Biochar Suppresses Bacterial Wilt of Tomato by Improving Soil Chemical Properties and Shifting Soil Microbial Community

Abstract
:1. Introduction
2. Materials and Methods
2.1. Biochar and Soil Characteristics
2.2. Plant Growth and R. Solanacearum Culture
2.3. Experiment Design
2.4. Disease Severity Survey
2.5. Soil Sampling and Analysis
2.6. Soil DNA Extraction, PCR Amplification, and Illumina MiSeq Sequencing
2.7. Statistical Analysis
3. Results
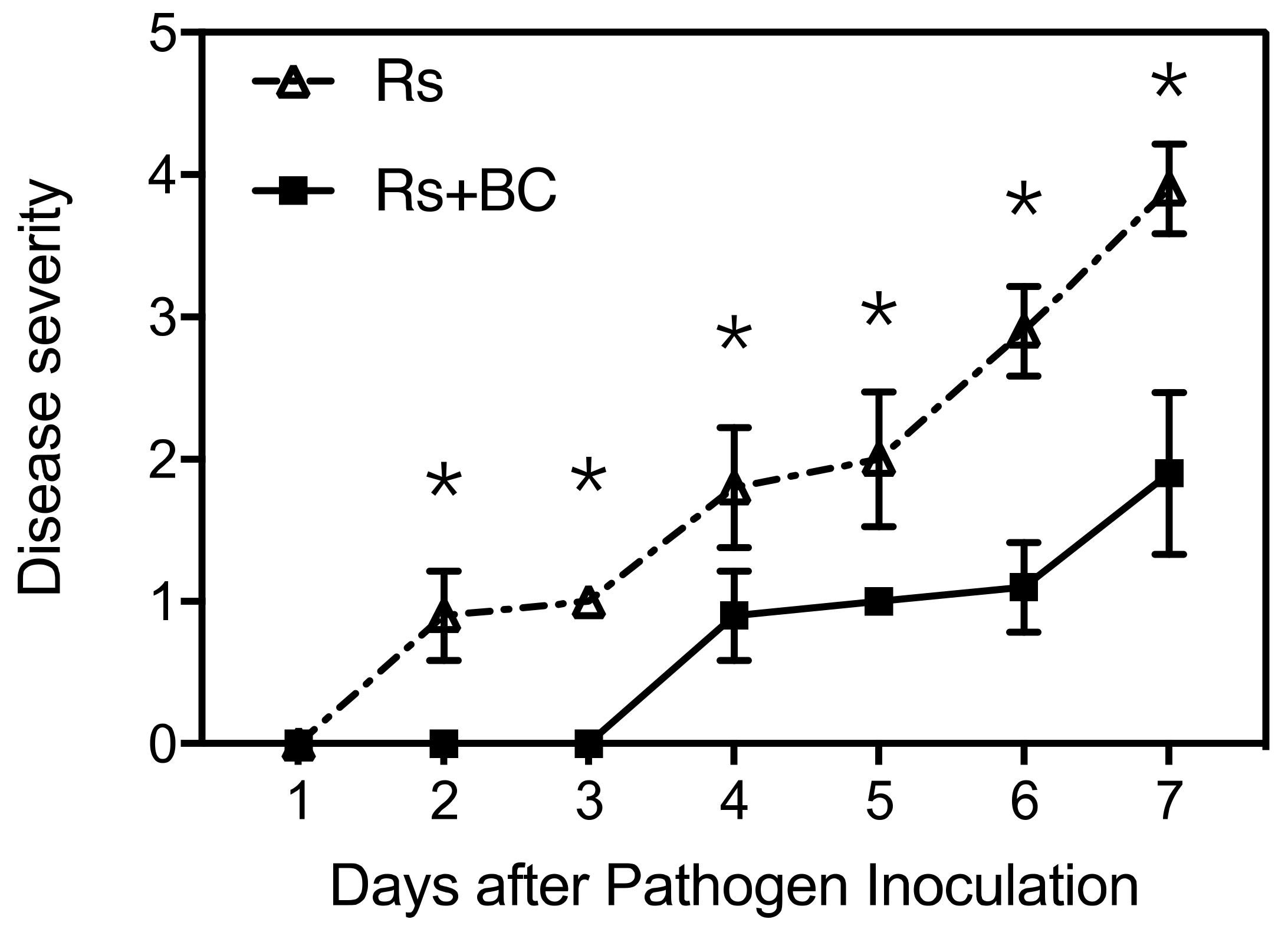
3.1. Disease Severity of Bacterial Wilt
3.2. Effects of Biochar on Soil Chemical Properties
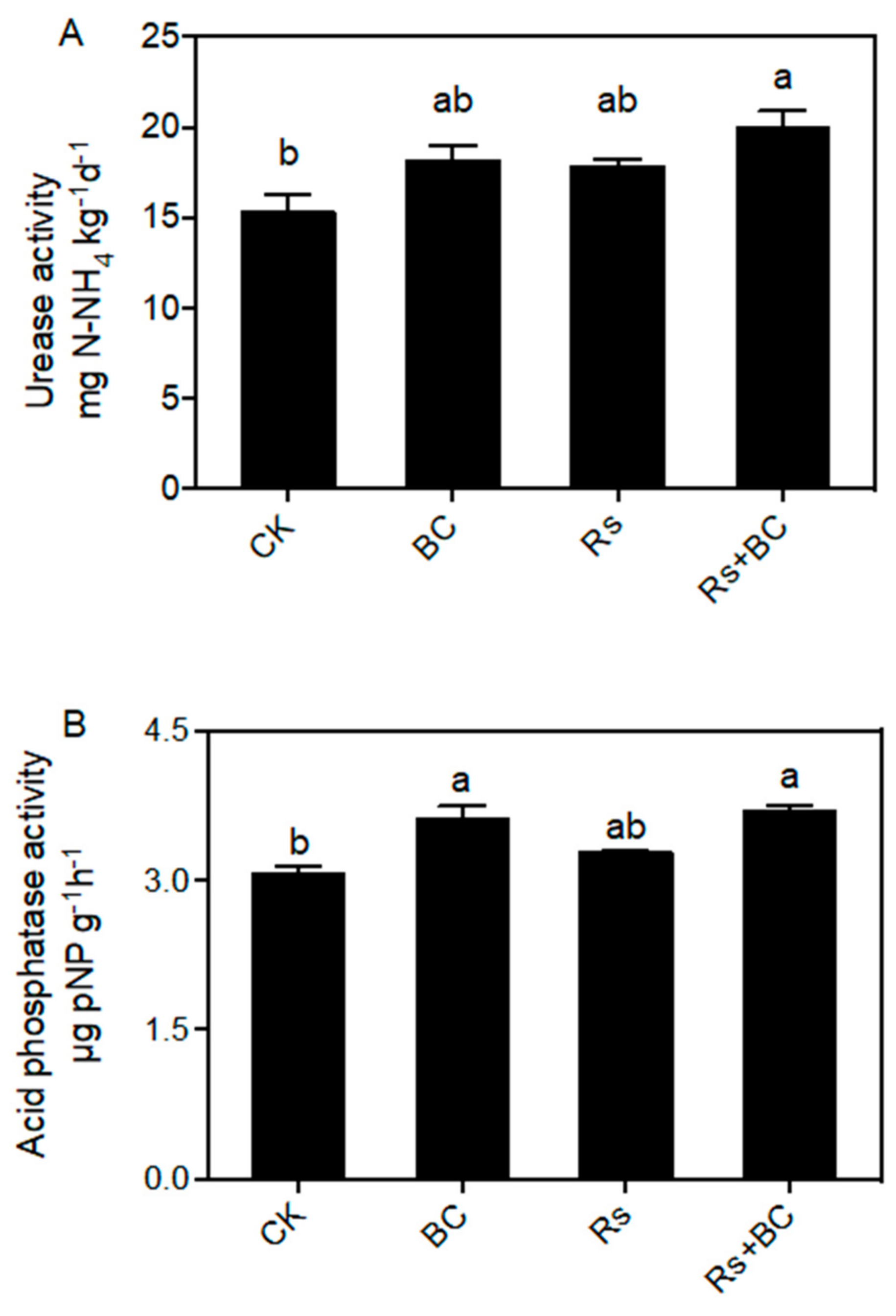
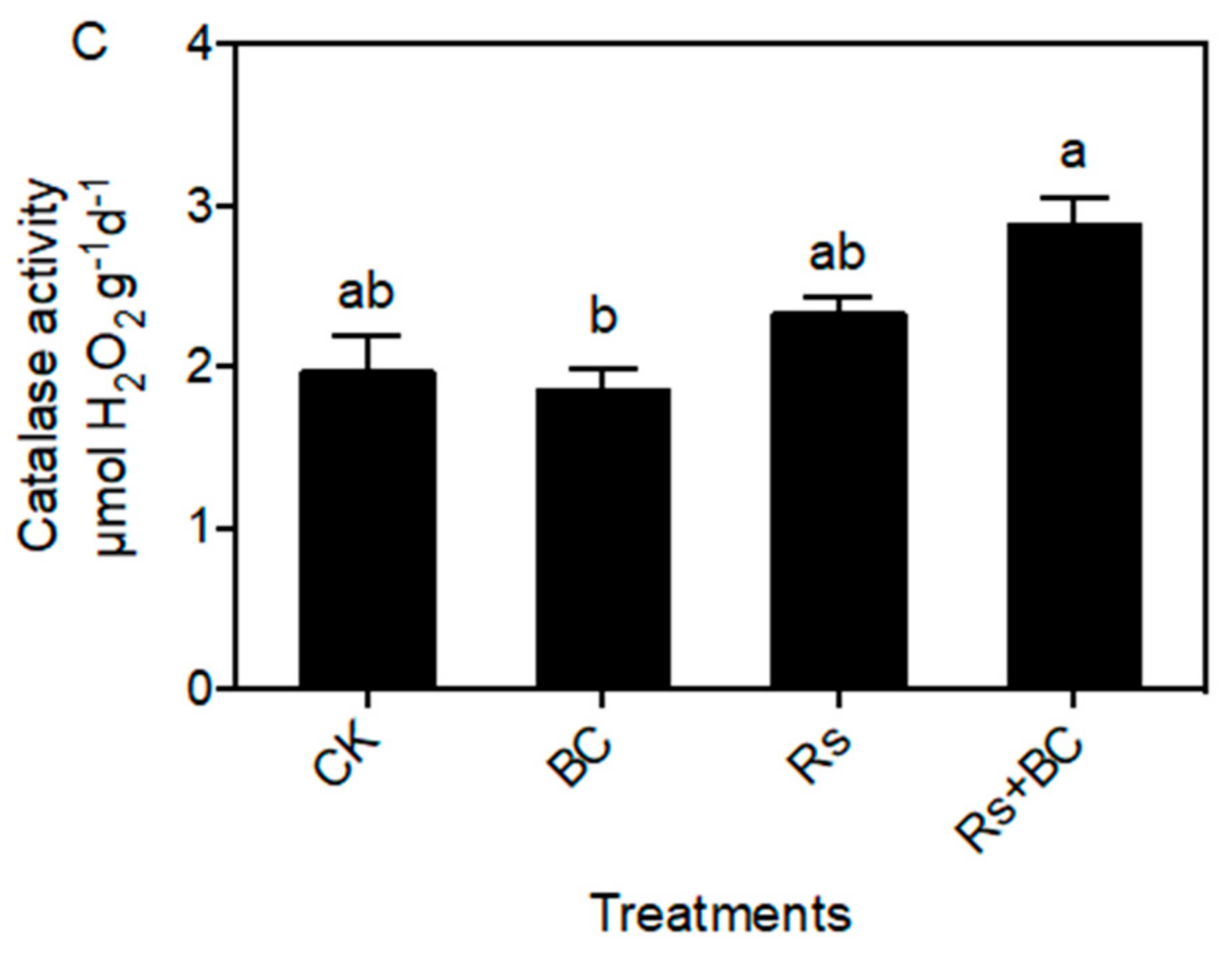
3.3. Effects of Biochar on Soil Enzyme Activities
3.4. Effects of Biochar Application on Soil Bacterial Taxonomic Richness
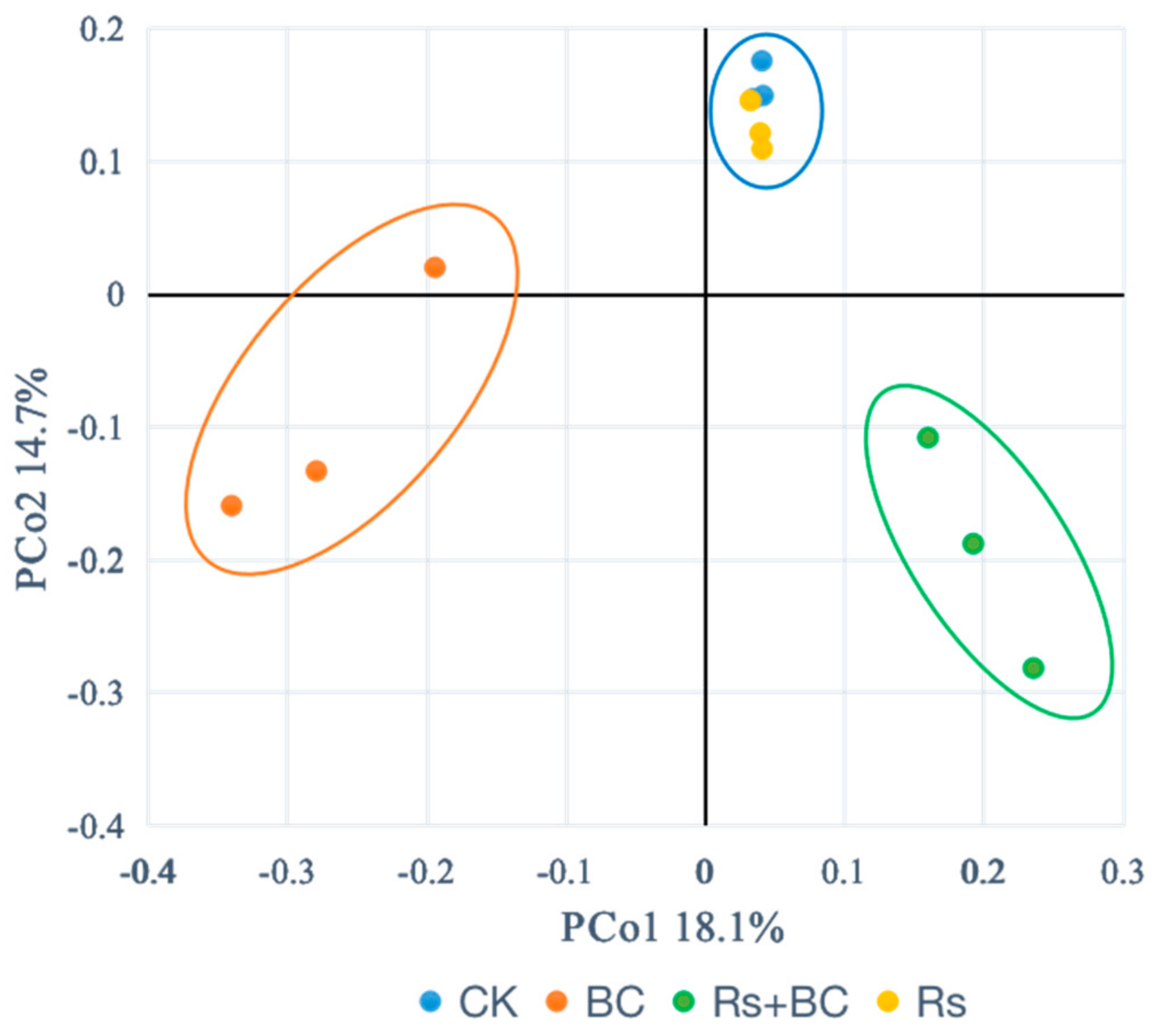
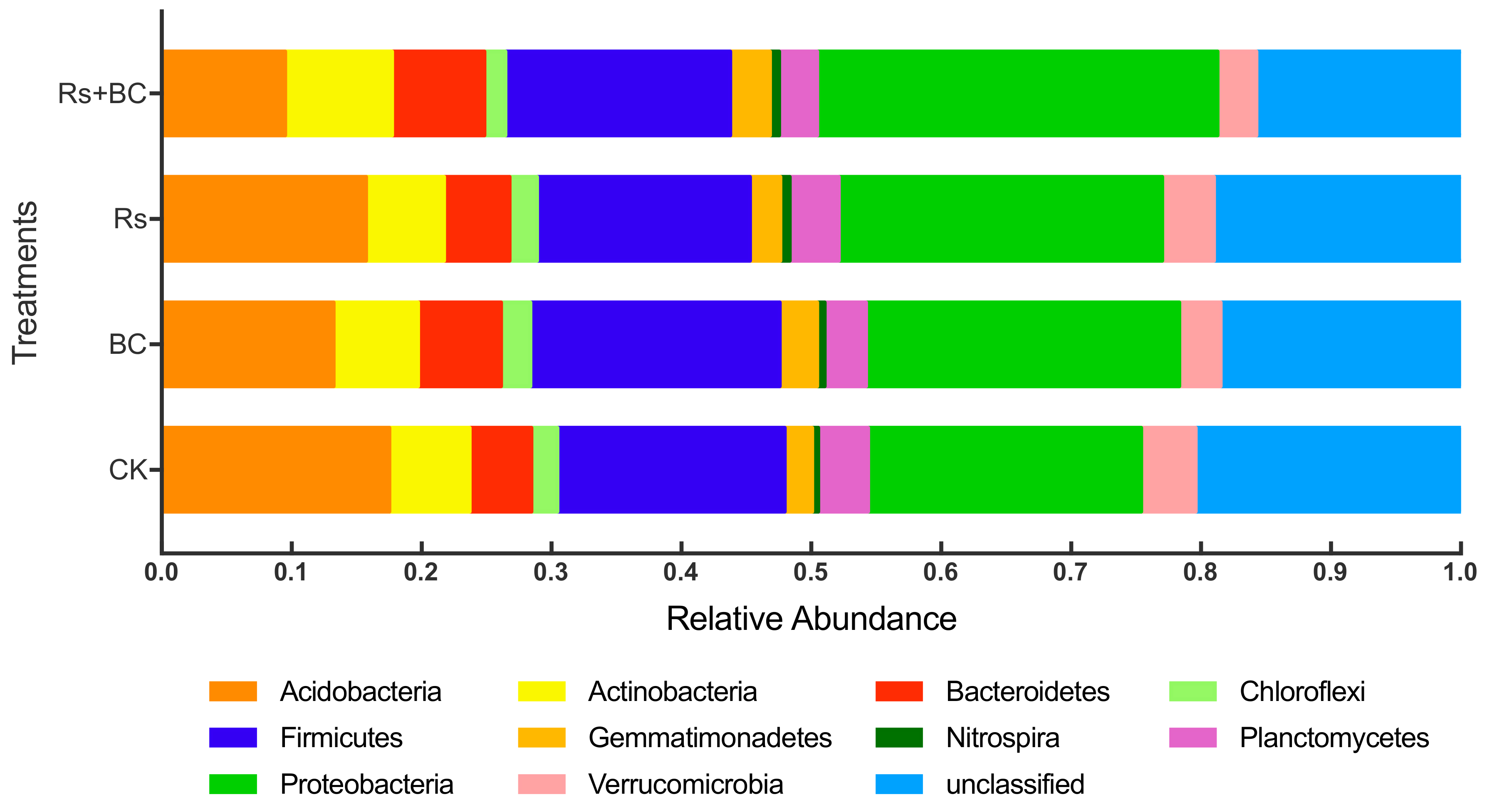
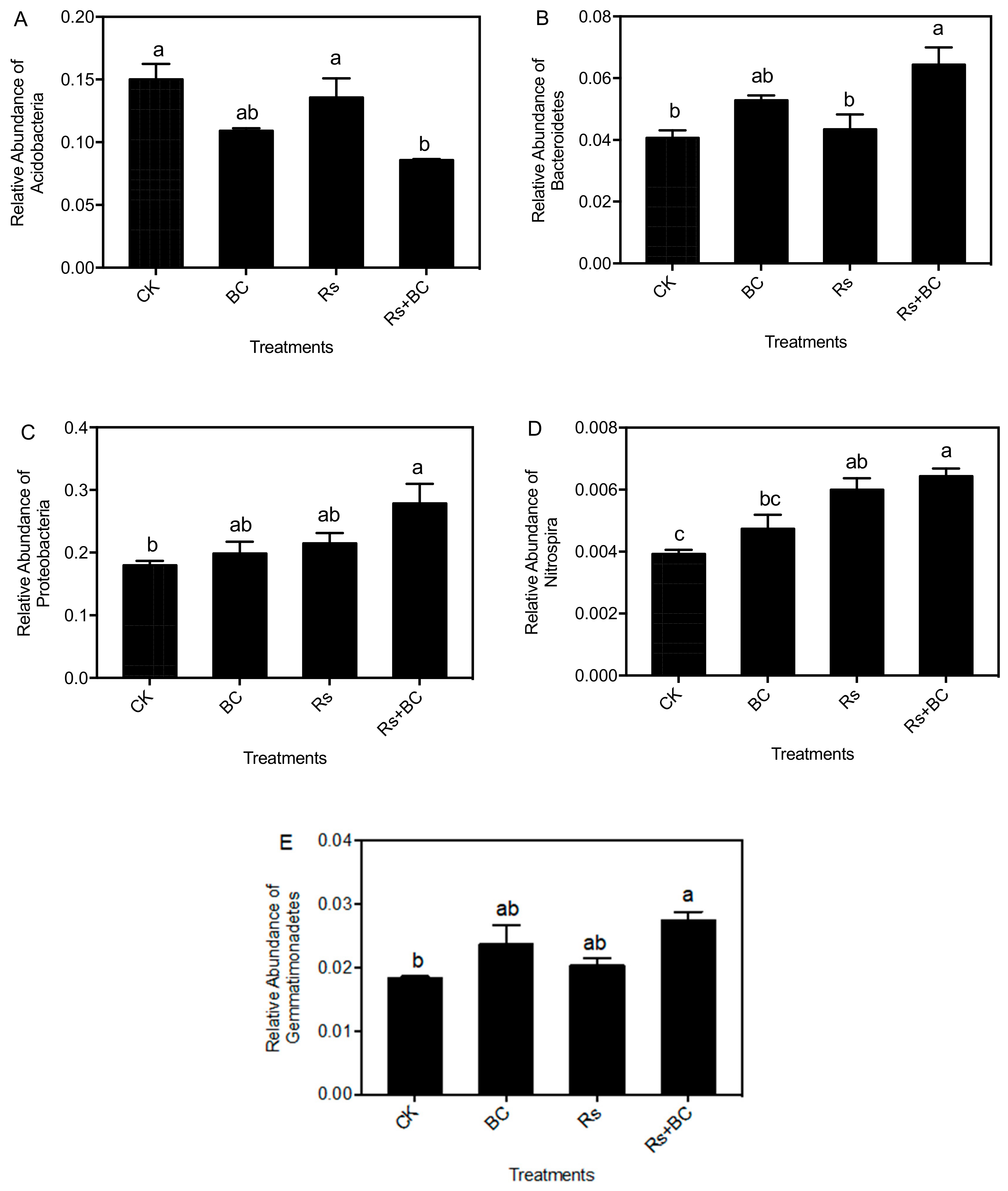
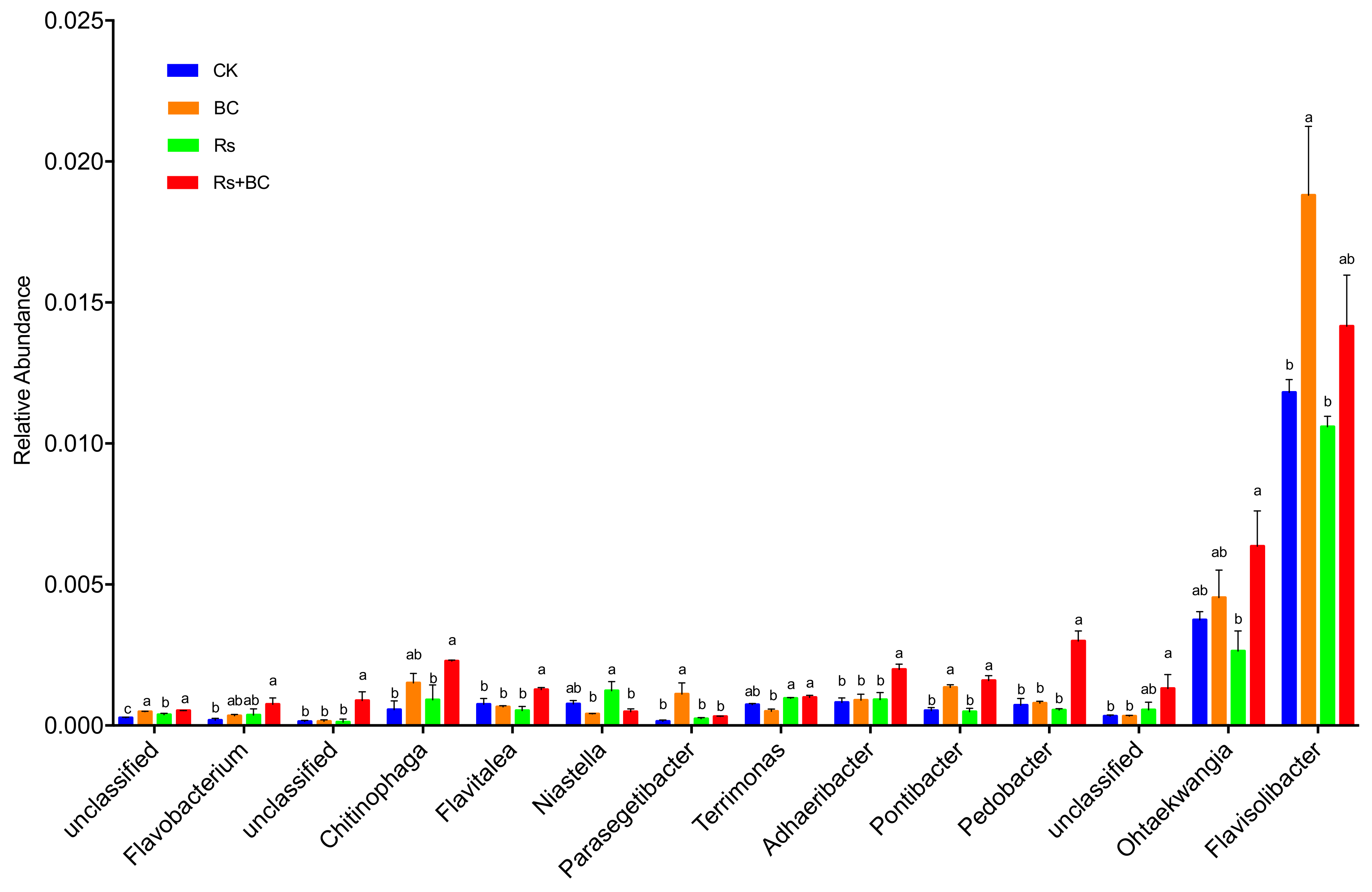
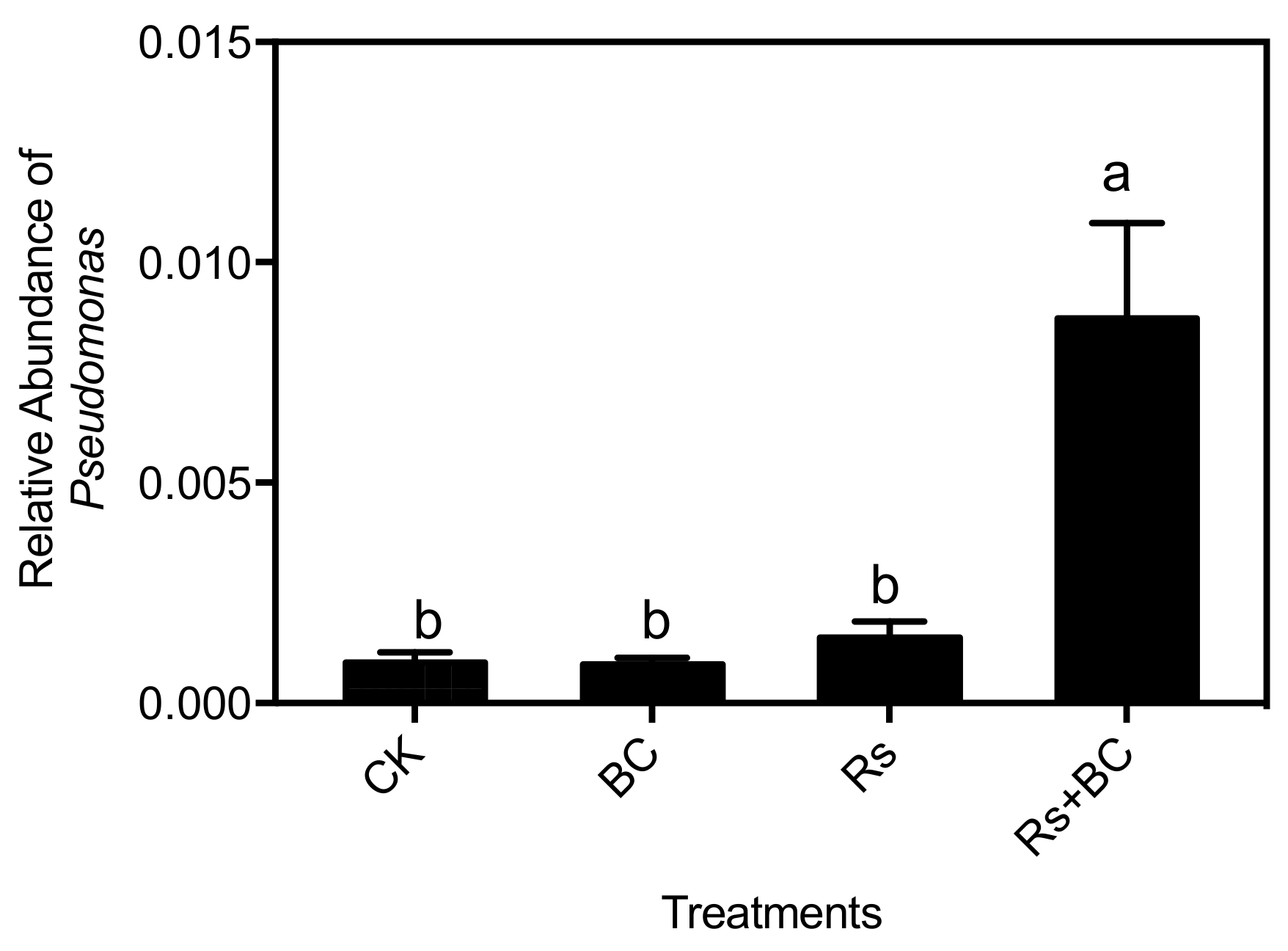
3.5. Effects of Biochar Application on Soil Bacterial Community Composition
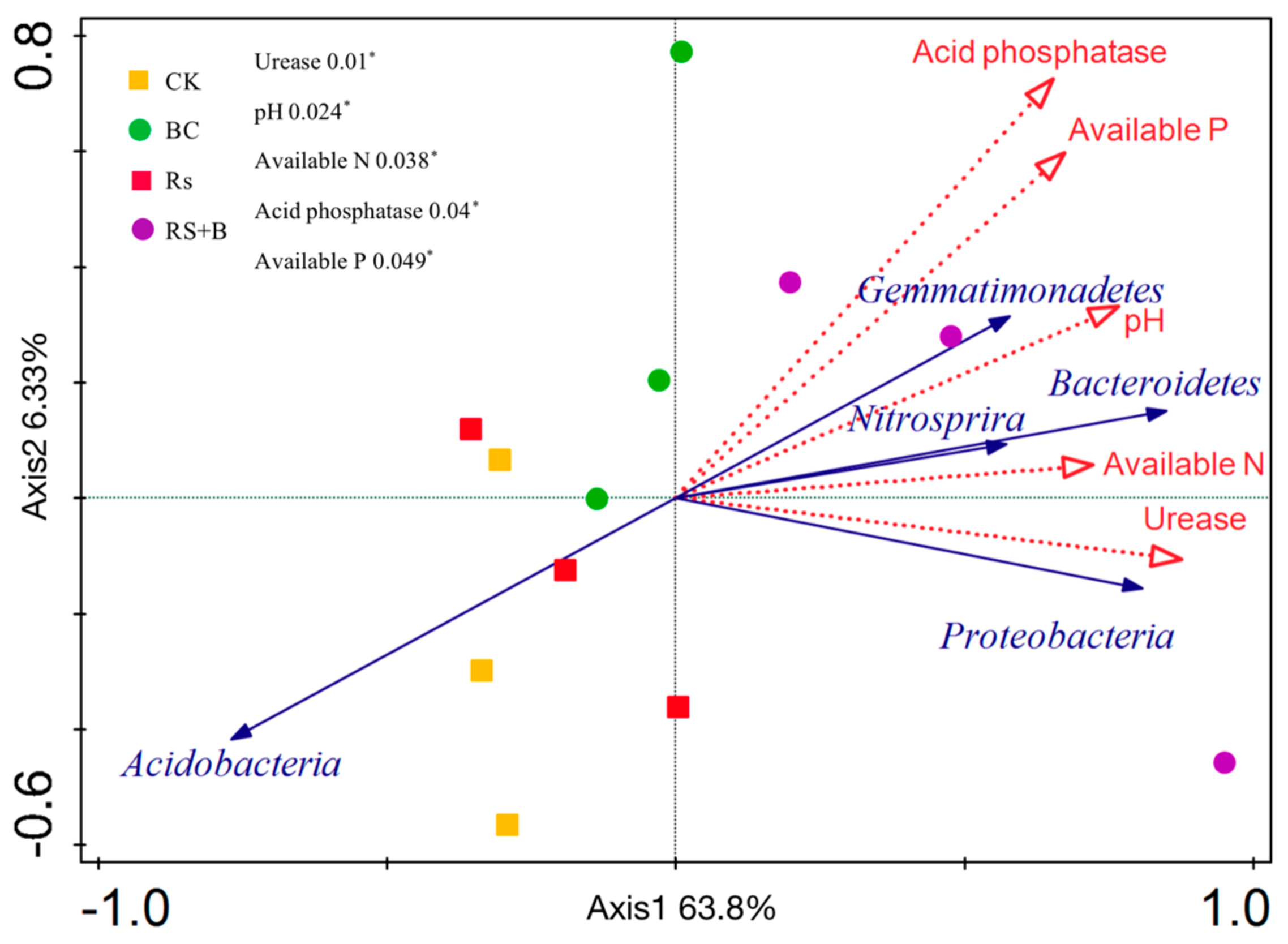
3.6. The Relationship between Soil Properties and Rhizosphere Bacterial Community
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wicker, E.; Grassart, L.; Coranson-Beaudu, R.; Mian, D.; Guilbaud, C.; Fegan, M.; Prior, P. Ralstonia solanacearum strains from Martinique (French West Indies) exhibiting a new pathogenic potential. Appl. Environ. Microb. 2007, 71, 6790–6801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhanang, P.M.; Momol, M.T.; Olson, S.M.; Jones, J. Effects of plant essential oils on Ralstonia solanacearum population density and bacterial wilt incidence in tomato. Plant Dis. 2003, 87, 423–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trigalet, A.; Trigalet, D. Invasiveness in tomato plant of Tn5-induced avirulent mutants of Pseudomonas solanacearun. Physiol. Mol. Plant Pathol. 1986, 28, 423–430. [Google Scholar] [CrossRef]
- Trigalet, A.; Trigalet-Demery, D. Use of avirulent mutants of Pseudomonas solanacearum for the biological control of bacterial wilt of tomato plants. Physiol. Mol. Plant Pathol. 1990, 36, 27–38. [Google Scholar] [CrossRef]
- Yuliar; Nion, Y.A.; Toyota, K.K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Environ. 2015, 30, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, K.G.; Gloy, B.A.; Joseph, S.; Scott, N.R.; Lehmann, J. Life cycle assessment of biochar systems: Estimating the energetic, economic, and climate change potential. Environ. Sci. Technol. 2009, 4, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Golber, E.D. Black Carbon in the Environment: Properties and Distribution; John & Wiley: New York, NY, USA, 1985. [Google Scholar]
- Glaser, B.; Haumaier, L.; Guggenberger, G.; Zech, W. The ‘Terra Preta’ phenomenon: A model for sustainable agriculture in the humid tropics. Naturwissenschaften 2001, 88, 37–41. [Google Scholar] [CrossRef]
- Zhang, W.L.; Li, G.H.; Gao, W.D. Effect of biomass charcoal on soil character and crop yield. Chin. Agric. Sci. Bull. 2009, 25, 153–157. [Google Scholar]
- Liang, B.; Lehmann, J.; Solomon, D.; Kinyangi, J.; Grossman, J.; O’Neill, B.O.; Skjemstad, J.O.; Thies, J.; Luizão, F.J.; Petersen, J.; et al. Black carbon increases cation exchange capacity in soils. Soil Sci. Soc. Am. J. 2006, 70, 1719–1730. [Google Scholar] [CrossRef] [Green Version]
- Kasozi, G.N.; Zimmerman, A.R.; Nkedi-Kizza, P.; Gao, B. Catechol and humic acid sorption onto a range of laboratory-produced black carbons (biochars). Environ. Sci. Technol. 2010, 44, 6189–6195. [Google Scholar] [CrossRef]
- Shi, M.; Lin, C.H.; Huang, Z.Q.; Si, H.P.; Lin, K.Y. The influence of bio-char inputting on the adsorption of phenanthrene by soils and by maize seedlings. J. Agro-Environ. Sci. 2011, 30, 912–916. [Google Scholar]
- Novak, J.M.; Busscher, W.J.; Laird, D.L.; Ahmedna, M.; Watts, D.W.; Niandou, M.A. Impact of biochar amendment on fertility of a southeastern coastal plain soil. Soil Sci. 2009, 174, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Uchimiya, M.; Klasson, K.T.; Wartelle, L.H.; Lima, I.M. Influence of soil properties on heavy metal sequestration by biochar amendment: 1. Copper sorption isotherms and the release of cations. Chemosphere 2011, 82, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y.; Cytryn, E.; Harel, Y.M.; Lew, B.; Graber, E.R. The biochar effect: Plant resistance to biotic stresses. Phytopathol. Mediterr. 2011, 50, 335–349. [Google Scholar]
- Schmidt, H.P.; Kammann, C.; Niggli, C.; Evangelou, M.W.; Mackie, K.A.; Abiven, S. Biochar and biochar-compost as soil amendments to a vineyard soil: Influences on plant growth, nutrient uptake, plant health and grape quality. Agri. Ecosyst. Environ. 2014, 191, 117–121. [Google Scholar] [CrossRef]
- Harel, Y.M.; Elad, Y.; Rav-David, D.; Borenstein, M.; Shulchani, R.; Lew, B.; Graber, E.R. Biochar mediates systemic response of strawberry to foliar fungal pathogens. Plant Soil. 2012, 357, 245–257. [Google Scholar] [CrossRef]
- Elad, Y.; David, D.R.; Harel, Y.M.; Borenshtein, M.; Kalifa, H.B.; Silber, A.; Graber, E.R. Induction of systemic resistance in plants by biochar, a soil-applied carbon sequestering agent. Phytopathology 2010, 100, 913–921. [Google Scholar] [CrossRef] [Green Version]
- Elmer, W.H.; Pignatello, J.J. Effect of biochar amendments on mycorrhizal associations and Fusarium crown and root rot of asparagus in replant soils. Plant Dis. 2011, 95, 960–966. [Google Scholar] [CrossRef] [Green Version]
- Graber, E.R.; Frenkel, O.; Jaiswal, A.K.; Elad, Y. How may biochar influence severity of diseases caused by soilborne pathogens? Carbon Manag. 2014, 5, 169–183. [Google Scholar] [CrossRef]
- Bonanomi, G.; Ippolito, F.; Scala, F. A” black” future for plantpathology? Biochar as a new soil amendment for controlling plant diseases. J. Plant Pathol. 2015, 97, 223–234. [Google Scholar]
- Akhter, A.; Hage-Ahmed, K.; Soja, G.; Steinkellner, S. Compost and biochar alter mycorrhization, tomato root exudation, and development of Fusarium oxysporum f. sp. lycopersici. Front. Plant Sci. 2015, 6, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerome, M.; Toyota, K.; Islam, T.; Nishijima, T.; Matsuoka, T.; Sato, K.; Yamaguchi, Y. Suppression of bacterial wilt of tomato by incorporation of municipal biowaste charcoal into soil. Soil Microorg. (Japan) 2005, 59, 0912–2184. [Google Scholar]
- Zhang, C.; Lin, Y.; Tian, X.; XU, Q.; Chen, Z.H.; Lin, W. Tobacco bacterial wilt suppression with biochar soil addition associates to improved soil physiochemical properties and increased rhizosphere bacteria abundance. Appl. Soil Ecol. 2017, 112, 90–96. [Google Scholar] [CrossRef]
- Noguera, D.; Rondón, M.; Laossi, K.R.; Hoyos, V.; Lavelle, P.; Carvalho, M.H.C.; Barot, S. Contrasted effect of biochar and earthworms on rice growth and resource allocation in different soils. Soil Biol. Biochem. 2010, 42, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—a review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Kolton, M.; Harel, Y.M.; Pasternak, Z.; Graber, E.R.; Elad, Y.; Cytryn, E. Impact of biochar application to soil on the root-associated bacterial community structure of fully developed greenhouse pepper plants. Appl. Environ. Microbiol. 2011, 77, 4924–4930. [Google Scholar] [CrossRef] [Green Version]
- Mehari, Z.H.; Elad, Y.; Rav-David, D.; Graber, E.R.; Harel, Y.M. Induced systemic resistance in tomato (Solanum lycopersicum) against Botrytis cinerea by biochar amendment involves jasmonic acid signaling. Plant Soil. 2015, 395, 31–44. [Google Scholar] [CrossRef]
- Kolton, M.; Graber, E.R.; Tsehansky, L.; Elad, Y.; Cytryn, E. Biochar-stimulated plant performance is strongly linked to microbial diversity and metabolic potential in the rhizosphere. New Phytol. 2017, 213, 1393–1404. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Elad, Y.; Paudel, I.; Graber, E.R.; Cytryn, E.; Frenkel, O. Linking the belowground microbial composition, diversity and activity to soilborne disease suppression and growth promotion of tomato amended with biochar. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Rogovska, N.; Laird, D.; Leandro, L.; Aller, D. Biochar effect on severity of soybean root disease caused by Fusarium virguliforme. Plant Soil. 2017, 413, 111–126. [Google Scholar] [CrossRef]
- Lu, Y.; Rao, S.; Huang, F.; Cai, Y.X.; Cai, K.Z. Effects of biochar amendment on tomato bacterial wilt resistance, soil microbial amount and activity. Int. J. Agron. 2016, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Elie, D.; Wydra, K. Interaction between silicon amendment, bacterial wilt development and phenotype of Ralstonia solanacearum in tomato genotypes. Physiol. Mol. Plant Pathol. 2004, 64, 233–243. [Google Scholar]
- Fang, Z. Research Methods of Plant Pathology; China Agriculture Press: Beijing, China, 1998. [Google Scholar]
- Wang, S.; Tian, H.; Liu, J.; Pan, S. Pattern and change of soil organic carbon storage in China: 1960s–1980s. Tellus B Chem. Phys. Meteorol. 2003, 55, 416–427. [Google Scholar]
- Lu, R.K. Analysis Method of Soil Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; United States Department of Agriculture: Washington, DC, USA, 1954.
- Zantua, M.I.; Bremner, J.M. Stability of urease in soils. Soil Biol. Biochem. 1977, 9, 135–140. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Havir, E.; Mchale, N.A. Biochemical and developmental characterization of multiple forms of catalases in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.; Brandt, K.K.; Al-Soud, W.A.; Holm, P.E.; Hansen, L.H.; Sørensen, S.J.; Nybroe, O. Selection for Cu-tolerant bacterial communities with altered composition, but unaltered richness, via long-term Cu exposure. Appl. Environ. Microbiol. 2012, 78, 7438–7446. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’ Index. Global Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Jaiswal, A.K.; Elad, Y.; Graber, E.R.; Frenkel, O. Rhizoctonia solani suppression and plant growth promotion in cucumber as affected by biochar pyrolysis temperature, feedstock and concentration. Soil Bio. Biochem. 2014, 69, 110–118. [Google Scholar] [CrossRef]
- Korolev, N.; David, D.R.; Elad, Y. The role of phytohormones in basal resistance and Trichoderma-induced systemic resistance to Botrytis cinerea in Arabidopsis thaliana. BioControl 2008, 53, 667–683. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate Soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Gregory, S.J.; Anderson, C.W.; Camps-Arbestain, M.; Biggs, P.J.; Ganley, A.R.; O’Sullivan, J.M.; McManus, M.T. Biochar in co-contaminated soil manipulates arsenic solubility and microbiological community structure, and promotes organochlorine degradation. PLoS ONE 2015, 10, e0125393. [Google Scholar] [CrossRef]
- Githinji, L. Effect of biochar application rate on soil physical and hydraulic properties of a sandy loam. Arch. Agron. Soil Sci. 2014, 60, 457–470. [Google Scholar] [CrossRef]
- Schulz, H.; Dunst, G.; Glaser, B. No effect level of co-composted biochar on plant growth and soil properties in a greenhouse experiment. Agronomy 2014, 4, 34–51. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, Z.; Wang, X.; Sun, Q.R.; Dong, H.Q.; Wang, G.S.; Chen, X.S.; Yin, C.M.; Han, Z.H.; Mao, Z.Q. Effects of biochar on the growth of apple seedlings, soil enzyme activities and fungal communities in replant disease soil. Sci. Hortic. 2019, 256, 108641. [Google Scholar] [CrossRef]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil enzyme activities as biological indicators of soil health. Rev. Environ. Health 2003, 18, 65–73. [Google Scholar] [CrossRef]
- Ghani, M.I.; Ali, A.; Atif, M.J.; Ali, M.; Amin, B.; Anees, M.; Khurshid, H.; Cheng, H.Z. Changes in the soil microbiome in eggplant monoculture revealed by high-throughput Illumina MiSeq Sequencing as influenced by raw garlic stalk amendment. Int. J. Mol. Sci. 2019, 20, 2125. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Mazzola, M. Soil immune responses. Science 2016, 352, 1392–1393. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Su, L.; Fang, Z.Y.; Yuan, S.F.; Wang, L.L.; Shen, B.; Shen, Q.R. Competitive use of root. exudates by Bacillus amyloliquefaciens with Ralstonia solanacearum decreases the pathogenic population density and effectively controls tomato bacterial wilt. Sci. Hortic. 2017, 218, 132–138. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; Romano, M.; Marzaioli, R.; Baglivo, I.; Baronti, S.; Miglietta, F.; Castaldi, S. Effect of biochar addition on soil microbial community in a wheat crop. Eur. J. Soil Biol. 2011, 60, 9–15. [Google Scholar] [CrossRef]
- Luo, Y.; Durenkamp, M.; Nobili, M.D.; Lin, Q.; Devonshire, B.J.; Brookes, P.C. Microbial biomass growth following incorporation of biochars produced at 350 °C or 700 °C, in a silty-clay loam soil of high and low pH. Soil Biol. Biochem. 2013, 57, 513–523. [Google Scholar] [CrossRef]
- Hu, L.; Cao, L.; Zhang, R. Bacterial and fungal taxon changes in soil microbial community composition induced by short-term biochar amendment in red oxidized loam soil. World J. Microb. Biot. 2014, 30, 1085–1092. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.H.; Li, H.; Yao, H.Y.; Su, J.Q.; Zhu, Y.G. Biochar impacts soil microbial community composition and nitrogen cycling in an acidic soil planted with rape. Environ. Sci. Technol. 2014, 48, 9391–9399. [Google Scholar] [CrossRef]
- Khodadad, C.L.; Zimmerman, A.R.; Green, S.J.; Uthandi, S.; Foster, J.S. Taxa-specific changes in soil microbial community composition induced by pyrogn-c carbon amendments. Soil Biol. Biochem. 2011, 43, 385–392. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Tan, G.C.; Wang, H.Y.; Gai, X.P. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Naumoff, D.G.; Dedysh, S.N. Lateral gene transfer between the Bacteroidetes and Acidobacteria: The case of ɑ-L-Rhamnosidases. FEBS Lett. 2012, 586, 3843–3851. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Xing, K.; Jiang, J.H.; Xu, L.H.; Li, W.J. Biodiversity, bioactive natural products and biotechnological potential of plant associated endophytic actinobacteria. Appl. Microbiol. Biot. 2011, 89, 457–473. [Google Scholar] [CrossRef]
- Haas, D.; Defago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Bakker, P.A.; Pieterse, C.M.; Loon, L.C. Induced systemic resistance by fluorescent Pseudomonas spp. Phytopathology 2007, 97, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; Voor, M.; Schneider, J.H.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Ramesh, R.; Joshi, A.A.; Ghanekar, M.P. Pseudomonads: Major antagonistic endophytic bacteria to suppress bacterial wilt pathogen, Ralstonia solanacearum in the eggplant (Solanum melongena L.). World J. Microb. Biot. 2009, 25, 47–55. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.; Bakker, P.A. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | TOC g·kg−1 | TN g·kg−1 | C/N | SOM g·kg−1 | pH | EC µS·cm−1 |
|---|---|---|---|---|---|---|
| CK | 7.3 ± 0.05 b | 0.85 ± 0.025 b | 7.16 ± 0.34 b | 12.4 ± 0.08 b | 5.9 ± 0.20 b | 111.3 ± 19.7 b |
| BC | 11.2 ± 0.56 a | 1.07 ± 0.005 a | 10.80 ± 0.4 a | 19.0 ± 0.95 a | 6.5 ± 0.05 a,b | 384.0 ± 32.0 a |
| Rs | 6.6 ± 0.12 b | 0.83 ± 0.030 b | 7.05 ± 0.52 b | 11.1 ± 0.20 b | 6.1 ± 0.10 a,b | 115.5 ± 9.5 b |
| Rs + BC | 9.5 ± 0.28 a | 0.94 ± 0.025 a | 10.20 ± 0.41 a | 16.2 ± 0.48 a | 6.7 ± 0.05 a | 380.0 ± 21.4 a |
| Treatment | Available N mg·kg−1 | Available P mg·kg−1 | Available K mg·kg−1 |
|---|---|---|---|
| CK | 420.0 ± 7.50 b | 128.8 ± 0.38 b | 162.8 ± 0.01 b |
| BC | 393.8 ± 6.25 b,c | 137.3 ± 0.56 a | 661.1 ± 0.08 a |
| Rs | 367.5 ± 5.05 c | 131.7 ± 0.85 b | 147.6 ± 0.04 b |
| Rs + BC | 520.0 ± 10.55 a | 139.8 ± 1.06 a | 736.8 ± 0.10 a |
| Treatment | Shannon Index | Simpson Index | ACE | Chao |
|---|---|---|---|---|
| CK | 6.51 ± 0.10 a | 0.99 ± 0.002 a | 6854 ± 517 a | 5590 ± 94 a |
| BC | 6.41 ± 0.12 a | 0.98 ± 0.002 a | 8805 ± 2561 a | 6674 ± 1401 a |
| Rs | 6.64 ± 0.06 a | 0.99 ± 0.001 a | 8370 ± 232 a | 6348 ± 179 a |
| Rs + BC | 6.61 ± 0.13 a | 0.99 ± 0.003 a | 9623 ± 1370 a | 6999 ± 794 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Lu, Y.; Lin, W.; Tian, J.; Cai, K. Biochar Suppresses Bacterial Wilt of Tomato by Improving Soil Chemical Properties and Shifting Soil Microbial Community. Microorganisms 2019, 7, 676. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120676
Gao Y, Lu Y, Lin W, Tian J, Cai K. Biochar Suppresses Bacterial Wilt of Tomato by Improving Soil Chemical Properties and Shifting Soil Microbial Community. Microorganisms. 2019; 7(12):676. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120676
Chicago/Turabian StyleGao, Yang, Yang Lu, Weipeng Lin, Jihui Tian, and Kunzheng Cai. 2019. "Biochar Suppresses Bacterial Wilt of Tomato by Improving Soil Chemical Properties and Shifting Soil Microbial Community" Microorganisms 7, no. 12: 676. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120676