Deciphering the Symbiotic Significance of Quorum Sensing Systems of Sinorhizobium fredii HH103

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Well Diffusion Assay and Thin Layer Chromatography Analysis
2.3. HPLC and Mass Spectrometry Analysis
2.4. EPS Production and Analysis of Lipopolysaccharide (LPS) and K-Antigens Polysaccharide (KPS)
2.5. Motility Assays
2.6. Biofilm Formation Assays
2.7. Nodulation Assays
2.8. Protein Alignment
2.9. RNA-seq Data Accession Number and Gene Transcript Assignment
2.10. Calculation of pSym Conjugation Frequency
2.11. Quantification of Plasmid Copy Number
3. Results
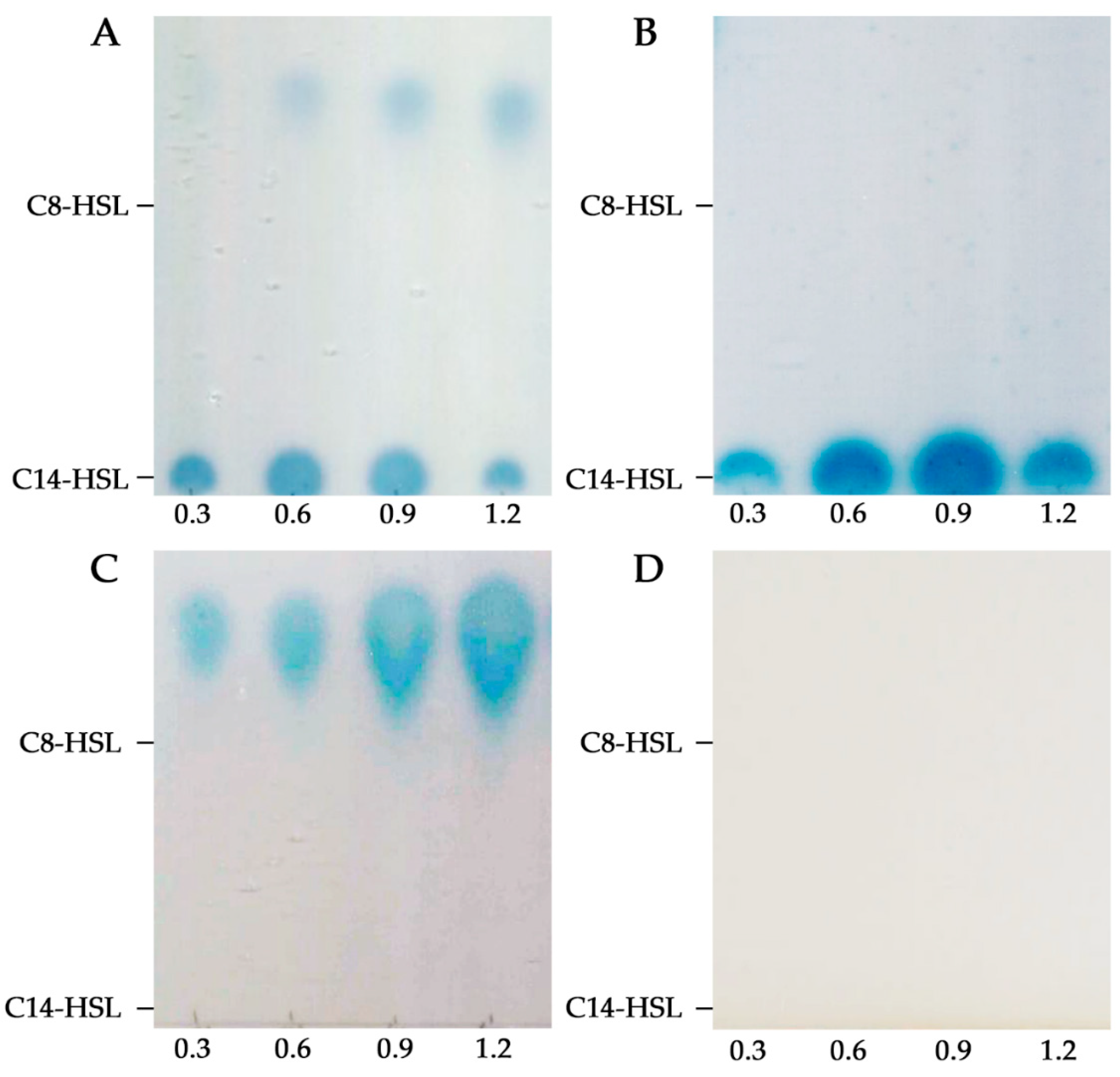
3.1. S. fredii HH103 Produces Short- and Long-Chain AHLs at not Physiological Concentrations by Means of TraI and SinI, Respectively
3.2. S. fredii HH103 AHLs Do Not Regulate Symbiotically Important Traits and Have No Influence on the Symbiotic Performance with G. max and G. uralensis
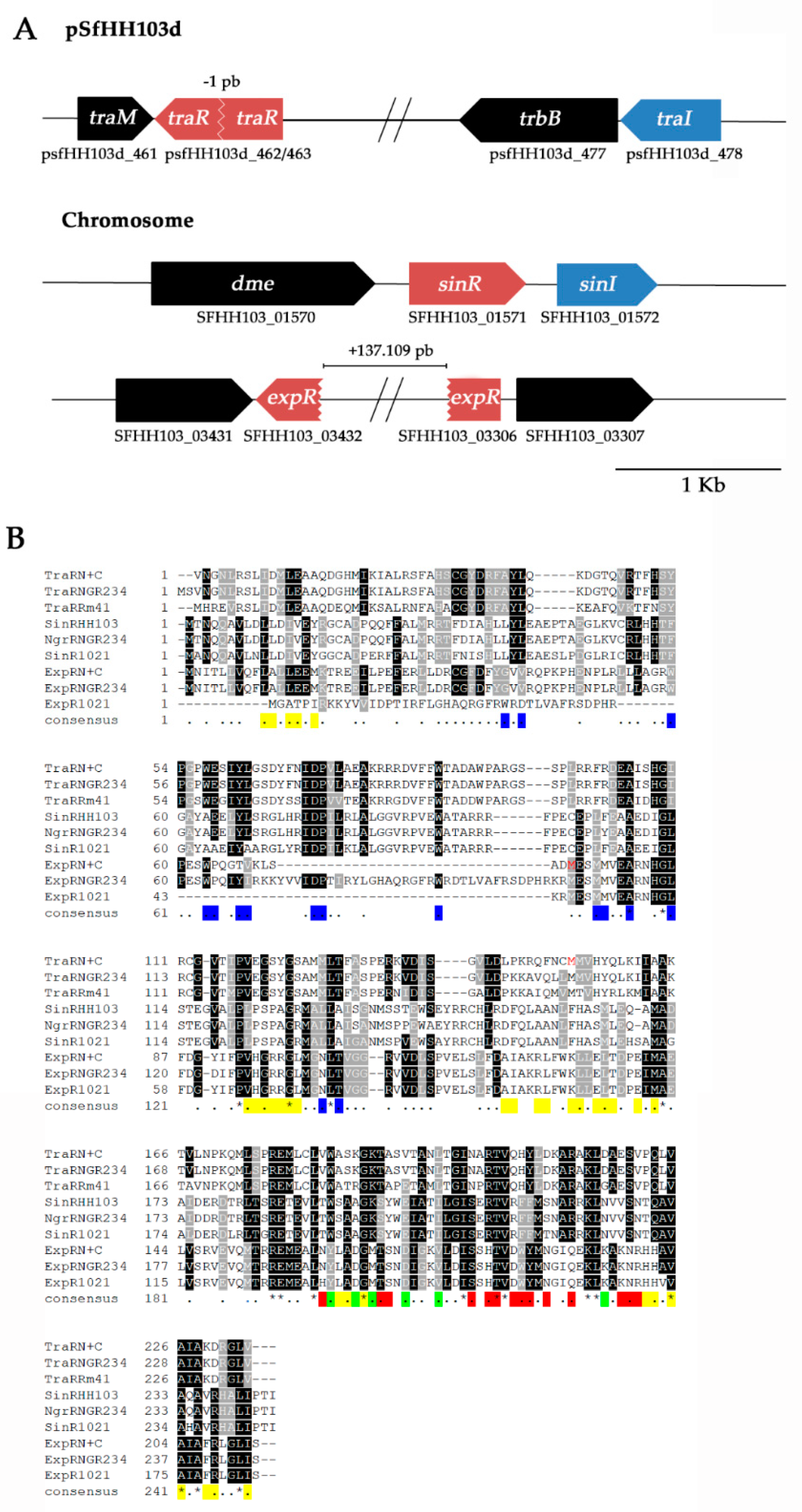
3.3. The traR and expR Genes of S. fredii HH103 Are Divided in Two ORFs
3.4. The Terminal Parts of traR and expR Genes Code for Proteins That Conserve the DNA-binding Motives and the RNA Polymerase-Recruitment Residues
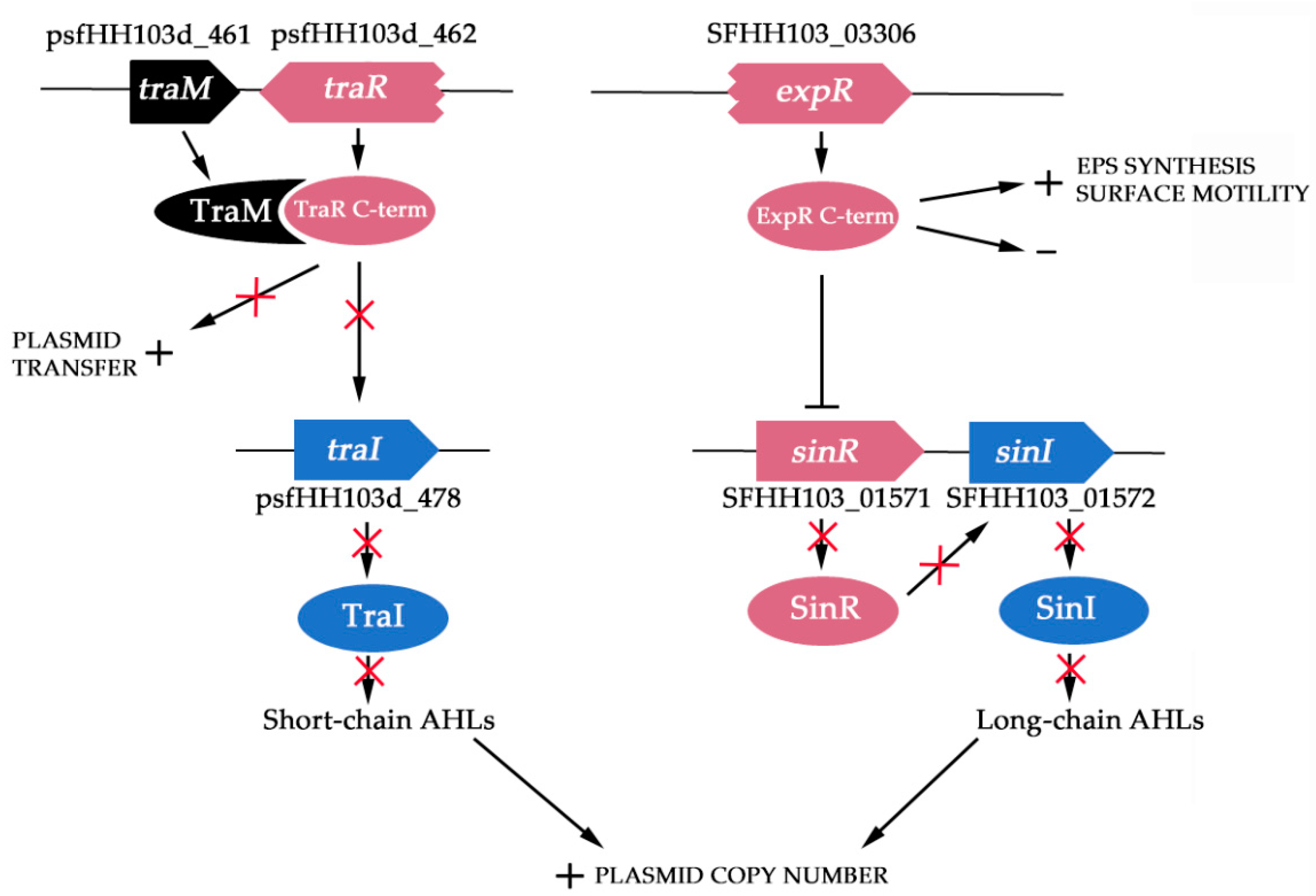
3.5. EPS Production Genes But Not Motility or Plasmid Transfer Genes Are Highly Expressed at High Population Densities
3.6. S. fredii HH103 sinI and/or traI Mutants Show Similar Conjugation Rates of Their pSym Than the Wild-Type Strain
3.7. The Symbiotic Plasmid of S. fredii HH103 Is Present in about 3 Copies per Cell with Regard to the Chromosome
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gage, D.J. Infection and invasion of roots by symbiotic, nitrogen-fixing rhizobia during nodulation of temperate legumes. Microbiol. Mol. Biol. Rev. 2004, 68, 280–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How rhizobial symbionts invade plants: The Sinorhizobium-Medicago model. Nat. Rev. Microbiol. 2007, 5, 619–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deakin, W.J.; Broughton, W.J. Symbiotic use of pathogenic strategies, rhizobial protein secretion systems. Nat. Rev. Microbiol. 2009, 7, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Spaink, H.P. Root nodulation and infection factors produced by rhizobial bacteria. Annu. Rev. Microbiol. 2000, 54, 257–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.S.; Sadowsky, M.J. Secretion systems and signal exchange between nitrogen-fixing rhizobia and legumes. Front. Plant Sci. 2015, 6, 491. [Google Scholar] [CrossRef] [Green Version]
- Kondorosi, E.; Gyuris, J.; Schmidt, J.; John, M.; Duda, E.; Hoffmann, B.; Schell, J.; Kondorosi, A. Positive and negative control of nod gene expression in Rhizobium meliloti is required for optimal nodulation. EMBO J. 1989, 8, 1331–1340. [Google Scholar] [CrossRef]
- Oldroyd, G.E. Speak, friend, and enter: Signaling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Broughton, W.J.; Perret, X. Genealogy of legume- Rhizobium symbioses. Curr. Opin. Plant Biol. 1999, 2, 305–311. [Google Scholar] [CrossRef]
- Krysciak, D.; Orbegoso, M.R.; Schmeisser, C.; Streit, W.R. Molecular keys to broad host range in Sinorhizobium fredii NGR234, USDA257, and HH103. In Biological Nitrogen Fixation; de Bruijn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 325–336. [Google Scholar]
- Pueppke, S.G.; Broughton, W.J. Rhizobium sp. Strain NGR234 and R. fredii USDA257 share exceptionally broad, nested host ranges. Mol. Plant Microbe Interact. 1999, 12, 293–318. [Google Scholar] [CrossRef] [Green Version]
- Schmeisser, C.; Liesegang, H.; Krysciak, D.; Bakkou, N.; Le Quéré, A.; Wollherr, A.; Heinemeyer, I.; Morgenstern, B.; Pommerening-Röser, A.; Flores, M.; et al. Rhizobium sp. strain NGR234 possesses a remarkable number of secretion systems. Appl. Environ. Microbiol. 2009, 75, 4035–4045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuldes, J.; Rodriguez-Orbegoso, M.; Schmeisser, C.; Krishnan, H.B.; Daniel, R.; Streit, W.R. Complete genome sequence of the broad-host-range strain Sinorhizobium fredii USDA257. J. Bacteriol. 2012, 194, 4483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinardell, J.M.; Acosta-Jurado, S.; Zehner, S.; Göttfert, M.; Becker, A.; Baena, I.; Blom, J.; Crespo-Rivas, J.C.; Goesmann, A.; Jaenicke, S.; et al. The Sinorhizobium fredii HH103 genome: A comparative analysis with S. fredii strains differing in their symbiotic behavior with soybean. Mol. Plant Microbe Interact. 2015, 28, 811–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Baena, F.J.; Vinardell, J.M.; Pérez-Montano, F.; Crespo-Rivas, J.C.; Bellogín, R.A.; Espuny, M.R.; Ollero, F.J. Regulation and symbiotic significance of nodulation outer proteins secretion in Sinorhizobium fredii HH103. Microbiology 2008, 154, 1825–1836. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Montaño, F.; Guasch-Vidal, B.; González-Barroso, S.; López-Baena, F.J.; Cubo, T.; Ollero, F.J.; Gil-Serrano, A.M.; Rodríguez-Carvajal, M.Á.; Bellogín, R.A.; Espuny, M.R. Nodulation-gene-inducing flavonoids increase overall production of autoinducers and expression of N-acyl homoserine lactone synthesis genes in rhizobia. Res. Microbiol. 2011, 162, 715–723. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Navarro-Gómez, P.; Murdoch, P.S.; Crespo-Rivas, J.C.; Jie, S.; Cuesta-Berrio, L.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.Á.; Vinardell, J.M. Exopolysaccharide Production by Sinorhizobium fredii HH103 Is Repressed by Genistein in a NodD1-Dependent Manner. PLoS ONE 2016, 11, e0160499. [Google Scholar] [CrossRef]
- Downie, J.A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev. 2010, 34, 150–170. [Google Scholar] [CrossRef]
- Wisniewski-Dyé, F.; Downie, J.A. Quorum-sensing in Rhizobium. Antonie Leeuwenhoek 2002, 81, 397–407. [Google Scholar] [CrossRef]
- González, J.E.; Marketon, M.M. Quorum sensing in nitrogen-fixing rhizobia. Microbiol. Mol. Biol. Rev. 2003, 67, 574–592. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Contreras, M.; Bauer, W.D.; Gao, M.; Robinson, J.B.; Allan Downie, J. Quorum-sensing regulation in rhizobia and its role in symbiotic interactions with legumes. Philos. Trans. R. Soc. Lond B Biol. Sci. 2007, 362, 1149–1163. [Google Scholar] [CrossRef] [Green Version]
- Calatrava-Morales, N.; McIntosh, M.; Soto, M.J. Regulation Mediated by N-acyl homoserine lactone quorum sensing signals in the Rhizobium-legume sSymbiosis. Genes 2018, 9, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beringer, J.E. R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 1974, 84, 188–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, J.M. The modified Fåhraeus slide technique. In A Manual for the Practical Study of Root Nodule Bacteria; Blackwell Scientific Publications: Oxford, UK, 1970; pp. 144–145. [Google Scholar]
- Bertani, G. Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [PubMed]
- Lamrabet, Y.; Bellogín, R.A.; Cubo, T.; Espuny, M.R.; Gil-Serrano, A.; Krishnan, H.B.; Megias, M.; Ollero, F.J.; Pueppke, S.G.; Ruiz-Sainz, J.E.; et al. Mutation in GDP fucose synthesis genes of Sinorhizobium fredii alters Nod factors and significantly decreases competitiveness to nodulate soybeans. Mol. Plant Microbe Interact. 1999, 12, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Madinabeitia, N.; Bellogín, R.A.; Buendía-Clavería, A.M.; Camacho, M.; Cubo, T.; Espuny, M.R.; Gil-Serrano, A.M.; Lyra, M.C.; Moussaid, A.; Ollero, F.J.; et al. Sinorhizobium fredii HH103 has a truncated nolO gene due to a -1 frameshift mutation that is conserved among other geographically distant S. fredii strains. Mol. Plant Microbe Interact. 2002, 15, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Cha, C.; Gao, P.; Chen, Y.C.; Shaw, P.D.; Farrand, S.K. Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant associated bacteria. Mol. Plant Microbe Interact. 1998, 11, 1119–1129. [Google Scholar] [CrossRef] [Green Version]
- Vinardell, J.M.; López-Baena, F.J.; Hidalgo, A.; Ollero, F.J.; Bellogín, R.; Espuny, M.R.; Temprano, F.; Romero, F.; Krishnan, H.B.; Pueppke, S.G.; et al. The effect of FITA mutations on the symbiotic properties of Sinorhizobium fredii varies in a chromosomal-background-dependent manner. Arch. Microbiol. 2004, 181, 144–154. [Google Scholar] [CrossRef]
- Figurski, D.H.; Helinski, D.R. Replication of an origin-containing derivative of plasmid RK2 dependent on a plasmid function provided in trans. Proc. Natl. Acad. Sci. USA 1979, 76, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1995, 145, 69–73. [Google Scholar] [CrossRef]
- Prentki, P.; Krisch, H.M. In vitro insertional mutagenesis with a selectable DNA fragment. Gene 1984, 29, 303–313. [Google Scholar] [CrossRef]
- Becker, A.; Schmidt, M.; Jäger, W.; Pühler, A. New gentamicin-resistance and lacZ promoter-probe cassettes suitable for insertion mutagenesis and generation of transcriptional fusions. Gene 1995, 162, 37–39. [Google Scholar] [CrossRef]
- Simon, R. High frequency mobilization of gram-negative bacterial replicons by the in vitro constructed TnS-Mob transposon. Mol. Gen. Genet. 1984, 196, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning. A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- López-Baena, F.J.; Monreal, J.A.; Pérez-Montaño, F.; Guasch-Vidal, B.; Bellogín, R.A.; Vinardell, J.M.; Ollero, F.J. The absence of Nops secretion in Sinorhizobium fredii HH103 increases GmPR1 expression in Williams soybean. Mol. Plant Microbe Interact. 2009, 22, 1445–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, D.; Grasland, B.; Vallee-Rehel, K.; Dufau, C.; Haras, D. On-line high-performance liquid chromatography-mass spectrometric detection and quantification of N-acylhomoserinelactones, quorum sensing signal molecules, in the presence of biological matrices. J. Chromatogr. 2003, 1002, 79–92. [Google Scholar] [CrossRef]
- Cataldi, T.R.I.; Bianco, G.; Abate, S.; Mattia, D. Analysis of S-adenosylmethionine and related sulfur metabolites in bacterial isolates of Pseudomonas aeruginosa (BAA-47) by liquid chromatography/electrospray ionization coupled to a hybrid linear quadrupole ion trap and fourier transform ion cyclotron resonance mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 3465–3477. [Google Scholar]
- Gould, T.A.; Herman, J.; Krank, J.; Murphy, R.C.; Churchill, M.E. Specificity of acyl-homoserine lactone synthases examined by mass spectrometry. J. Bacteriol. 2006, 188, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Pasini, P.; Daunert, S. Detection of bacterial quorum sensing N-acyl homoserine lactones in clinical samples. Anal. Bioanal. Chem. 2008, 391, 1619–1627. [Google Scholar] [CrossRef]
- Makemson, J.; Eberhard, A.; Mathee, K. Simple electrospray mass spectrometry detection of acylhomoserine lactones. Luminescence 2006, 21, 1–6. [Google Scholar] [CrossRef]
- Ortori, C.A.; Atkinson, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Barrett, D.A. Comprehensive profiling of N-acylhomoserine lactones produced by Yersinia pseudotuberculosis using liquid chromatography coupled to hybrid quadrupole-linear ion trap mass spectrometry. Anal. Bioanal. Chem. 2007, 387, 497–511. [Google Scholar] [CrossRef]
- Buendía-Clavería, A.M.; Moussaid, A.; Ollero, F.J.; Vinardell, J.M.; Torres, A.; Moreno, J.; Gil-Serrano, A.M.; Rodríguez-Carvajal, M.A.; Tejero-Mateo, P.; Peart, J.L.; et al. A purL mutant of Sinorhizobium fredii HH103 is symbiotically defective and altered in its lipopolysaccharide. Microbiology 2003, 149, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A.; Margaret, I.; Crespo-Rivas, J.C.; Parada, M.; Murdoch, P.S.; López, A.; Buendía-Clavería, A.M.; Moreno, J.; Albareda, M.; Gil-Serrano, A.M.; et al. The rkpU gene of Sinorhizobium fredii HH103 is required for bacterial K-antigen polysaccharide production and for efficient nodulation with soybean but not with cowpea. Microbiology 2010, 156, 3398–3411. [Google Scholar] [CrossRef] [PubMed]
- Sourjik, V.; Schmitt, R. Different roles of CheY1 and CheY2 in the chemotaxis of Rhizobium meliloti. Mol. Microbiol. 1996, 22, 427–436. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- Mueller, K.; González, J.E. Complex regulation of symbiotic functions is coordinated by MucR and quorum sensing in Sinorhizobium meliloti. J. Bacteriol. 2011, 193, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Montaño, F.; Jiménez-Guerrero, I.; Acosta-Jurado, S.; Navarro-Gómez, P.; Ollero, F.J.; Ruiz-Sainz, J.E.; López-Baena, F.J.; Vinardell, J.M. A transcriptomic analysis of the effect of flavonoids on Sinorhizobium fredii HH103 reveals novel rhizobial genes putatively involved in symbiosis. Sci. Rep. 2016, 6, 31592. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Santos-Garcia, D.; Pérez-Montaño, F.; da Silva, G.M.; Zhao, M.; Jiménez-Guerrero, I.; Rosenberg, T.; Chen, G.; Plaschkes, I.; Morin, S.; et al. Complete Assembly of the Genome of an Acidovorax citrulli Strain Reveals a Naturally Occurring Plasmid in This Species. Front. Microbiol. 2019, 10, 1400. [Google Scholar] [CrossRef] [Green Version]
- Hoang, H.H.; Becker, A.; González, J.E. The LuxR homolog ExpR, in combination with the Sin quorum sensing system, plays a central role in Sinorhizobium meliloti gene expression. J. Bacteriol. 2004, 186, 5460–5472. [Google Scholar] [CrossRef] [Green Version]
- López-Baena, F.J.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.A.; Vinardell, J.M. Bacterial Molecular Signals in the Sinorhizobium fredii-Soybean Symbiosis. Int. J. Mol. Sci. 2016, 17, 755. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Montaño, F.; Jiménez-Guerrero, I.; Del Cerro, P.; Baena-Ropero, I.; López-Baena, F.J.; Ollero, F.J.; Bellogín, R.; Lloret, J.; Espuny, R. The Symbiotic Biofilm of Sinorhizobium fredii SMH12, Necessary for Successful Colonization and Symbiosis of Glycine max cv Osumi, is Regulated by Quorum Sensing Systems and Inducing Flavonoids via NodD1. PLoS ONE 2014, 9, e105901. [Google Scholar] [CrossRef] [Green Version]
- Marketon, M.M.; Gronquist, M.R.; Eberhard, A.; González, J.E. Characterization of the Sinorhizobium meliloti sinR/sinI locus and the production of novel N-acyl homoserine lactones. J. Bacteriol. 2002, 184, 5686–5695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marketon, M.M.; González, J.E. Identification of two quorum-sensing systems in Sinorhizobium meliloti. J. Bacteriol. 2002, 184, 3466–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoenpanich, P.; Meyer, S.; Becker, A.; McIntosh, M. Temporal expression program of quorum sensing-based transcription regulation in Sinorhizobium meliloti. J. Bacteriol. 2013, 195, 3224–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; Chen, H.; Eberhard, A.; Gronquist, M.R.; Robinson, J.B.; Rolfe, B.G.; Bauer, W.D. sinI- and expR-dependent quorum sensing in Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 7931–7944. [Google Scholar] [CrossRef] [Green Version]
- Gurich, N.; González, J.E. Role of quorum sensing in Sinorhizobium meliloti-alfalfa symbiosis. J. Bacteriol. 2009, 191, 4372–4382. [Google Scholar] [CrossRef] [Green Version]
- Nasser, W.; Reverchon, S. New insights into the regulatory mechanisms of the LuxR family of quorum sensing regulators. Anal. Bioanal. Chem. 2007, 387, 381. [Google Scholar] [CrossRef]
- Choi, S.H.; Greenberg, E.P. The C-terminal region of the Vibrio fischeri LuxR protein contains an inducer-independent lux gene activating domain. Proc. Natl. Acad. Sci. USA 1991, 88, 11115–11119. [Google Scholar] [CrossRef] [Green Version]
- Fuqua, W.C.; Winans, S.C. A LuxR-LuxI type regulatory system activates Agrobacterium Ti plasmid conjugal transfer in the presence of a plant tumor metabolite. J. Bact. 1994, 176, 2796–2806. [Google Scholar] [CrossRef] [Green Version]
- Grote, J.; Krysciak, D.; Petersen, K.; Gullert, S.; Schmeisser, C.; Forstner, K.U.; Krishnan, H.B.; Schwalbe, H.; Kubatova, N.; Streit, W.R. The absence of the N-acyl-homoserine-lactone autoinducer synthase genes traI and ngrI increases the copy number of the symbiotic plasmid in Sinorhizobium fredii NGR234. Front. Microbiol. 2016, 7, 1858. [Google Scholar] [CrossRef]
- He, X.; Chang, W.; Pierce, D.L.; Seib, L.O.; Wagner, J.; Fuqua, C. Quorum sensing in Rhizobium sp. Strain NGR234 regulates conjugal transfer (tra) gene expression and influences growth rate. J. Bacteriol. 2003, 185, 809–822. [Google Scholar] [CrossRef] [Green Version]
- Krysciak, D.; Grote, J.; Rodriguez Orbegoso, M.; Utpatel, C.; Forstner, K.U.; Li, L.; Schmeisser, C.; Krishnan, H.B.; Streit, W.R. RNA sequencing analysis of the broad-host-range strain Sinorhizobium fredii NGR234 identifies a large set of genes linked to quorum sensing-dependent regulation in the background of a traI and ngrI deletion mutant. Appl. Environ. Microbiol. 2014, 80, 5655–5671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.; Smyth, A.J.; Luo, Z.Q.; Farrand, S.K. Modulating quorum sensing by antiactivation: TraM interacts with TraR to inhibit activation of Ti plasmid conjugal transfer genes. Mol. Microbiol. 1999, 34, 282–294. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, M.; Meyer, S.; Becker, A. Novel Sinorhizobium meliloti quorum sensing positive and negative regulatory feedback mechanisms respond to phosphate availability. Mol. Microbiol. 2009, 74, 1238–1256. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Jurado, S.; Rodríguez-Navarro, D.N.; Kawaharada, Y.; Rodríguez-Carvajal, M.A.; Gil-Serrano, A.; Soria-Díaz, M.E.; Pérez-Montaño, F.; Fernández-Perea, J.; Niu, Y.; Alias-Villegas, C.; et al. Sinorhizobium fredii HH103 nolR and nodD2 mutants gain capacity for infection thread invasion of Lotus japonicus Gifu and Lotus burttii. Environ. Microbiol. 2019, 21, 1718–1739. [Google Scholar] [CrossRef]
- Fujishige, N.A.; Lum, M.R.; De Hoff, P.L.; Whitelegge, J.P.; Faull, K.F.; Hirsch, A.M. Rhizobium common nod genes are required for biofilm formation. Mol. Microbiol. 2008, 67, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Alias-Villegas, C.; Navarro-Gómez, P.; Zehner, S.; Murdoch, P.D.S.; Rodríguez-Carvajal, M.A.; Soto, M.J.; Ollero, F.J.; Ruiz-Sainz, J.E.; Göttfert, M.; et al. The Sinorhizobium fredii HH103 MucR1 global regulator is connected with the nod regulon and is required for efficient symbiosis with Lotus burttii and Glycine max cv. Williams. Mol. Plant Microbe Interact. 2016, 29, 700–712. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Alias-Villegas, C.; Navarro-Gómez, P.; Almozara, A.; Rodríguez-Carvajal, M.A.; Medina, C.; Vinardell, J.M. Sinorhizobium fredii HH103 syrM inactivation affects the expression of a large number of genes, impairs nodulation with soybean, and extends the host-range to the Lotus japonicus. Environ. Microbiol. 2019. [Google Scholar] [CrossRef]
- Luo, Z.Q.; Farrand, S.K. Signal-dependent DNA binding and functional domains of the quorum-sensing activator TraR as identified by repressor activity. Proc. Natl. Acad. Sci. USA 1999, 96, 9009–9014. [Google Scholar] [CrossRef] [Green Version]
- Parada, M.; Vinardell, J.M.; Ollero, F.J.; Hidalgo, A.; Gutiérrez, R.; Buendía-Clavería, A.M.; Lei, W.; Margaret, I.; López-Baena, F.J.; Gil-Serrano, A.M.; et al. Sinorhizobium fredii HH103 mutants affected in capsular polysaccharide (KPS) are impaired for nodulation with soybean and Cajanus cajan. Mol. Plant Microbe Interact. 2006, 19, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Alías-Villegas, C.; Navarro-Gómez, P.; Almorzara, A.; Soto, M.J.; Vinardell, J.M.; Acosta-Jurado, S. Surface motility in Sinorhizobium fredii HH103 Is activated by Genistein in a NodD1-Dependent Manner. 2019; Manuscript in preparation. [Google Scholar]
- Fraysse, N.; Couderc, F.; Poinsot, V. Surface polysaccharide involvement in establishing the Rhizobium legume symbiosis. Eur. J. Biochem. 2003, 270, 1365–1380. [Google Scholar] [CrossRef]
- Janczarek, M. Environmental signals and regulatory pathways that influence exopolysaccharide production in rhizobia. Int. J. Mol. Sci. 2011, 12, 7898–7933. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Relevant Properties | Source or Reference |
|---|---|---|
| S. fredii | ||
| HH103 | Wild-type strain, spontaneous RifR | [27] |
| HH103 traI | HH103 traI::Ω, SpcR | This work |
| HH103 sinI | HH103 sinI::lacZ-GmR, GmR | This work |
| HH103 traI sinI | HH103 traI::Ω sinI::lacZ-GmR, SpcR GmR | This work |
| E. coli | ||
| DH5α | fhuA2 lac(del)U169 phoA glnV44Φ80’ lacZ(del)M15 gyrA96 recA1 relA1 endA1 thi-1 hsdR17, NalR | Stratagene (USA) |
| A. tumefaciens | ||
| NT1 (pZRL4) | A. tumefaciens devoid of pTiC58 and harboring pZRL4, which carries the fusion traG::lacZ and the traR gene | [28] |
| GMI9023 (pMUS248) | A. tumefaciens strain devoid of pTi and pAtC58 (C58 derivative), harboring plasmid pMUS248, KmR and stable in rhizobia, which carries the fusion pnodA::tetΔp, KmR | [29] |
| Plasmids | ||
| pRK2013 | Helper plasmid, KmR | [30] |
| pGEM-T Easy | PCR cloning vector, ApR | Promega (USA) |
| pK18mob | Cloning vector, suicide in rhizobia, KmR | [31] |
| pHP45Ω | ApR vector containing the Ω interposon, ApR SpcR | [32] |
| pAB2001 | ApR vector containing the lacZ-GmR cassette, ApR GmR | [33] |
| pMUS997 | pGEM-T Easy::traI, ApR | This work |
| pMUS1079 | pGEM-T Easy::sinI, ApR | This work |
| pMUS989 | pK18mob::traI, KmR | This work |
| pMUS1083 | pK18mob::sinI, KmR | This work |
| pMUS1006 | pK18mob::traI::Ω, KmR SpcR | This work |
| pMUS1087 | pK18mob::sinI::lacZ-GmR, KmR GmR | This work |
| Name | Sequence | Usage |
|---|---|---|
| traI F | 5′-CCAGAAGATTGGGATTGACA-3′ | Amplification of the traI gene |
| traI R | 5′-TGTCCGCCTATCGGAAGCTCA-3′ | |
| sinI F | 5′-TTTTCATGCGTCGATGCTCGA-3′ | Amplification of the sinI gene |
| sinI R | 5′-CCGTAGGTCG GAACAATGACA-3′ | |
| ligEq_F | 5′-AAGACCAAGCTGTCGCTC-3′ | Chromosomal gene, qPCR assays |
| ligEq_R | 5′-ATGTCGAAGCTGTCGCTG-3′ | |
| ftsZ1q_F | 5′-ATACGCTGATCGTCATCC-3′ | Chromosomal gene, qPCR assays |
| ftsZ1q_R | 5′-GCCTTCCTTGACCATGAG-3′ | |
| flgJq_F | 5′-TGCTGAATTCCTCGGAAG-3′ | Chromosomal gene, qPCR assays |
| flgJq_R | 5′-CAGCATCGACTTGACGAA-3′ | |
| nolRq_F | 5′-CCAAAACGCCTGCTCATT-3′ | Chromosomal gene, qPCR assays |
| nolRq_R | 5′-ATTCTGGGCACGCAACTT-3′ | |
| nodAq_F | 5′-ACGTCATGTATCCGGTGCTGCA-3′ | pSym gene, qPCR assays |
| nodAq_R | 5′-CGTTGGCGGCAGGTTGAGA-3′ | |
| syrMq_F | 5′-GTTCAATGACGATCTCTTGGT-3′ | pSym gene, qPCR assays |
| syrMq_R | 5′-ATTGCCATAGTTACCTTCGAC-3′ | |
| d373q_F | 5′-TCGACGATTCAATAAGGGTG-3′ | pSym gene, qPCR assays |
| d373q_R | 5′-CATATCCTCTCCGCAATAGC-3′ | |
| d161q_F | 5′-AGAATGTCGCATACCTCTTAG-3′ | pSym gene, qPCR assays |
| d161q_R | 5′-GTGAAGGCTGTTATCCCATC-3′ | |
| qnodD1-F | 5′-GCGAGCACGGACTGCGAA-3′ | pSym gene, confirmation of conjugation transfer of this plasmid to GMI9023 (pMUS248) |
| qnodD1-R | 5′-CGGGAAAAATGGGTTGCGGA-3′ |
| Standars | Strain | |||
|---|---|---|---|---|
| HH103 | traI | sinI | traI/sinI | |
| C6-HSL | − | − | − | − |
| 3-oxo-C6-HSL | − | − | − | − |
| C8-HSL | + | − | + | − |
| 3-oxo-C8-HSL | + | − | + | − |
| C10-HSL | − | − | − | − |
| 3-oxo-C10-HSL | − | − | − | − |
| C12-HSL | + | + | − | − |
| 3-oxo-C12-HSL | − | − | − | − |
| 3-OH-C12-HSL | − | − | − | − |
| C14-HSL | + | + | − | − |
| 3-oxo-C14-HSL | + | + | − | − |
| 3-OH-C14-HSL | − | + | − | − |
| Gene ID | Gene Name | Number of Transcripts | Relative Position (among 7014 Total Number of ORFs) | Transcriptomic Ranking |
|---|---|---|---|---|
| Quorum sensing genes | ||||
| psfHH103d_478 | traI | 54 | 5662 | 19.3 |
| SFHH103_01572 | sinI | 82 | 4910 | 30.0 |
| psfHH103d_461 | traM | 2208 | 500 | 92.9 |
| psfHH103d_462 | traR end | 541 | 1629 | 76.8 |
| psfHH103d_463 | traR start | 1730 | 638 | 90.9 |
| SFHH103_1571 | sinR | 512 | 1704 | 75.7 |
| SFHH103_03306 | expR end | 450 | 1863 | 73.4 |
| SFHH103_03432 | expR start | 350 | 2207 | 68.5 |
| Nodulation genes | ||||
| psfHH103d_386 | nodD1 | 1579 | 696 | 90.1 |
| psfHH103d_126 | nodA | 75 | 5090 | 27.4 |
| psfHH103d_127 | nodB | 156 | 3611 | 48.5 |
| psfHH103d_128 | nodC | 316 | 2363 | 66.3 |
| psfHH103d_129 | nodI | 86 | 4819 | 31.3 |
| psfHH103d_130 | nodJ | 116 | 4199 | 40.1 |
| psfHH103d_131 | nolO’ | 376 | 2101 | 70.0 |
| psfHH103d_132 | noeI | 186 | 3281 | 53.2 |
| psfHH103d_381 | nodZ | 1022 | 979 | 86.0 |
| psfHH103d_380 | noeL | 571 | 1562 | 77.7 |
| psfHH103d_339 | nolU | 189 | 3243 | 53.8 |
| Average | 425 | 2904 | 58.6 | |
| Chemotaxis and motility genes | ||||
| SFHH103_00293 | mcpE | 107 | 4367 | 37.7 |
| SFHH103_00294 | cheX | 23 | 6642 | 5.3 |
| SFHH103_00295 | cheY1 | 21 | 6700 | 4.5 |
| SFHH103_00296 | cheA | 128 | 3200 | 54.4 |
| SFHH103_00297 | cheW1 | 9 | 6939 | 1.1 |
| SFHH103_00298 | cheR | 30 | 6436 | 8.2 |
| SFHH103_00299 | cheB | 46 | 5931 | 15.4 |
| SFHH103_00300 | cheY2 | 33 | 6345 | 9.5 |
| SFHH103_00301 | cheD | 30 | 6437 | 8.2 |
| SFHH103_00303 | fliF | 108 | 4341 | 38.1 |
| SFHH103_00304 | visN | 660 | 1410 | 79.9 |
| SFHH103_00305 | visR | 489 | 1757 | 75.0 |
| SFHH103_00307 | flhB | 142 | 3775 | 46.2 |
| SFHH103_00308 | fliG | 45 | 5958 | 15.1 |
| SFHH103_00309 | fliN | 44 | 5993 | 14.6 |
| SFHH103_00310 | fliM | 37 | 6215 | 11.4 |
| SFHH103_00311 | motA | 37 | 6225 | 11.2 |
| SFHH103_00313 | flgF | 29 | 6454 | 8.0 |
| SFHH103_00314 | fliI | 42 | 6058 | 13.6 |
| SFHH103_00316 | flgB | 51 | 5757 | 17.9 |
| SFHH103_00317 | flgC | 16 | 6819 | 2.8 |
| SFHH103_00318 | fliE | 21 | 6686 | 4.7 |
| SFHH103_00319 | flgG | 29 | 6469 | 7.8 |
| SFHH103_00320 | flgA | 29 | 6470 | 7.8 |
| SFHH103_00321 | flgI | 27 | 6534 | 6.8 |
| SFHH103_00323 | flgH | 48 | 5857 | 16.5 |
| SFHH103_00324 | fliL | 44 | 5991 | 14.6 |
| SFHH103_00325 | fliP | 135 | 3886 | 44.6 |
| SFHH103_00326 | flaC | 493 | 1746 | 75.1 |
| SFHH103_00327 | flaB | 592 | 1516 | 78.4 |
| SFHH103_00328 | flaA | 193 | 3192 | 54.5 |
| SFHH103_00329 | flaD | 140 | 3810 | 45.7 |
| SFHH103_00331 | motB | 82 | 4918 | 29.9 |
| SFHH103_00332 | motC | 59 | 5549 | 20.9 |
| SFHH103_00333 | motD | 93 | 4647 | 33.7 |
| SFHH103_00336 | flgE | 84 | 4865 | 30.6 |
| SFHH103_00337 | flgK | 66 | 5315 | 24.2 |
| SFHH103_00338 | flgL | 103 | 4446 | 36.6 |
| SFHH103_00339 | flaF | 61 | 5498 | 21.6 |
| SFHH103_00340 | flbT | 62 | 5444 | 22.4 |
| SFHH103_00341 | flgD | 66 | 5338 | 23.9 |
| SFHH103_00342 | fliQ | 135 | 3890 | 44.5 |
| SFHH103_00343 | flhA | 119 | 4144 | 40.9 |
| SFHH103_00344 | fliR | 75 | 5092 | 27.4 |
| SFHH103_00346 | flgJ | 27 | 6541 | 6.7 |
| Average | 109 | 5147 | 26.6 | |
| Exopolysaccharydes genes | ||||
| SFHH103_01240 | exoR | 27591 | 27 | 99.6 |
| SFHH103_02875 | exoN | 2678 | 377 | 94.6 |
| SFHH103_03541 | exoS | 777 | 1237 | 82.4 |
| SFHH103_03846 | exoD | 1785 | 623 | 91.1 |
| SFHH103_05372 | exoP | 7993 | 110 | 98.4 |
| SFHH103_05373 | exoN | 2916 | 422 | 94.0 |
| SFHH103_05374 | exoO | 1191 | 881 | 87.4 |
| SFHH103_05375 | exoM | 2698 | 420 | 94.0 |
| SFHH103_05376 | exoA | 4029 | 270 | 96.2 |
| SFHH103_05377 | exoL | 7280 | 124 | 98.2 |
| SFHH103_05378 | exoK | 15966 | 52 | 99.3 |
| SFHH103_05380 | exoI | 278 | 2537 | 63.8 |
| SFHH103_05382 | exoU | 2212 | 498 | 92.9 |
| SFHH103_05383 | exoX | 787 | 1222 | 82.6 |
| SFHH103_05384 | exoY2 | 33321 | 23 | 99.7 |
| SFHH103_05386 | exoF1 | 5959 | 160 | 97.7 |
| SFHH103_05387 | exoQ | 1783 | 625 | 91.1 |
| SFHH103_05388 | exoZ | 981 | 1020 | 85.5 |
| SFHH103_05389 | exoB | 7200 | 126 | 98.2 |
| SFHH103_05659 | exoF2 | 66 | 5332 | 24.0 |
| SFHH103_05660 | exoY1 | 123 | 4088 | 41.7 |
| SFHH103_05850 | exoF3 | 519 | 1686 | 76.0 |
| Average | 5824 | 994 | 85.8 | |
| Plasmid transfer genes | ||||
| psfHH103d_56 | traG | 75 | 5082 | 27.5 |
| psfHH103d_57 | traD | 16 | 6820 | 2.8 |
| psfHH103d_57_5 | traC | 20 | 6721 | 4.2 |
| psfHH103d_58 | traA | 260 | 2638 | 62.4 |
| psfHH103d_465 | trbI | 87 | 4801 | 31.6 |
| psfHH103d_466 | trbH | 25 | 6570 | 6.3 |
| psfHH103d_467 | trbG | 23 | 6643 | 5.3 |
| psfHH103d_468 | trbF | 24 | 6612 | 5.7 |
| psfHH103d_469 | trbL | 65 | 5364 | 23.5 |
| psfHH103d_470 | trbK | 17 | 6777 | 3.4 |
| psfHH103d_471 | trbJ | 49 | 5820 | 17.0 |
| psfHH103d_472 | trbE | 68 | 5265 | 24.9 |
| psfHH103d_475 | trbD | 42 | 6055 | 13.7 |
| psfHH103d_476 | trbC | 35 | 6292 | 10.3 |
| psfHH103d_477 | trbB | 60 | 5531 | 21.1 |
| SFHH103_06246 | traA | 244 | 2740 | 60.9 |
| SFHH103_06247 | traD | 48 | 5878 | 16.2 |
| SFHH103_06248 | traG | 116 | 4200 | 40.1 |
| SFHH103_03975 | traG | 176 | 3396 | 51.6 |
| SFHH103_03977 | traA | 1219 | 861 | 87.7 |
| SFHH103_03995 | traG | 362 | 2158 | 69.2 |
| SFHH103_03999 | traA | 1443 | 746 | 89.4 |
| Average | 203 | 4862 | 30.7 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acosta-Jurado, S.; Alías-Villegas, C.; Almozara, A.; Espuny, M.R.; Vinardell, J.-M.; Pérez-Montaño, F. Deciphering the Symbiotic Significance of Quorum Sensing Systems of Sinorhizobium fredii HH103. Microorganisms 2020, 8, 68. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010068
Acosta-Jurado S, Alías-Villegas C, Almozara A, Espuny MR, Vinardell J-M, Pérez-Montaño F. Deciphering the Symbiotic Significance of Quorum Sensing Systems of Sinorhizobium fredii HH103. Microorganisms. 2020; 8(1):68. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010068
Chicago/Turabian StyleAcosta-Jurado, Sebastián, Cynthia Alías-Villegas, Andrés Almozara, M. Rosario Espuny, José-María Vinardell, and Francisco Pérez-Montaño. 2020. "Deciphering the Symbiotic Significance of Quorum Sensing Systems of Sinorhizobium fredii HH103" Microorganisms 8, no. 1: 68. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8010068