Cold Adapted Nitrosospira sp.: A Potential Crucial Contributor of Ammonia Oxidation in Cryosols of Permafrost-Affected Landscapes in Northeast Siberia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Investigation Site
2.2. Sampling and Investigation Strategy
2.3. Ammonia Oxidizing Potentials
2.4. Temperature Adaptation of Soil Incubations
2.5. DNA Extraction and amoA Analysis of Archaea and Bacteria
2.6. Analysis of 16S rRNA Amplicon Sequencing Diversity
2.7. Characterization of Enrichments of Nitrifying Microorganisms
2.8. Molecular Biologic Approaches
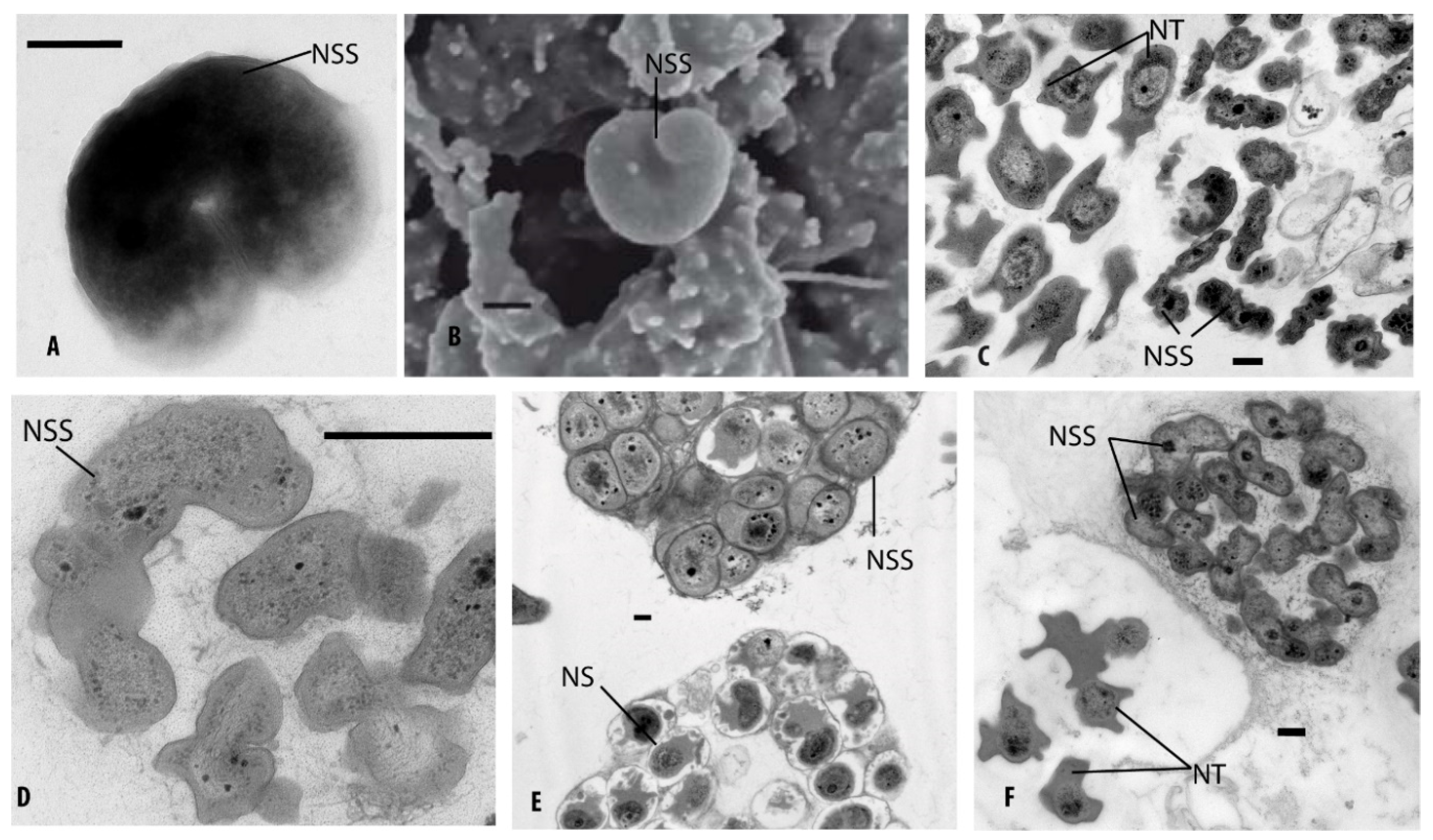
2.9. Electron Microscopy
2.10. Temperature Adaptation of Enrichment Cultures
3. Results
3.1. Potential Ammonia Oxidizing Activities
3.2. Temperature Adaptation of Potential Ammonia Oxidation in Soils
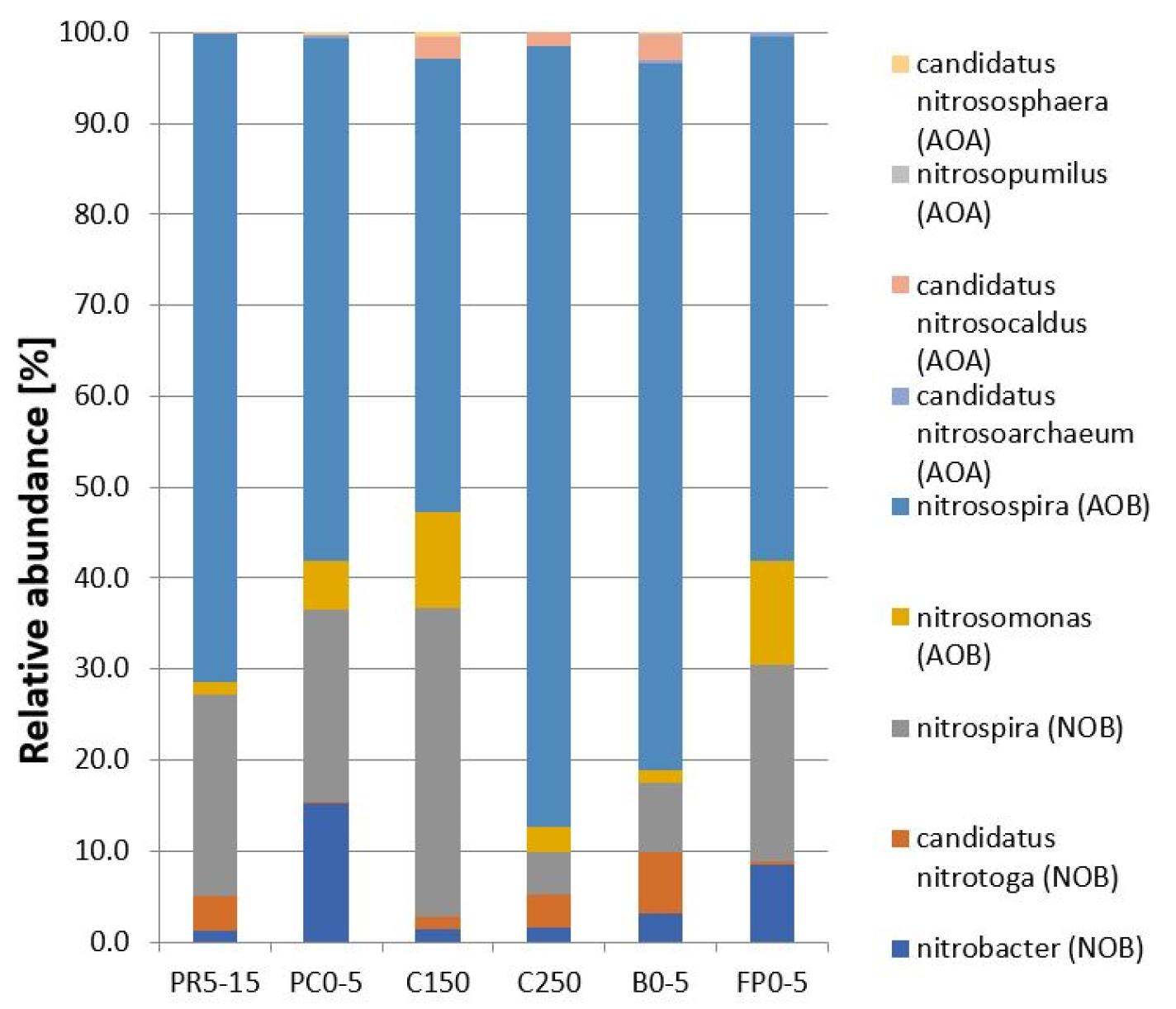
3.3. Microbial and Nitrifying Community Composition
3.4. Detection of amoA-genes in Soil Samples
3.5. AOA in Permafrost-Affected Soils
3.6. Identification of Ammonia-Oxidizing Microorganisms in Enrichment Cultures
3.7. Temperature Adaptation of Ammonia Oxidation in Enrichment Cultures
4. Discussion
4.1. Potential Ammonia Oxidizing Activities in Permafrost-Affected Soils
4.2. Occurrence of AOB and AOA in Permafrost-Affected Soils
4.3. Microbial Communities
4.4. Nitrosospira: the Primary Enriched AOB Representative
4.5. Which Nitrifiers are Adapted to the Extreme Environment of Low Temperature?
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Post, W.M.; Pasto, J.; Zinke, P.J.; Stangenberger, A.G. Global patterns of soil nitrogen storage. Nature 1985, 317, 613–616. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Schimel, J.P.; Trumbore, S.E.; Randerson, J.R. Controls over carbon storage and turnover in high-latitude soils. Glob. Chang. Biol. 2000, 6, 196–210. [Google Scholar] [CrossRef] [Green Version]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Kaiser, C.; Meyer, H.; Biasi, C.; Rusalimova, O.; Barsukov, P.; Richter, A. Storage and mineralization of carbon and nitrogen in soils of a frost-boil tundra ecosystem in siberia. Appl. Soil Ecol. 2005, 29, 173–183. [Google Scholar] [CrossRef]
- Holland, E.A.; Dentener, F.; Braswell, B.H.; Sulzman, J. Contemporary and pre-industrial global reactive nitrogen budgets. Biogeochemistry 1999, 46, 7–43. [Google Scholar] [CrossRef]
- Stewart, K.J.; Grogan, P.; Coxson, D.S.; Siciliano, S.D. Topography as a key factor driving atmospheric nitrogen exchanges in arctic terrestrial ecosystems. Soil Biol. Biochem. 2014, 70, 96–112. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R.; Schimel, D.S.; Fisher, H.; Howarth, R.W.; Hedin, L.O.; Perakis, S.S.; Latty, E.F.; Von Fischer, J.C.; Elseroad, A.; et al. Global patterns of terrestrial biological nitrogen (n2) fixation in natural ecosystems. Glob. Biogeochem. Cycles 1999, 13, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Hobara, S.; McCalley, C.; Koba, K.; Giblin, A.; Weiss, M.; Gettel, G.; Shaver, G. Nitrogen fixation in surface soils and vegetation in an arctic tundra watershed: A key source of atmospheric nitrogen. Arct. Antarct. Alp. Res. 2006, 38, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Mack, M.C.; Schuur, E.A.G.; Bret-Harte, M.S.; Shaver, G.R.; Chapin, F.S.I. Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 2004, 431, 440–443. [Google Scholar] [CrossRef]
- Meyer, H.; Kaiser, C.; Biasi, C.; Hämmerle, R.; Rusalimova, O.; Lashchinsky, N.; Baranyi, C.; Daims, H.; Barsukov, P.; Richter, A. Soil carbon and nitrogen dynamics along a latitudinal transect in western siberia, russia. Biogeochemistry 2006, 81, 239–252. [Google Scholar] [CrossRef]
- Rodionov, A.; Flessa, H.; Grabe, M.; Kazansky, O.A.; Shibistova, O.; Guggenberger, G. Organic carbon and total nitrogen variability in permafrost-affected soils in a forest tundra ecotone. Eur. J. Soil Sci. 2007, 58, 1260–1272. [Google Scholar] [CrossRef]
- Koyama, A.; Wallenstein, M.D.; Simpson, R.T.; Moore, J.C. Soil bacterial community composition altered by increased nutrient availability in arctic tundra soils. Front. Microbiol. 2014, 5, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, A.M.; Epstein, H.E.; Ping, C.-L.; Walker, D.A. Soil nitrogen transformations associated with small patterned-ground features along a north american arctic transect. Permafr. Periglac. Process. 2012, 23, 196–206. [Google Scholar] [CrossRef]
- Biasi, C.; Wanek, W.; Rusalimova, O.; Kaiser, C.; Meyer, H.; Barsukov, P.; Richter, A. Microtopography and plant-cover controls on nitrogen dynamics in hummock tundra ecosystems in siberia. Arct. Antarct. Alp. Res. 2005, 37, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.; Lamprecht, R.E.; Marushchak, M.E.; Lind, S.E.; Novakovskiy, A.; Aurela, M.; Martikainen, P.J.; Biasi, C. Warming of subarctic tundra increases emissions of all three important greenhouse gases—Carbon dioxide, methane, and nitrous oxide. Glob. Chang. Biol. 2017, 23, 3121–3138. [Google Scholar] [CrossRef]
- Voigt, C.; Marushchak, M.E.; Lamprecht, R.E.; Jackowicz-Korczyński, M.; Lindgren, A.; Mastepanov, M.; Granlund, L.; Christensen, T.R.; Tahvanainen, T.; Martikainen, P.J.; et al. Increased nitrous oxide emissions from arctic peatlands after permafrost thaw. Proc. Natl. Acad. Sci. USA 2017, 114, 6238–6243. [Google Scholar] [CrossRef] [Green Version]
- Siljanen, H.M.; Alves, R.J.; Ronkainen, J.G.; Lamprecht, R.E.; Bhattarai, H.R.; Bagnoud, A.; Marushchak, M.E.; Martikainen, P.J.; Schleper, C.; Biasi, C. Archaeal nitrification is a key driver of high nitrous oxide emissions from arctic peatlands. Soil Biol. Biochem. 2019, 137, 107539. [Google Scholar] [CrossRef]
- Harms, T.K.; Jones, J.B. Thaw depth determines reaction and transport of inorganic nitrogen in valley bottom permafrost soils. Glob. Chang. Biol. 2012, 18, 2958–2968. [Google Scholar] [CrossRef]
- Malard, L.A.; Pearce, D.A. Microbial diversity and biogeography in arctic soils. Environ. Microbiol. Rep. 2018, 10, 611–625. [Google Scholar] [CrossRef]
- Chapin, D.M. Nitrogen mineralization, nitrification, and denitrification in a high arctic lowland ecosystem, devon island, N.W.T., Canada. Arct. Alp. Res. 1996, 28, 85–92. [Google Scholar] [CrossRef]
- Petersen, D.G.; Blazewicz, S.J.; Firestone, M.; Herman, D.J.; Turetsky, M.; Waldrop, M. Abundance of microbial genes associated with nitrogen cycling as indices of biogeochemical process rates across a vegetation gradient in alaska. Environ. Microbiol. 2012, 14, 993–1008. [Google Scholar] [CrossRef] [PubMed]
- Nadelhoffer, K.J.; Giblin, A.E.; Shaver, G.R.; Laundre, J.A. Effects of termperature and substrate quality on element mineralization in six arctic soils. Ecology 1991, 72, 242–253. [Google Scholar] [CrossRef]
- Banerjee, S.; Siciliano, S.D. Factors driving potential ammonia oxidation in canadian arctic ecosystems: Does spatial scale matter? Appl. Environ. Microbiol. 2012, 78, 346–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Shimomura, Y.; Morimoto, S.; Uchida, M.; Nakatsubo, T.; Hayatsu, M. Characteristics of ammonia oxidation potentials and ammonia oxidizers in mineral soil under salix polaris–moss vegetation in ny-ålesund, svalbard. Polar Biol. 2016, 39, 725–741. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.J.E.; Wanek, W.; Zappe, A.; Richter, A.; Svenning, M.M.; Schleper, C.; Urich, T. Nitrification rates in arctic soils are associated with functionally distinct populations of ammonia-oxidizing archaea. ISME J 2013, 7, 1620. [Google Scholar] [CrossRef] [Green Version]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A. Complete nitrification by nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef]
- van Kessel, M.A.H.J.; Speth, D.R.; Albertsen, M.; Nielsen, P.H.; Op den Camp, H.J.M.; Kartal, B.; Jetten, M.S.M.; Lücker, S. Complete nitrification by a single microorganism. Nature 2015, 528, 555. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.Q.; Bakken, L.R. Comparison of nitrosospira strains isolated from terrestrial environments. Fems Microbiol. Ecol. 1999, 30, 171–186. [Google Scholar] [CrossRef]
- Koops, H.P.; Pommerening-Röser, A. Distribution and ecophysiology of the nitrifying bacteria emphasizing cultured species. Fems Microbiol. Ecol. 2001, 37, 1–9. [Google Scholar] [CrossRef]
- Avrahami, S.; Conrad, R. Cold-temperate climate: A factor for selection of ammonia oxidizers in upland soil? Can. J. Microbiol. 2005, 51, 709–714. [Google Scholar] [CrossRef]
- Avrahami, S.; Bohannan, B.J.M. Response of nitrosospira sp. Strain af-like ammonia oxidizers to changes in temperature, soil moisture content, and fertilizer concentration. Appl. Environ. Microbiol. 2007, 73, 1166–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martikainen, P.J.; Nurmiaho-Lassila, E.-L. Nitrosospira, an important ammonium-oxidizing bacterium in fertilized coniferous forest soil. Can. J. Microbiol. 1985, 31, 190–197. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, J.; Su, J.; Jia, Z.; Shi, X.; Wright, A.L.; Zhu-Barker, X.; Jiang, X. Neutrophilic bacteria are responsible for autotrophic ammonia oxidation in an acidic forest soil. Soil Biol. Biochem. 2018, 119, 83–89. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; He, Y.; Liu, H.; Dumont, M.G.; Brookes, P.C.; Xu, J. Nitrosospira cluster 3-like bacterial ammonia oxidizers and nitrospira-like nitrite oxidizers dominate nitrification activity in acidic terrace paddy soils. Soil Biol. Biochem. 2019, 131, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Urakawa, H.; Garcia, J.C.; Nielsen, J.L.; Le, V.Q.; Kozlowski, J.A.; Stein, L.Y.; Lim, C.K.; Pommerening-Röser, A.; Martens-Habbena, W.; Stahl, D.A. Nitrosospira lacus sp. Nov., a psychrotolerant, ammonia-oxidizing bacterium from sandy lake sediment. Int. J. Syst. Evol. Microbiol. 2015, 65, 242–250. [Google Scholar] [CrossRef]
- Long, X.; Chen, C.; Xu, Z.; Oren, R.; He, J.-Z. Abundance and community structure of ammonia-oxidizing bacteria and archaea in a temperate forest ecosystem under ten-years elevated CO2. Soil Biol. Biochem. 2012, 46, 163–171. [Google Scholar] [CrossRef]
- Pester, M.; Rattei, T.; Flechl, S.; Grongroft, A.; Richter, A.; Overmann, J.; Reinhold-Hurek, B.; Loy, A.; Wagner, M. Amoa-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoa genes from soils of four different geographic regions. Environ. Microbiol. 2012, 14, 525–539. [Google Scholar] [CrossRef] [Green Version]
- Brochier-Armanet, C.; Boussau, B.; Gribaldo, S.; Forterre, P. Mesophilic crenarchaeota: Proposel for a third archaeal phylum, the thaumarchaeota. Nat. Rev. 2008, 6, 245–252. [Google Scholar] [CrossRef]
- Sauder, L.A.; Albertsen, M.; Engel, K.; Schwarz, J.; Nielsen, P.H.; Wagner, M.; Neufeld, J.D. Cultivation and characterization of candidatus nitrosocosmicus exaquare, an ammonia-oxidizing archaeon from a municipal wastewater treatment system. ISME J. 2017, 11, 1142. [Google Scholar] [CrossRef]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.l. The influence of soil ph on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef]
- Schleper, C. Ammonia oxidation: Different niches for bacteria and archaea? ISME J. 2010, 4, 1092–1094. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.; Tang, J.; Riley, W.J.; Brodie, E.L. Trait-based representation of biological nitrification: Model development, testing, and predicted community composition. Front. Microbiol. 2012, 3, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makhalanyane, T.P.; Van Goethem, M.W.; Cowan, D.A. Microbial diversity and functional capacity in polar soils. Curr. Opin. Biotechnol. 2016, 38, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yergeau, E.; Hogues, H.; Whyte, L.G.; Greer, C.W. The functional potential of high arctic permafrost revealed by metagenomic sequencing, qpcr and microarray analyses. ISME J 2010, 4, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.J.E.; Kerou, M.; Zappe, A.; Bittner, R.; Abby, S.S.; Schmidt, H.; Pfeifer, K.; Schleper, C. Ammonia oxidation by the arctic terrestrial thaumarchaeote ca. Nitrosocosmicus arcticus is stimulated by increasing temperatures. Front. Microbiol. 2019, 10, 1571. [Google Scholar] [PubMed] [Green Version]
- Xia, F.; Wang, J.-G.; Zhu, T.; Zou, B.; Rhee, S.-K.; Quan, Z.-X. Ubiquity and diversity of complete ammonia oxidizers (comammox). Appl. Environ. Microbiol. 2018, 84, e01390-18. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.-W.; He, J.-Z. Comammox—A newly discovered nitrification process in the terrestrial nitrogen cycle. J. Soils Sediments 2017, 17, 2709–2717. [Google Scholar] [CrossRef]
- Aigle, A.; Prosser, J.I.; Gubry-Rangin, C. The application of high-throughput sequencing technology to analysis of amoa phylogeny and environmental niche specialisation of terrestrial bacterial ammonia-oxidisers. Environ. Microbiome 2019, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Pithan, F.; Mauritsen, T. Arctic amplification dominated by temperature feedbacks in contemporary climate models. Nat. Geosci. 2014, 7, 181. [Google Scholar] [CrossRef]
- Schwamborn, G.; Rachold, V.; Grigoriev, M.N. Late quaternary sedimentation history of the lena delta. Quat. Int. 2002, 89, 119–134. [Google Scholar] [CrossRef] [Green Version]
- Roshydromet (Russian Federal Service for Hydrometeorology and Environment Monitoring). Weather Information for Tiksi; World Meteorological Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Boike, J.; Kattenstroth, B.; Abramova, K.; Bornemann, N.; Chetverova, A.; Fedorova, I.; Fröb, K.; Grigoriev, M.; Grüber, M.; Kutzbach, L. Baseline characteristics of climate, permafrost and land cover from a new permafrost observatory in the lena river delta, siberia (1998–2011). Biogeosciences 2013, 10, 2105–2128. [Google Scholar] [CrossRef] [Green Version]
- Sanders, T.; Fiencke, C.; Pfeiffer, E.M. Small-scale variability of dissolved inorganic nitrogen (din), c/n rations and ammonia oxidizing capacities in various permafrost affected soils of samoylov island, lena river delta, northeast siberia. Polarforschung 2010, 80, 23–35. [Google Scholar]
- Staff, S.S. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Zimmermann, U. Methanoxidierende Bakteriengemeinschaften in Böden und Sedimenten des sibirischen Permafrostes. Ph.D. Thesis, Universität Hamburg, Hamburg, Germany, 2006. [Google Scholar]
- Bodenbeschaffenheit—Bestimmung der potentiellen Nitrifizierung—Schnellverfahren mittels Ammoniumoxidation; DIN15685; Deutsches Institut für Normung e.V.: Berlin, Germany, 2001.
- Rotthauwe, J.-H.; Witzel, K.-P.; Liesack, W. The ammonia monooxygenase structural gene amoa as a funvtional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [PubMed]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [Green Version]
- Rotthauwe, J.-H.; de Boer, W.; Liesack, W. Comparative analysis of gen sequences encoding ammonia monooxygenase of nitrosospira sp. Ahb 1 and nitrosolobus multiformis c-71. Fems Microbiol. Lett. 1995, 133, 131–135. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [Green Version]
- Dowd, S.E.; Sun, Y.; Wolcott, R.D.; Domingo, A.; Carroll, J.A. Bacterial tag–encoded flx amplicon pyrosequencing (btefap) for microbiome studies: Bacterial diversity in the ileum of newly weaned salmonella-infected pigs. Foodborne Pathog. Dis. 2008, 5, 459–472. [Google Scholar] [CrossRef]
- DeSantis, T.Z., Jr.; Hugenholtz, P.; Keller, K.; Brodie, E.L.; Larsen, N.; Piceno, Y.M.; Phan, R.; Andersen, G.L. Nast: A multiple sequence alignment server for comparative analysis of 16s rrna genes. Nucleic Acids Res. 2006, 34, W394–W399. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. Past: Paleontological statistics softwarepackage for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Koops, H.-P.; Purkhold, U.; Pommerening-Röser, A.; Timmermann, G.; Wagner, M. The lithoautotrophic ammonia-oxidizing bacteria. In The Prokaryotes; Springer: Berlin, Germany, 2006; pp. 778–811. [Google Scholar]
- Schmidt, E.L.; Belser, L.W. Nitrifying bacteria. In Methods of Soil Analysis Properties; Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 1027–1042. [Google Scholar]
- Spieck, E.; Hartwig, C.; McCormack, I.; Maixner, F.; Wagner, M.; Lipski, A.; Daims, H. Selective enrichment and molecular characterization of a previously uncultured nitrospira-like bacterium from activated sludge. Environ. Microbiol. 2006, 8, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Steinmüller, W.; Bock, E. Growth of nitrobacter in the presence of organic matter. Arch. Microbiol. 1976, 108, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Alawi, M. Diversität Nitrit oxidierender Bakterien in Böden des nordsibirischen Permafrostes und Sedimenten der Laptew-See. Ph.D. Thesis, Universität Hamburg, Hamburg, Germany, 2007. [Google Scholar]
- Sanders, T. Vergleichende Untersuchungen kälteliebender nitrifizierender Bakterien aus Permafrostböden im Lena Delta, Sibirien. Diploma Thesis, Universität Hamburg, Hamburg, Germany, 2006. [Google Scholar]
- Sanders, T. Charakteriserung Ammoniak oxidierenden Mikroorganismen in Böden kalter und gemäßigter Klimate und ihre Bedeutung für den globalen Stickstoffkreislauf. Ph.D. Thesis, Universität Hamburg, Hamburg, Germany, 2011. [Google Scholar]
- Lane, D.J. 16s/23s rrna sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- DeLong, E.F. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. Mega4: Molecular evolutionary genetics analysis (mega) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Hatzenpichler, R.; Lebedeva, E.V.; Spieck, E.; Stoecker, K.; Richter, A.; Daims, H.; Wagner, M. A moderately thermophilic ammonia-oxidizing crenarchaeote from a hot spring. Proc. Natl. Acad. Sci. USA 2008, 105, 2134–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tourna, M.; Stieglmeier, M.; Spang, A.; Könneke, M.; Schintlmeister, A.; Urich, T.; Engel, M.; Schloter, M.; Wagner, M.; Richter, A.; et al. Nitrososphaera viennensis, an ammonia oxidizing archaeon from soil. Proc. Natl. Acad. Sci. USA 2011, 108, 8420–8425. [Google Scholar] [CrossRef] [Green Version]
- Purkhold, U.; Wagner, M.; Timmermann, G.; Pommerening-Röser, A.; Koops, H.-P. 16s rrna and amoa-based phylogeny of 12 novel betaproteobacterial ammonia-oxidizing isolates: Extension of the dataset and proposel of a new leneage within the nitrosomonades. Int. J. Syst. Evol. Microbiol. 2003, 53, 1485–1494. [Google Scholar] [CrossRef]
- Morita, R.Y. Low-teperature environments. Encycl. Microbiol. 2000, 3, 93–98. [Google Scholar]
- Wild, B.; Schnecker, J.; Bárta, J.; Čapek, P.; Guggenberger, G.; Hofhansl, F.; Kaiser, C.; Lashchinsky, N.; Mikutta, R.; Mooshammer, M. Nitrogen dynamics in turbic cryosols from siberia and greenland. Soil Biol. Biochem. 2013, 67, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Yeom, J.; Kim, J.; Han, J.; Lim, H.S.; Park, H.; Hyun, S.; Park, W. Change in gene abundance in the nitrogen biogeochemical cycle with temperature and nitrogen addition in antarctic soils. Res. Microbiol. 2011, 162, 1018–1026. [Google Scholar] [CrossRef]
- Boyd, E.S.; Lange, R.K.; Mitchell, A.C.; Havig, J.R.; Hamilton, T.L.; Lafrenière, M.J.; Shock, E.L.; Peters, J.W.; Skidmore, M. Diversity, abundance, and potential activity of nitrifying and nitrate-reducing microbial assemblages in a subglacial ecosystem. Appl. Environ. Microbiol. 2011, 77, 4778–4787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubry-Rangin, C.; Novotnik, B.; Mandič-Mulec, I.; Nicol, G.W.; Prosser, J.I. Temperature responses of soil ammonia-oxidising archaea depend on ph. Soil Biol. Biochem. 2017, 106, 61–68. [Google Scholar] [CrossRef]
- Jansson, J.K.; Taş, N. The microbial ecology of permafrost. Nat. Rev. Microbiol. 2014, 12, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Liebner, S.; Harder, J.; Wagner, D. Bacterial diversity and community structure in polygonal tundra soils from samoylov island, lena delta, siberia. Int. Microbiol. 2008, 11, 195–202. [Google Scholar] [PubMed]
- Frey, B.; Stierli, B.; Rime, T.; Hartmann, M.; Phillips, M.; Hajdas, I.; Widmer, F. Microbial diversity in european alpine permafrost and active layers. Fems Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [Green Version]
- Lebedeva, E.V.; Alawi, M.; Fiencke, C.; Namsaraev, B.; Bock, E.; Spieck, E. Moderately thermophilic bacteria from a hot spring of the baikal rift zone. Fems Microbiol. Ecol. 2005, 54, 297–306. [Google Scholar] [CrossRef] [Green Version]
- De Boer, W.; Kowalchuk, G.A. Nitrification in acid soils: Micro-organisms and mechanisms. Soil Biol. Biochem. 2001, 33, 853–866. [Google Scholar] [CrossRef]
- Avrahami, S.; Liesack, W.; Conrad, R. Effects of temperature and fertilizer on activity and community structure of soil ammonia oxidizers. Environ. Microbiol. 2003, 5, 691–705. [Google Scholar] [CrossRef]
- Miazga-Rodriguez, M.; Han, S.; Yakiwchuk, B.; Wei, K.; English, C.; Bourn, S.; Bohnet, S.; Stein, L.Y. Enhancing nitrification at low temperature with zeolite in a mining operations retention pond. Front. Microbiol. 2012, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Karkman, A.; Mattila, K.; Tamminen, M.; Virta, M. Cold temperature decreases bacterial species richness in nitrogen-removing bioreactors treating inorganic mine waters. Biotechnol. Bioeng. 2011, 108, 2876–2883. [Google Scholar] [CrossRef]
- Jones, R.D.; Morita, R.Y.; Koops, H.-P.; Watson, S.W. A new marine ammonium-oxidizing bacterium, nitrosomonas cryotolerans sp. Nov. Can. J. Microbiol. 1988, 34, 1122–1128. [Google Scholar] [CrossRef]
- Jones, R.D.; Morita, R.Y. Low-temperature growth and whole-cell kinetics of a marine ammonium oxidizer. Mar. Ecol. Proz. Ser. 1985, 21, 239–243. [Google Scholar] [CrossRef]
- Thamdrup, B.; Fleischer, S. Temperature dependence of oxygen respiration, nitrogen mineralization, and nitrification in arctic sediments. Aquat. Microb. Ecol. 1998, 15, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, K.; Kemp, W.M. Nitrification in estuarine and coastal marine sediments. In Nitrogen Cycling in Coastal Marine Environments; Blackburn, T.H., Sorensen, J., Eds.; John Wiley and Sons: New York, NY, USA, 1988; pp. 207–249. [Google Scholar]
- Hollibaugh, J.T.; Bano, N.; Ducklow, H.W. Widerspread distribution in polar oceans of a 16 s rrna gene seqeunce with affinity to nitrosospira-like ammonia-oxidizing bacteria. Appl. Environ. Microbiol. 2002, 68, 1478–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuchter, C.; Abbas, B.; Coolen, M.J.; Herfort, L.; van Bleijswijk, J.; Timmers, P.; Strous, M.; Teira, E.; Herndl, G.J.; Middelburg, J.J.; et al. Archaeal nitrification in the ocean. Proc. Natl. Acad. Sci. USA 2006, 103, 12317–12322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.E.; Giguere, A.T.; Zoebelein, C.M.; Myrold, D.D.; Bottomley, P.J. Modeling of soil nitrification responses to temperature reveals thermodynamic differences between ammonia-oxidizing activity of archaea and bacteria. ISME J. 2017, 11, 896–908. [Google Scholar] [CrossRef]
- Boike, J.; Wille, C.; Abnizova, A. Climatology and summer energy and water balance of polygonal tundra in the lena river delta, siberia. J. Geophys. Res. 2008, 113, G03025. [Google Scholar] [CrossRef] [Green Version]
- Alawi, M.; Lipski, A.; Sanders, T.; Pfeiffer, E.M.; Spieck, E. Cultivation of a novel cold-adapted nitrite oxidizing betaproteobacterium from the siberian arctic. ISME J. 2007, 1, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Nowka, B.; Daims, H.; Spieck, E. Comparison of oxidation kinetics of nitrite-oxidizing bacteria: Nitrite availability as a key factor in niche differentiation. Appl. Environ. Microbiol. 2015, 81, 745–753. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Sample name | Depth (cm) | Organic matter (%) | pH (H2O) | Corg (%) | N (%) | CN | NH4+ (g g–1 dw) | N02- (g g–1 dw) | NO3- (g g–1 dw) |
|---|---|---|---|---|---|---|---|---|---|---|
| “River terrace” | ||||||||||
| Polygon rim (IS1) | PR0–5 | 0–5 | 17.1 | 6.6 | 10.0 | 0.3 | 37 | 0.3 | 0.02 | bdl |
| Polygon rim (IS1) | PR5–15 | 5–15 | 5.3 | 6.6 | 3.1 | 0.2 | 18 | 0.3 | bdl | bdl |
| Polygon rim (IS1) | PR15–25 | 15–25 | 4.9 | 6.2 | 2.9 | 0.2 | 16 | 0.1 | bdl | bdl |
| Polygon center (IS1) | PC0–5 | 0–5 | 37.1 | 6.3 | 18.6 | 0.6 | 33 | 0.5 | 0.01 | bdl |
| Polygon center (IS1) | PC5–15 | 5–15 | 36.1 | 6.1 | 18.0 | 0.5 | 37 | 0.2 | 0.01 | bdl |
| Polygon center (IS1) | PC15–25 | 15–25 | 13.0 | 6.0 | 6.5 | 0.2 | 37 | 0.6 | bdl | bdl |
| Dry river terrace (IS2) | RT0–5 | 0–5 | 1.3 | 6.4 | 0.8 | 0.1 | 17 | 0.3 | 0.01 | bdl |
| Dry river terrace (IS2) | RT5–15 | 5–15 | 1.1 | 6.4 | 0.7 | 0.1 | 14 | 0.0 | 0.01 | bdl |
| Cliff* (IS2) | C150 | 150 | 1.0 | 6.8 | 0.6 | 0.0 | 15 | 5.5 | 0.03 | 0.1 |
| Cliff* (IS2) | C250 | 250 | 4.9 | 6.9 | 2.9 | 0.2 | 16 | 10.0 | 0.02 | bdl |
| “Modern floodplain” | ||||||||||
| Beach* (IS4) | B0–5 | 0–5 | 3.9 | 6.9 | 2.3 | 0.2 | 15 | 9.4 | 0.04 | bdl |
| Beach* (IS4) | B10–20 | 10–20 | 0.3 | 7.0 | 0.2 | 0.0 | 12 | 0.3 | 0.02 | 0.2 |
| Floodplain (IS5) | FP0–5 | 0–5 | 1.6 | 7.3 | 0.9 | 0.1 | 14 | 0.1 | bdl | 0.1 |
| Floodplain (IS5) | FP5–15 | 5–15 | 1.8 | 7.3 | 1.0 | 0.1 | 16 | 0.2 | bdl | 0.1 |
| Sampling Site | Ammonia Oxidizing Capacities (ng N g–1 h–1) | Detection of amoA by PCR | Microbial Diversity | ||||
|---|---|---|---|---|---|---|---|
| Bacterial 2 | Archaeal 2 | Reads | Nitrifiers | ||||
| Total 1 | Plus Streptomycin | Total | % | ||||
| “River terrace” | |||||||
| Polygon rim (IS1) | PR0–5 | bdl | bdl | + | – | nd | nd |
| Polygon rim (IS1) | PR5–15 | 114 ± 8 | bdl | + | – | 87,419 | 6.25 |
| Polygon rim (IS1) | PR15–25 | bdl | bdl | + | – | nd | nd |
| Polygon center (IS1) | PC0–5 | 33 | bdl | – | – | nd | nd |
| Polygon center (IS1) | PC5–15 | bdl | bdl | – | – | nd | nd |
| Polygon center (IS1) | PC15–25 | bdl | bdl | – | – | nd | nd |
| Dry river terrace (IS2) | RT0–5 | 103 | bdl | + | + | 107,960 | 1.35 |
| Dry river terrace (IS2) | RT5–15 | 78 ± 8 | 3 | + | + | nd | nd |
| Cliff3(IS2) | C150 | 197 ± 11 | bdl | + | – | 79,183 | 0.81 |
| Cliff3(IS2) | C250 | 571 ± 113 | bdl | nd | nd | 71,803 | 1.76 |
| “Modern Floodplain” | |||||||
| Beach3 (IS4) | B0–5 | 542 ± 13 | 197 | + | + | 84,338 | 4.77 |
| Beach3 (IS4) | B10–20 | 102 ± 8 | bdl | nd | nd | nd | nd |
| Floodplain (IS5) | FP0–5 | 208 ± 3 | 6 | + | + | 103,120 | 0.66 |
| Floodplain (IS5) | FP5–15 | 225 ± 6 | 2 | + | + | nd | nd |
| Enrichment Culture | Sampling Site | Depth (cm) | Year Enriched | Incubation Temperature | DGGE Analyses of 16S rRNA or EM images | Detection of amoA by PCR | ||
|---|---|---|---|---|---|---|---|---|
| AOB | NOB | Bacteria | Archaea | |||||
| 3304/3306 | Samoylov Cliff (IS3) | 240/350 | 2004 | 4 °C | Nitrosospira | Nitrotoga* | + | (+) |
| W0–5 | Polygon rim (IS1) | 0–5 | 2004 | 4 °C | Nitrosospira | - | + | – |
| W12–17 | 12–17 | 2004 | 10 °C | Nitrosospira | - | + | – | |
| 11 | Polygon slope (IS1) | 12–17 | 2005 | 28 °C | Nitrosospira | Nitrospira | + | – |
| B0–5 | Beach (IS4) | 0–5 | 2008 | 4 °C | Nitrosospira* | Nitrotoga | + | (+) |
| CT0840 | 0–5 | 2008 | 18/28 °C | - | - | + | + | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanders, T.; Fiencke, C.; Hüpeden, J.; Pfeiffer, E.M.; Spieck, E. Cold Adapted Nitrosospira sp.: A Potential Crucial Contributor of Ammonia Oxidation in Cryosols of Permafrost-Affected Landscapes in Northeast Siberia. Microorganisms 2019, 7, 699. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120699
Sanders T, Fiencke C, Hüpeden J, Pfeiffer EM, Spieck E. Cold Adapted Nitrosospira sp.: A Potential Crucial Contributor of Ammonia Oxidation in Cryosols of Permafrost-Affected Landscapes in Northeast Siberia. Microorganisms. 2019; 7(12):699. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120699
Chicago/Turabian StyleSanders, Tina, Claudia Fiencke, Jennifer Hüpeden, Eva Maria Pfeiffer, and Eva Spieck. 2019. "Cold Adapted Nitrosospira sp.: A Potential Crucial Contributor of Ammonia Oxidation in Cryosols of Permafrost-Affected Landscapes in Northeast Siberia" Microorganisms 7, no. 12: 699. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7120699