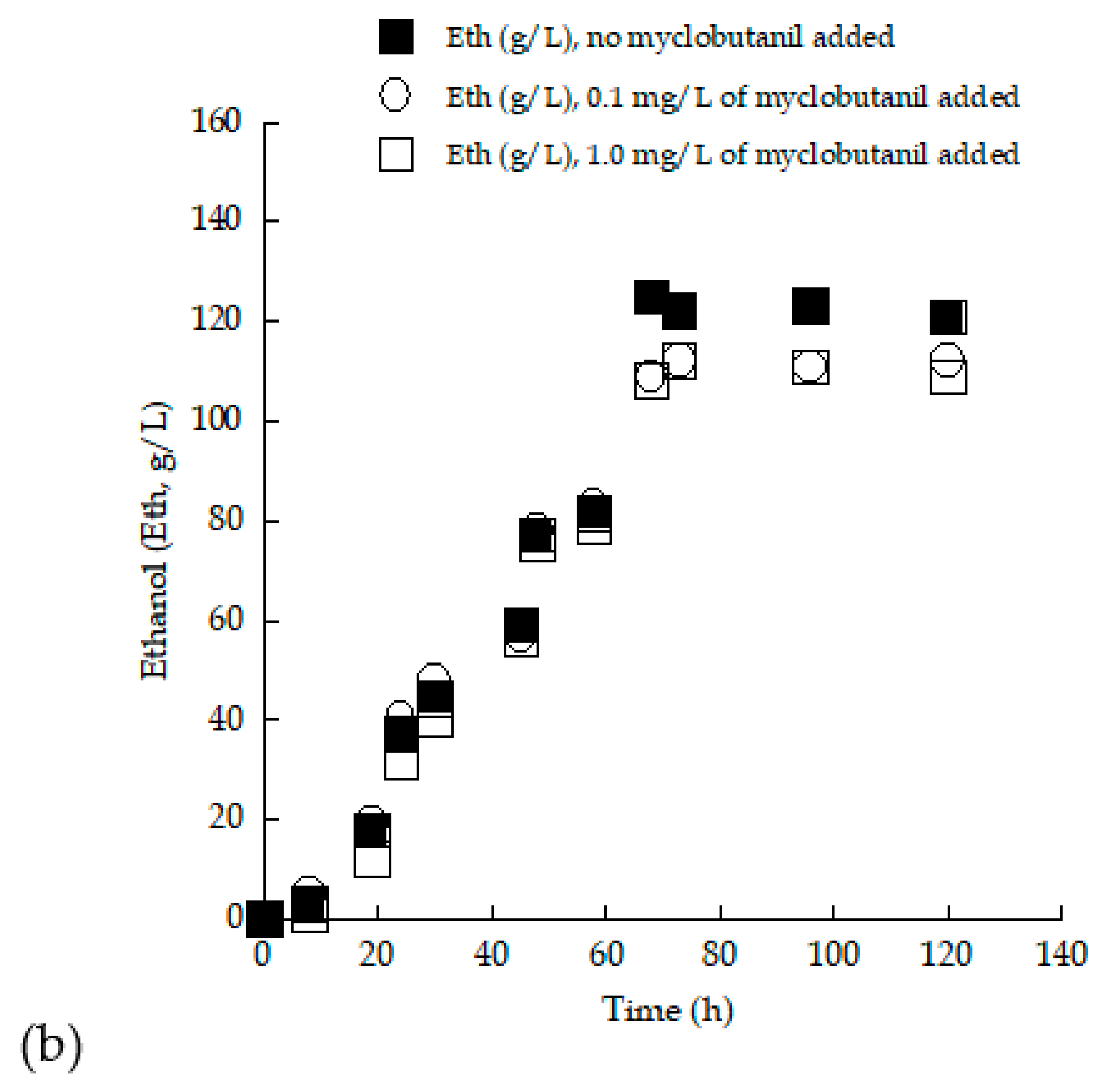
3.3. Trials in 2.0 L Flasks with (Enriched) Grape Must Employed as Substrate—Effect of Myclobutanil Addition on Cell Growth
3.4. Removal of Myclobutanil Pesticide
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Straathof, A.J.J.; Wahl, S.A.; Benjamin, K.R.; Takors, R.; Wierckx, N.; Noorman, H.J. Grand research challenges for sustainable industrial biotechnology. Trends Biotechnol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Camarasa, C.; Chiron, H.; Daboussi, F.; Della Valle, G.; Dumas, C.; Farines, V.; Floury, J.; Gagnaire, V.; Gorret, N.; Leonil, J.; et al. INRA’s research in industrial biotechnology: For food, chemicals, materials and fuels. Innov. Food Sci. Emerg. Technol. 2018, 46, 140–152. [Google Scholar] [CrossRef]
- Mohd Azhar, S.H.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Mohd Faik, A.A.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Koutinas, A.A.; Wang, R.H.; Webb, C. The biochemurgist -Bioconversion of agricultural raw materials for chemical production. Biofuels Bioprod. Biorefining 2007, 1, 24–38. [Google Scholar] [CrossRef]
- Sarris, D.; Papanikolaou, S. Biotechnological production of ethanol: Biochemistry, processes and technologies. Eng. Life Sci. 2016, 16, 307–329. [Google Scholar] [CrossRef] [Green Version]
- Sonderegger, M.; Schümperli, M.; Sauer, U. Metabolic engineering of a phosphoketolase pathway for pentose catabolism in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2004, 70, 2892–2897. [Google Scholar] [CrossRef] [Green Version]
- Widiastuti, H.; Lee, D.-Y.; Karimi, I.A. In silico analysis to explore the effect of various carbon sources on ethanol production in Zymomonas mobilis. In Computer Aided Chemical Engineering; Bogle, I.D.L., Fairweather, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 30, pp. 1382–1386. [Google Scholar]
- Koutinas, A.A.; Vlysidis, A.; Pleissner, D.; Kopsahelis, N.; Lopez Garcia, I.; Kookos, I.K.; Papanikolaou, S.; Kwan, T.H.; Lin, C.S.K. Valorization of industrial waste and by-product streams via fermentation for the production of chemicals and biopolymers. Chem. Soc. Rev. 2014, 43, 2587–2627. [Google Scholar] [CrossRef]
- Selim, K.A.; El-Ghwas, D.E.; Easa, S.M.; Abdelwahab Hassan, M.I. Bioethanol a microbial biofuel metabolite; New insights of yeasts Metabolic Engineering. Fermentation 2018, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Tanaka, S. Ethanol fermentation from biomass resources: Current state and prospects. Appl. Microbiol. Biotechnol. 2006, 69, 627–642. [Google Scholar] [CrossRef]
- Puligundla, P.; Smogrovicova, D.; Mok, C.; Obulam, V.S.R. A review of recent advances in high gravity ethanol fermentation. Renew. Energy 2019, 133, 1366–1379. [Google Scholar] [CrossRef]
- Phukoetphim, N.; Chan-u-tit, P.; Laopaiboon, P.; Laopaiboon, L. Improvement of bioethanol production from sweet sorghum juice under very high gravity fermentation: Effect of nitrogen, osmoprotectant, and aeration. Energies 2019, 12, 3620. [Google Scholar] [CrossRef] [Green Version]
- Puligundla, P.; Poludasu, R.M.; Rai, J.K.; Reddy Obulam, V.S. Repeated batch ethanolic fermentation of very high gravity medium by immobilized Saccharomyces cerevisiae. Ann. Microbiol. 2011, 61, 863–869. [Google Scholar] [CrossRef]
- Johnson, E.A.; Echavarri-Erasun, C. Chapter 3—Yeast Biotechnology. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 321–344. [Google Scholar] [CrossRef]
- Kunkee, R.E. Selection and modification of yeasts and lactic acid bacteria for wine fermentation. Food Microbiol. 1984, 1, 315–332. [Google Scholar] [CrossRef]
- Le Jeune, C.; Erny, C.; Demuyter, C.; Lollier, M. Evolution of the population of Saccharomyces cerevisiae from grape to wine in a spontaneous fermentation. Food Microbiol. 2006, 23, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Lepe, J.A.; Morata, A. New trends in yeast selection for winemaking. Trends Food Sci. Technol. 2012, 23, 39–50. [Google Scholar] [CrossRef]
- Querol, A.; Pérez-Torrado, R.; Alonso-del-Real, J.; Minebois, R.; Stribny, J.; Oliveira, B.M.; Barrio, E. Chapter Five—New Trends in the uses of yeasts in Oenology. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 85, pp. 177–210. [Google Scholar]
- Ganatsios, V.; Terpou, A.; Gialleli, A.-I.; Kanellaki, M.; Bekatorou, A.; Koutinas, A.A. A ready-to-use freeze-dried juice and immobilized yeast mixture for low temperature sour cherry (Prunus cerasus) wine making. Food Bioprod. Process. 2019, 117, 373–379. [Google Scholar] [CrossRef]
- Cappello, M.S.; Poltronieri, P.; Blaiotta, G.; Zacheo, G. Molecular and physiological characteristics of a grape yeast strain containing atypical genetic material. Int. J. Food Microbiol. 2010, 144, 72–80. [Google Scholar] [CrossRef]
- Rai, A.K.; Pandey, A.; Sahoo, D. Biotechnological potential of yeasts in functional food industry. Trends Food Sci. Technol. 2019, 83, 129–137. [Google Scholar] [CrossRef]
- Rousseaux, S.; Diguta, C.F.; Radoï-Matei, F.; Alexandre, H.; Guilloux-Bénatier, M. Non-Botrytis grape-rotting fungi responsible for earthy and moldy off-flavors and mycotoxins. Food Microbiol. 2014, 38, 104–121. [Google Scholar] [CrossRef]
- IARC. Monographs on the Evaluation of carcinogenic risk to humans. In Toxins Derived From Fusarium moniliforme: Fumonisins B1 and B2 and Fusarin C; International Agency of Research on Cancer: Lion, France, 1993; pp. 445–466. [Google Scholar]
- Marín, S.; Cano-Sancho, G.; Sanchis, V.; Ramos, A.J. The role of mycotoxins in the human exposome: Application of mycotoxin biomarkers in exposome-health studies. Food Chem. Toxicol. 2018, 121, 504–518. [Google Scholar] [CrossRef] [Green Version]
- Dzedze, N.; Van Breda, V.; Hart, R.S.; Van Wyk, J. Wine chemical, sensory, aroma compound and protein analysis of wines produced from chemical and biological fungicide treated Chenin blanc grapes. Food Control 2019, 105, 265–276. [Google Scholar] [CrossRef]
- Ben Taheur, F.; Kouidhi, B.; Al Qurashi, Y.M.A.; Ben Salah-Abbès, J.; Chaieb, K. Review: Biotechnology of mycotoxins detoxification using microorganisms and enzymes. Toxicon 2019, 160, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Briz-Cid, N.; Castro-Sobrino, L.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Fungicide residues affect the sensory properties and flavonoid composition of red wine. J. Food Compos. Anal. 2018, 66, 185–192. [Google Scholar] [CrossRef]
- Noguerol-Pato, R.; Sieiro-Sampedro, T.; González-Barreiro, C.; Cancho-Grande, B.; Simal-Gándara, J. Evaluation of the effect of fenhexamid and mepanipyrim in the volatile composition of Tempranillo and Graciano wines. Food Res. Int. 2015, 71, 108–117. [Google Scholar] [CrossRef]
- Noguerol-Pato, R.; Torrado-Agrasar, A.; González-Barreiro, C.; Cancho-Grande, B.; Simal-Gándara, J. Influence of new generation fungicides on Saccharomyces cerevisiae growth, grape must fermentation and aroma biosynthesis. Food Chem. 2014, 146, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Sarris, D.; Kotseridis, Y.; Linga, M.; Galiotou-Panayotou, M.; Papanikolaou, S. Enhanced ethanol production, volatile compound biosynthesis and fungicide removal during growth of a newly isolated Saccharomyces cerevisiae strain on enriched pasteurized grape musts. Eng. Life Sci. 2009, 9, 29–37. [Google Scholar] [CrossRef]
- Sieiro-Sampedro, T.; Figueiredo-González, M.; González-Barreiro, C.; Simal-Gandara, J.; Cancho-Grande, B.; Rial-Otero, R. Impact of mepanipyrim and tetraconazole in Mencía wines on the biosynthesis of volatile compounds during the winemaking process. Food Chem. 2019, 300, 125223. [Google Scholar] [CrossRef]
- Athanasopoulos, P.E.; Pappas, C.J.; Kyriakidis, N.V. Decomposition of myclobutanil and triadimefon in grapes on the vines and during refrigerated storage. Food Chem. 2003, 82, 367–371. [Google Scholar] [CrossRef]
- Fonseca, F.S.; Carrão, D.B.; de Albuquerque, N.C.P.; Nardini, V.; Dias, L.G.; da Silva, R.M.; Lopes, N.P.; de Oliveira, A.R.M. Myclobutanil enantioselective risk assessment in humans through in vitro CYP450 reactions: Metabolism and inhibition studies. Food Chem. Toxicol. 2019, 128, 202–211. [Google Scholar] [CrossRef]
- Ling, C.; Liew, Z.; Von Ehrenstein, O.S.; Heck, J.E.; Park, A.S.; Cui, X.; Cockburn, M.; Wu, J.; Ritz, B. Prenatal exposure to ambient pesticides and preterm birth and term low birthweight in agricultural regions of California. Toxics 2018, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Tully, D.B.; Bao, W.; Goetz, A.K.; Blystone, C.R.; Ren, H.; Schmid, J.E.; Strader, L.F.; Wood, C.R.; Best, D.S.; Narotsky, M.G.; et al. Gene expression profiling in liver and testis of rats to characterize the toxicity of triazole fungicides. Toxicol. Appl. Pharmacol. 2006, 215, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Goetz, A.K.; Dix, D.J. Mode of action for reproductive and hepatic toxicity inferred from a genomic study of triazole antifungals. Toxicol. Sci. 2009, 110, 449–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetz, A.K.; Dix, D.J. Toxicogenomic effects common to triazole antifungals and conserved between rats and humans. Toxicol. Appl. Pharmacol. 2009, 238, 80–89. [Google Scholar] [CrossRef]
- Hao, W.; Zhang, Y.; Xie, Y.; Guo, B.; Chang, J.; Li, J.; Xu, P.; Wang, H. Myclobutanil accumulation, transcriptional alteration, and tissue injury in lizards (Eremias argus) treated with myclobutanil enantiomers. Ecotoxicol. Environ. Saf. 2019, 171, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.M.; Walker, R.S.K. Enhancing yeast alcoholic fermentations. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Ramón, D. A Comparative study of different methods of yeast strain characterization. Syst. Appl. Microbiol. 1992, 15, 439–446. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Weiss, S.; Samson, F.; Navarro, D.; Casaregola, S. Yeast IP: A database for identification and phylogeny of Saccharomycotina yeasts. FEMS Yeast Res. 2013, 13, 117–125. Available online: https://0-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/pdf/10.1111/1567-1364.12017 (accessed on 4 May 2019). [CrossRef] [Green Version]
- Legras, J.-L.; Karst, F. Optimisation of interdelta analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Legras, J.L.; Merdinoglu, D.; Cornuet, J.M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Aggelis, G.; Marc, I. Kinetic profile of the cellular lipid composition in an oleaginous Yarrowia lipolytica capable of producing a cocoa-butter substitute from industrial fats. Antonie Van Leeuwenhoek 2001, 80, 215–224. [Google Scholar] [CrossRef]
- Sarris, D.; Matsakas, L.; Aggelis, G.; Koutinas, A.A.; Papanikolaou, S. Aerated vs non-aerated conversions of molasses and olive mill wastewaters blends into bioethanol by Saccharomyces cerevisiae under non-aseptic conditions. Ind. Crop. Prod. 2014, 56, 83–93. [Google Scholar] [CrossRef]
- Miller, G.L. Use of DNS reagent for the determination of glucose. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Sarris, D.; Giannakis, M.; Philippoussis, A.; Komaitis, M.; Koutinas, A.A.; Papanikolaou, S. Conversions of olive mill wastewater-based media by Saccharomyces cerevisiae through sterile and non-sterile bioprocesses. J. Chem. Technol. Biotechnol. 2013, 88, 958–969. [Google Scholar] [CrossRef]
- Palaiogeorgou, A.M.; Papanikolaou, S.; de Castro, A.M.; Freire, D.M.G.; Kookos, I.K.; Koutinas, A.A. A newly isolated Enterobacter sp. strain produces 2,3-butanediol during its cultivation on low-cost carbohydrate-based substrates. FEMS Microbiol. Lett. 2019, 366, fny280. [Google Scholar] [CrossRef] [PubMed]
- Hrouzková, S. Analytical Methods for Pesticide Detection in Foodstuffs; Wiley: Hoboken, NJ, UAS, 2016. [Google Scholar] [CrossRef]
- Bakırcı, G.T.; Yaman Acay, D.B.; Bakırcı, F.; Ötleş, S. Pesticide residues in fruits and vegetables from the Aegean region, Turkey. Food Chem. 2014, 160, 379–392. [Google Scholar] [CrossRef]
- Doulia, D.S.; Anagnos, E.K.; Liapis, K.S.; Klimentzos, D.A. Effect of clarification process on the removal of pesticide residues in white wine. Food Control 2017, 72, 134–144. [Google Scholar] [CrossRef]
- Botezatu, A.; Pickering, G.J.; Kotseridis, Y. Development of a rapid method for the quantitative analysis of four methoxypyrazines in white and red wine using multi-dimensional Gas Chromatography—Mass Spectrometry. Food Chem. 2014, 160, 141–147. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Rontou, M.; Belka, A.; Athenaki, M.; Gardeli, C.; Mallouchos, A.; Kalantzi, O.; Koutinas, A.A.; Kookos, I.K.; Zeng, A.P.; et al. Conversion of biodiesel-derived glycerol into biotechnological products of industrial significance by yeast and fungal strains. Eng. Life Sci. 2017, 17, 262–281. [Google Scholar] [CrossRef]
- Piškur, J.; Rozpędowska, E.; Polakova, S.; Merico, A.; Compagno, C. How did Saccharomyces evolve to become a good brewer? Trends Genet. 2006, 22, 183–186. [Google Scholar] [CrossRef]
- Hagman, A.; Säll, T.; Compagno, C.; Piskur, J. Yeast “make-accumulate-consume” life strategy evolved as a multi-step process that predates the whole genome duplication. PLoS ONE 2013, 8, e68734. [Google Scholar] [CrossRef]
- Rosas-Lemus, M.; Uribe-Alvarez, C.; Chiquete-Félix, N.; Uribe-Carvajal, S. In Saccharomyces cerevisiae fructose-1,6-bisphosphate contributes to the Crabtree effect through closure of the mitochondrial unspecific channel. Arch. Biochem. Biophys. 2014, 555–556, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Avenot, H.F.; Michailides, T.J. Progress in understanding molecular mechanisms and evolution of resistance to succinate dehydrogenase inhibiting (SDHI) fungicides in phytopathogenic fungi. Crop Prot. 2010, 29, 643–651. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Arroyo, T.; Valero, E. A long term field study of the effect of fungicides penconazole and sulfur on yeasts in the vineyard. Int. J. Food Microbiol. 2014, 189, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Lavaisse, L.M.; Hollmann, A.; Nazareno, M.A.; Disalvo, E.A. Zeta potential changes of Saccharomyces cerevisiae during fermentative and respiratory cycles. Colloids Surf. B Biointerfaces 2019, 174, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Ancín-Azpilicueta, C.; Jiménez-Moreno, N.; Sola-Larrañaga, C. Chapter 9–Wine. In Innovations in Traditional Foods; Galanakis, C.M., Ed.; Woodhead Publishing: Cambridge, UK, 2019; pp. 221–256. [Google Scholar] [CrossRef]
- Roukas, T. Continuous ethanol production from carob pod extract by immobilized Saccharomyces cerevisiae in a packed-bed reactor. J. Chem. Technol. Biotechnol. 1994, 59, 387–393. [Google Scholar] [CrossRef]
- Roukas, T. Ethanol production from non-sterilized beet molasses by free and immobilized Saccharomyces cerevisiae cells using fed-batch culture. J. Food Eng. 1996, 27, 87–96. [Google Scholar] [CrossRef]
- Sanchez-Segado, S.; Salar-García, M.J.; Ortiz-Martínez, V.M.; de los Ríos, A.P.; Hernández-Fernández, F.J.; Lozano-Blanco, L.J. Evaluation of ionic liquids as in situ extraction agents during the alcoholic fermentation of carob pod extracts. Fermentation 2019, 5, 90. [Google Scholar] [CrossRef]
- Sasaki, K.; Tsuge, Y.; Sasaki, D.; Kawaguchi, H.; Sazuka, T.; Ogino, C.; Kondo, A. Repeated ethanol production from sweet sorghum juice concentrated by membrane separation. Bioresour. Technol. 2015, 186, 351–355. [Google Scholar] [CrossRef]
- Chan-u-tit, P.; Laopaiboon, L.; Jaisil, P.; Laopaiboon, P. High level ethanol production by nitrogen and osmoprotectant supplementation under very high gravity fermentation conditions. Energies 2013, 6, 884–899. [Google Scholar] [CrossRef] [Green Version]
- Vallet, C.; Saïd, R.; Rabiller, C.; Martin, M.L. Natural abundance isotopic fractionation in the fermentation reaction: Influence of the Nature of the Yeast. Bioorganic Chem. 1996, 24, 319–330. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, H. Ethanol fermentation of acid-hydrolyzed cellulosic pyrolysate with Saccharomyces cerevisiae. Bioresour. Technol. 2003, 90, 95–100. [Google Scholar] [CrossRef]
- Najafpour, G.; Younesi, H.; Syahidah Ku Ismail, K. Ethanol fermentation in an immobilized cell reactor using Saccharomyces cerevisiae. Bioresour. Technol. 2004, 92, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.R.; Sepúlveda, M.D.C.; Rubio, M.C. Bio-concentration of vinasse from the alcoholic fermentation of sugar cane molasses. Waste Manag. 2000, 20, 581–585. [Google Scholar] [CrossRef]
- Díez-Antolínez, R.; Hijosa-Valsero, M.; Paniagua-García, A.I.; Gómez, X. Very-high-gravity fermentation of non-supplemented cheese whey permeate by immobilized Kluyveromyces marxianus. Chem. Eng. Trans. 2016, 49, 529–534. [Google Scholar] [CrossRef]
- Ergun, M.; Ferda Mutlu, S. Application of a statistical technique to the production of ethanol from sugar beet molasses by Saccharomyces cerevisiae. Bioresour. Technol. 2000, 73, 251–255. [Google Scholar] [CrossRef]
- Baptista, C.M.S.G.; Cóias, J.M.A.; Oliveira, A.C.M.; Oliveira, N.M.C.; Rocha, J.M.S.; Dempsey, M.J.; Lannigan, K.C.; Benson, P.S. Natural immobilisation of microorganisms for continuous ethanol production. Enzyme Microb. Technol. 2006, 40, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Kopsahelis, N.; Kanellaki, M.; Bekatorou, A. Low temperature brewing using cells immobilized on brewer’s spent grains. Food Chem. 2007, 104, 480–488. [Google Scholar] [CrossRef]
- Wang, R.; Ji, Y.; Melikoglu, M.; Koutinas, A.; Webb, C. Optimization of innovative ethanol production from wheat by response surface methodology. Process Saf. Environ. Prot. 2007, 85, 404–412. [Google Scholar] [CrossRef]
- Cáceres-Farfán, M.; Lappe, P.; Larqué-Saavedra, A.; Magdub-Méndez, A.; Barahona-Pérez, L. Ethanol production from henequen (Agave fourcroydes Lem.) juice and molasses by a mixture of two yeasts. Bioresour. Technol. 2008, 99, 9036–9039. [Google Scholar] [CrossRef]
- Habegger, L.; Rodrigues Crespo, K.; Dabros, M. Preventing overflow metabolism in Crabtree-positive microorganisms through on-line monitoring and control of fed-batch fermentations. Fermentation 2018, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Crabtree, H.G. Observations on the carbohydrate metabolism of tumours. Biochem. J. 1929, 23, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Aldiguier, A.S.; Alfenore, S.; Cameleyre, X.; Goma, G.; Uribelarrea, J.L.; Guillouet, S.E.; Molina-Jouve, C. Synergistic temperature and ethanol effect on Saccharomyces cerevisiae dynamic behaviour in ethanol bio-fuel production. Bioprocess Biosystesms Eng. 2004, 26, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Cabras, P.; Farris, G.A.; Fiori, M.G.; Pusino, A. Interaction between fenhexamid and yeasts during the alcoholic fermentation of Saccharomyces cerevisiae. J. Agric. Food Chem. 2003, 51, 5012–5015. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Masek, A. Saccharomyces Cerevisiae cell wall components as tools for ochratoxin A decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium Type | Time (Hours) | Flask Type | TS0 (g/L) | TSr (g/L) | X (g/L) | Ethmax (g/L) | ΥEth/TS (g/g) | Glol (g/L) | YX/TS (g/g) |
|---|---|---|---|---|---|---|---|---|---|
| Glucose-based, salts added | 72 | 250 mL | 214.7 | 92.5 | 5.5 | 47.6 | 0.39 | 3.9 | 0.045 |
| Enriched grape must, no salts | 73 | 250 mL | 222.6 | 6.6 | 7.9 | 82.3 | 0.38 | 4.1 | 0.037 |
| Enriched grape must, no salts | 49 | 2.0 L | 211.8 | 2.6 | 9.8 | 85.8 | 0.41 | 4.2 | 0.047 |
| (a) | ||||||||
| Time (h) | Myclobutanil (mg/L) | TS0 (g/L) | TSr (g/L) | X (g/L) | Glol (g/L) | Ethmax (g/L) | ΥEth/TS (g/g) | YX/TS (g/g) |
| 45 | - | 147.8 | 2.4 | 9.1 | 3.8 | 68 | 0.47 | 0.06 |
| 46 | 0.1 | 150.2 | 1.7 | 9.3 | 4.4 | 60.1 | 0.4 | 0.06 |
| 44 | 1 | 156.2 | 5.1 | 8.4 | 4.2 | 62.2 | 0.41 | 0.06 |
| (b) | ||||||||
| Time (h) | Myclobutanil (mg/L) | TS0 (g/L) | TSr (g/L) | X (g/L) | Glol (g/L) | Ethmax (g/L) | ΥEth/TS (g/g) | YX/TS (g/g) |
| 68 | - | 243.2 | 1 | 7.6 | 5.4 | 112.3 | 0.46 | 0.03 |
| 73 | 0.1 | 248.3 | 3.2 | 10.2 | 5.2 | 105.2 | 0.43 | 0.04 |
| 69 | 1 | 252.2 | 3 | 9.5 | 5.3 | 105 | 0.42 | 0.04 |
| (a) | ||||||||
| Time (h) | Myclobutanil (mg/L) | TS0 (g/L) | TSr (g/L) | X (g/L) | Glol (g/L) | Ethmax (g/L) | ΥEth/TS (g/g) | YX/TS (g/g) |
| 46 | - | 167.6 | 3.5 | 9.6 | 2.4 | 79 | 0.48 | 0.06 |
| 48 | 0.1 | 160 | 2.2 | 10 | 2.4 | 62.6 | 0.4 | 0.06 |
| 48 | 1 | 160.6 | 5.2 | 9.8 | 2.2 | 62.4 | 0.4 | 0.06 |
| (b) | ||||||||
| Time (h) | Myclobutanil (mg/L) | TS0 (g/L) | TSr (g/L) | X (g/L) | Glol (g/L) | Ethmax (g/L) | ΥEth/TS (g/g) | YX/TS (g/g) |
| 68 | - | 259.3 | 2.5 | 9.6 | 3.5 | 125 | 0.49 | 0.04 |
| 73 | 0.1 | 250 | 3.8 | 10.6 | 3.5 | 112.2 | 0.45 | 0.04 |
| 73 | 1 | 255.2 | 4 | 10.5 | 3 | 112 | 0.45 | 0.04 |
| Yeast Strain | Carbon Source | Initial Sugar Concentration (g/L) | EtOH (g/L) | Reference |
|---|---|---|---|---|
| Bakers’ yeast | Carob pod | 200–350 | ~62 | [62] |
| Bakers’ yeast | Molasses | 150–300 | 53.0 | [63] |
| S. cerevisiae | Carob pod extract (Saccharose) | 200 | 95 | [64] |
| S. cerevisiae NP01 | Sweet sorghum juice | 280–300 | 134.3 | [12] |
| S. cerevisiae BY4741 | Sweet sorghum juice concentrated | 278.6 | 113.7 | [65] |
| S. cerevisiae NP01 | Sucrose | 280 | 95.3 | [66] |
| S. cerevisiae 27817 | Glucose | 50–200 | 5.1–91.8 | [67] |
| S. cerevisiae 2399 | Glucose | 32 | 13.7 | [68] |
| S. cerevisiae 24860 | Glucose | 150 | 48.0 | [69] |
| S. cerevisiae CMI237 | Sugar | 160 | 70.0 | [70] |
| K. marxianus | Lactose | 170–190 | 83.2 | [71] |
| S. cerevisiae ATCC 24860 | Molasses | 2–50 | 5.0–18.4 | [72] |
| S. cerevisiae NCYC 1119 | Molasses | 100 | 40.0 | [73] |
| S. cerevisiae AXAZ-1 | Molasses | ~216 | 71.3 | [74] |
| S. cerevisiae ATCC 26602 | Flour hydrolysates | 150 | 76 | [75] |
| S. cerevisiae sp. & K. marxianus blends | Henequen juice & molasses blends | ~215 | 41.2 | [76] |
| S. cerevisiae MAK-1 | Grape must | 250 | 106.4–119.2 | [30] |
| S. cerevisiae MAK-1 | Olive-mill wastewater & glucose | 115 | 52.0 | [48] |
| S. cerevisiae MAK-1 | Olive-mill wastewater & molasses | 135 | 52.4 | [46] |
| S. cerevisiae LMBF-Y 16 | Grape must | 250 | 112.3 | Present study |
| S. cerevisiae LMBF-Y 18 | Grape must | 250 | 125.0 | Present study |
| (a) | ||||
| Saccharomyces cerevisiae LMBF-Y 16 | ||||
| Initial sugars (g/L) | ~150 | ~250 | ~150 | ~250 |
| Myclobutanil (mg/L) | 1 | 1 | 0.1 | 0.1 |
| Ethanol (g/L) | 62.2 | 105 | 60.1 | 105.2 |
| Biomass (g/L) | 8.4 | 9.5 | 9.3 | 10.2 |
| Myclobutanil decomposition % | 5 | 16 | 23 | 27 |
| (b) | ||||
| Saccharomyces cerevisiae LMBF-Y 18 | ||||
| Sugars (g/L) | ~150 | ~250 | ~150 | ~250 |
| Myclobutanil (mg/L) | 1 | 1 | 0.1 | 0.1 |
| Ethanol (g/L) | 62.4 | 112 | 62.6 | 112.2 |
| Biomass (g/L) | 9.8 | 10.5 | 10 | 10.6 |
| Myclobutanil decomposition % | 6 | 9 | 16 | 19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terpou, A.; Dimopoulou, M.; Belka, A.; Kallithraka, S.; Nychas, G.-J.E.; Papanikolaou, S. Effect of Myclobutanil Pesticide on the Physiological Behavior of Two Newly Isolated Saccharomyces cerevisiae Strains during Very-High-Gravity Alcoholic Fermentation. Microorganisms 2019, 7, 666. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120666
Terpou A, Dimopoulou M, Belka A, Kallithraka S, Nychas G-JE, Papanikolaou S. Effect of Myclobutanil Pesticide on the Physiological Behavior of Two Newly Isolated Saccharomyces cerevisiae Strains during Very-High-Gravity Alcoholic Fermentation. Microorganisms. 2019; 7(12):666. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120666
Chicago/Turabian StyleTerpou, Antonia, Maria Dimopoulou, Aikaterini Belka, Stamatina Kallithraka, George-John E. Nychas, and Seraphim Papanikolaou. 2019. "Effect of Myclobutanil Pesticide on the Physiological Behavior of Two Newly Isolated Saccharomyces cerevisiae Strains during Very-High-Gravity Alcoholic Fermentation" Microorganisms 7, no. 12: 666. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120666