Proteomic 2D-DIGE Analysis of Milk Whey from Dairy Cows with Staphylococcus aureus Mastitis Reveals Overexpression of Host Defense Proteins


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cows and Sample Collection
2.2. Somatic Cell Counts (SCC) and Microbiological Examination
2.3. Sample Processing and Two-Dimensional Differential Gel Electrophoresis (2D-DIGE)
2.4. In-Gel Digestion and Liquid Chromatography–Tandem Mass Spectrometry (LC–MS/MS) Analysis
2.5. Protein Identification
2.6. Gene Ontology (GO) and Pathway Enrichment Analyses
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halasa, T.; Huijps, K.; Østerås, O.; Hogeveen, H. Economic effects of bovine mastitis and mastitis management: A review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Hogeveen, H.; Huijps, K.; Lam, T.J. Economic Aspects of Mastitis: New Developments. N. Z. Vet. J. 2011, 59, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.D.; Paape, M.J.; Lee, J.-W.; Zhao, X.; Hope, J.C.; Rainard, P. Escherichia coli and Staphylococcus aureus Elicit Differential Innate Immune Responses following Intramammary Infection. Clin. Diagn. Lab. Immunol. 2004, 11, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atalla, H.; Gyles, C.; Mallard, B. Persistence of a Staphylococcus aureus Small Colony Variants (S. Aureus Scv) within Bovine Mammary Epithelial Cells. Vet. Microbiol. 2010, 143, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Barkema, H.; Schukken, Y.; Zadoks, R. Invited Review: The Role of Cow, Pathogen, and Treatment Regimen in the Therapeutic Success of Bovine Staphylococcus aureus Mastitis. J. Dairy Sci. 2006, 89, 1877–1895. [Google Scholar] [CrossRef] [Green Version]
- Bannerman, D.D. Pathogen-dependent induction of cytokines and other soluble inflammatory mediators during intramammary infection of dairy cows1. J. Anim. Sci. 2009, 87, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Atalla, H.; Mallard, B.A.; Robert, C.; A Karrow, N. Changes in Holstein cow milk and serum proteins during intramammary infection with three different strains of Staphylococcus aureus. BMC Vet. Res. 2011, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Reinhardt, T.A.; Sacco, R.E.; Nonnecke, B.J.; Lippolis, J.D. Bovine milk proteome: Quantitative changes in normal milk exosomes, milk fat globule membranes and whey proteomes resulting from Staphylococcus aureus mastitis. J. Proteomics 2013, 82, 141–154. [Google Scholar] [CrossRef]
- Abdelmegid, S.; Murugaiyan, J.; Schenkel, F.; Caswell, J.L.; Kelton, D.; Kirby, G. Identification of Host Defense-Related Proteins Using Label-Free Quantitative Proteomic Analysis of Milk Whey from Cows with Staphylococcus aureus Subclinical Mastitis. Int. J. Mol. Sci. 2017, 19, 78. [Google Scholar] [CrossRef] [Green Version]
- Harmon, R.; Eberhart, R.; Jasper, D. Microbiological Procedures for the Diagnosis of Bovine Udder Infection; National Mastitis Council: Arlington, VA, USA, 1990. [Google Scholar]
- Atalla, H.; Gyles, C.; Jacob, C.L.; Moisan, H.; Malouin, F.; Mallard, B.A. Characterization of a Staphylococcus aureus Small Colony Variant (SCV) Associated with Persistent Bovine Mastitis. Foodborne Pathog. Dis. 2008, 5, 785–799. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using David Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, M.; Kanehisa, M. Using the Kegg Database Resource. Curr. Protoc. Bioinform. 2012, 11, 1.12.1–1.12.54. [Google Scholar] [CrossRef] [PubMed]
- Staphylococcus aureus—Swiss-Model Repository. Available online: https://swissmodel.expasy.org/repository/species/93061 (accessed on 26 November 2020).
- Maity, S.; Das, D.; Ambatipudi, K. Quantitative Alterations in Bovine Milk Proteome from Healthy, Subclinical and Clinical Mastitis During S. Aureus Infection. J. Proteomics 2020, 223, 103815. [Google Scholar] [CrossRef] [PubMed]
- Pisanu, S.; Cacciotto, C.; Pagnozzi, D.; Puggioni, G.M.G.; Uzzau, S.; Ciaramella, P.; Guccione, J.; Penati, M.; Pollera, C.; Moroni, P.; et al. Proteomic changes in the milk of water buffaloes (Bubalus bubalis) with subclinical mastitis due to intramammary infection by Staphylococcus aureus and by non-aureus staphylococci. Sci. Rep. 2019, 9, 15850. [Google Scholar] [CrossRef]
- Huang, J.; Luo, G.; Zhang, Z.; Wang, X.; Ju, Z.; Qi, C.; Zhang, Y.; Wang, C.; Li, R.; Li, J.; et al. iTRAQ-proteomics and bioinformatics analyses of mammary tissue from cows with clinical mastitis due to natural infection with Staphylococci aureus. BMC Genom. 2014, 15, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Smolenski, G.A.; Broadhurst, M.; Stelwagen, K.; Haigh, B.; Wheeler, T.T. Host defence related responses in bovine milk during an experimentally induced Streptococcus uberis infection. Proteome Sci. 2014, 12, 19. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.C.; Mudaliar, M.; Tassi, R.; McNeilly, T.N.; Burchmore, R.; Burgess, K.; Herzyk, P.; Zadoks, R.; Eckersall, P.D. Mastitomics, the integrated omics of bovine milk in an experimental model of Streptococcus uberis mastitis: 3. Untargeted metabolomics. Mol. BioSyst. 2016, 12, 2762–2769. [Google Scholar] [CrossRef]
- Larsen, L.; Hinz, K.; Jørgensen, A.; Møller, H.; Wellnitz, O.; Bruckmaier, R.; Kelly, A.L. Proteomic and peptidomic study of proteolysis in quarter milk after infusion with lipoteichoic acid from Staphylococcus aureus. J. Dairy Sci. 2010, 93, 5613–5626. [Google Scholar] [CrossRef] [Green Version]
- Boehmer, J.L. Proteomic Analyses of Host and Pathogen Responses during Bovine Mastitis. J. Mammary Gland. Biol. Neoplasia 2011, 16, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Boehmer, J.L.; Bannerman, D.; Shefcheck, K.; Ward, J. Proteomic Analysis of Differentially Expressed Proteins in Bovine Milk During Experimentally Induced Escherichia coli Mastitis. J. Dairy Sci. 2008, 91, 4206–4218. [Google Scholar] [CrossRef] [Green Version]
- Hogarth, C.J.; Fitzpatrick, J.L.; Nolan, A.M.; Young, F.J.; Pitt, A.R.; Eckersall, P.D. Differential protein composition of bovine whey: A comparison of whey from healthy animals and from those with clinical mastitis. Proteomics 2004, 4, 2094–2100. [Google Scholar] [CrossRef] [PubMed]
- Smolenski, G.; Haines, S.; Kwan, F.Y.-S.; Bond, J.; Farr, V.; Davis, S.R.; Stelwagen, A.K.; Wheeler, T.T. Characterisation of Host Defence Proteins in Milk Using a Proteomic Approach. J. Proteome Res. 2007, 6, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Murakami, K.; Wallingford, J.C.; Yuki, Y. Identification of low-abundance proteins of bovine colostral and mature milk using two-dimensional electrophoresis followed by microsequencing and mass spectrometry. Electrophoresis 2002, 23, 1153–1160. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Ibeagha, A.E.; Messier, S.; Zhao, X. Proteomics, Genomics, and Pathway Analyses of Escherichia Coli and Staphylococcus Aureus Infected Milk Whey Reveal Molecular Pathways and Networks Involved in Mastitis. J. Proteome Res. 2010, 9, 4604–4619. [Google Scholar] [CrossRef]
- Rainard, P.; Riollet, C. Innate immunity of the bovine mammary gland. Vet. Res. 2006, 37, 369–400. [Google Scholar] [CrossRef] [Green Version]
- Kaetzel, C.S. The polymeric immunoglobulin receptor: Bridging innate and adaptive immune responses at mucosal surfaces. Immunol. Rev. 2005, 206, 83–99. [Google Scholar] [CrossRef]
- Stelwagen, K.; Carpenter, E.A.; Haigh, B.; Hodgkinson, A.J.; Wheeler, T.T. Immune components of bovine colostrum and milk1. J. Anim. Sci. 2009, 87, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Ezzat Alnakip, M.; Quintela-Baluja, M.; Böhme, K.; Fernández-No, I.; Caamaño-Antelo, S.; Calo-Mata, P.; Barros-Velázquez, J. The Immunology of Mammary Gland of Dairy Ruminants between Healthy and Inflammatory Conditions. J. Vet. Med. 2014, 2014, 659801. [Google Scholar] [CrossRef] [Green Version]
- Murgiano, L.; Timperio, A.M.; Zolla, L.; Bongiorni, S.; Valentini, A.; Pariset, L. Comparison of Milk Fat Globule Membrane (MFGM) Proteins of Chianina and Holstein Cattle Breed Milk Samples Through Proteomics Methods. Nutrients 2009, 1, 302–315. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.; Rainard, P.; Lippolis, J.D.; Salmon, H.; Kacskovics, I. The Mammary Gland in Mucosal and Regional Immunity. Mucosal Immunol. 2015, 2269–2306. [Google Scholar] [CrossRef]
- Katsafadou, A.I.; Tsangaris, G.T.; Anagnostopoulos, A.K.; Billinis, C.; Barbagianni, M.S.; Vasileiou, N.G.; Spanos, S.A.; Mavrogianni, V.S.; Fthenakis, G.C. Proteomics data of ovine mastitis associated with Mannheimia haemolytica. Data Brief 2019, 25, 104259. [Google Scholar] [CrossRef] [PubMed]
- Bian, Y.; Lv, Y.; Li, Q.-Z. Identification of Diagnostic Protein Markers of Subclinical Mastitis in Bovine Whey Using Comparative Proteomics. Bull. Vet. Inst. Pulawy 2014, 58, 385–392. [Google Scholar] [CrossRef] [Green Version]
- Larsen, L.B.; Rasmussen, M.D.; Bjerring, M.; Nielsen, J.H. Proteases and protein degradation in milk from cows infected with Streptococcus uberis. Int. Dairy J. 2004, 14, 899–907. [Google Scholar] [CrossRef]
- Zhao, X.; Lacasse, P. Mammary tissue damage during bovine mastitis: Causes and control1. J. Anim. Sci. 2008, 86, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Scanu, A.M.; Edelstein, C. HDL: Bridging past and present with a look at the future. FASEB J. 2008, 22, 4044–4054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehmer, J.L.; DeGrasse, J.A.; McFarland, M.A.; Tall, E.; Shefcheck, K.J.; Ward, J.L.; Bannerman, D.D. The proteomic advantage: Label-free quantification of proteins expressed in bovine milk during experimentally induced coliform mastitis. Vet. Immunol. Immunopathol. 2010, 138, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, M.; Codrea, M.C.; Ingvartsen, K.L.; Friggens, N.C.; Bendixen, E.; Røntved, C.M. Quantitative milk proteomics—Host responses to lipopolysaccharide-mediated inflammation of bovine mammary gland. Proteomics 2010, 10, 2240–2249. [Google Scholar] [CrossRef] [PubMed]
- Bendixen, E.; Danielsen, M.; Hollung, K.; Gianazza, E.; Miller, I. Farm Animal Proteomics—A Review. J. Proteomics 2011, 74, 282–293. [Google Scholar] [CrossRef]
- Chiaradia, E.; Valiani, A.; Tartaglia, M.; Scoppetta, F.; Renzone, G.; Arena, S.; Avellini, L.; Benda, S.; Gaiti, A.; Scaloni, A. Ovine subclinical mastitis: Proteomic analysis of whey and milk fat globules unveils putative diagnostic biomarkers in milk. J. Proteomics 2013, 83, 144–159. [Google Scholar] [CrossRef]
- Chang, W.W.P.; Hobson, C.; Bomberger, D.C.; Schneider, L.V. Rapid separation of protein isoforms by capillary zone electrophoresis with new dynamic coatings. Electrophoresis 2005, 26, 2179–2186. [Google Scholar] [CrossRef]


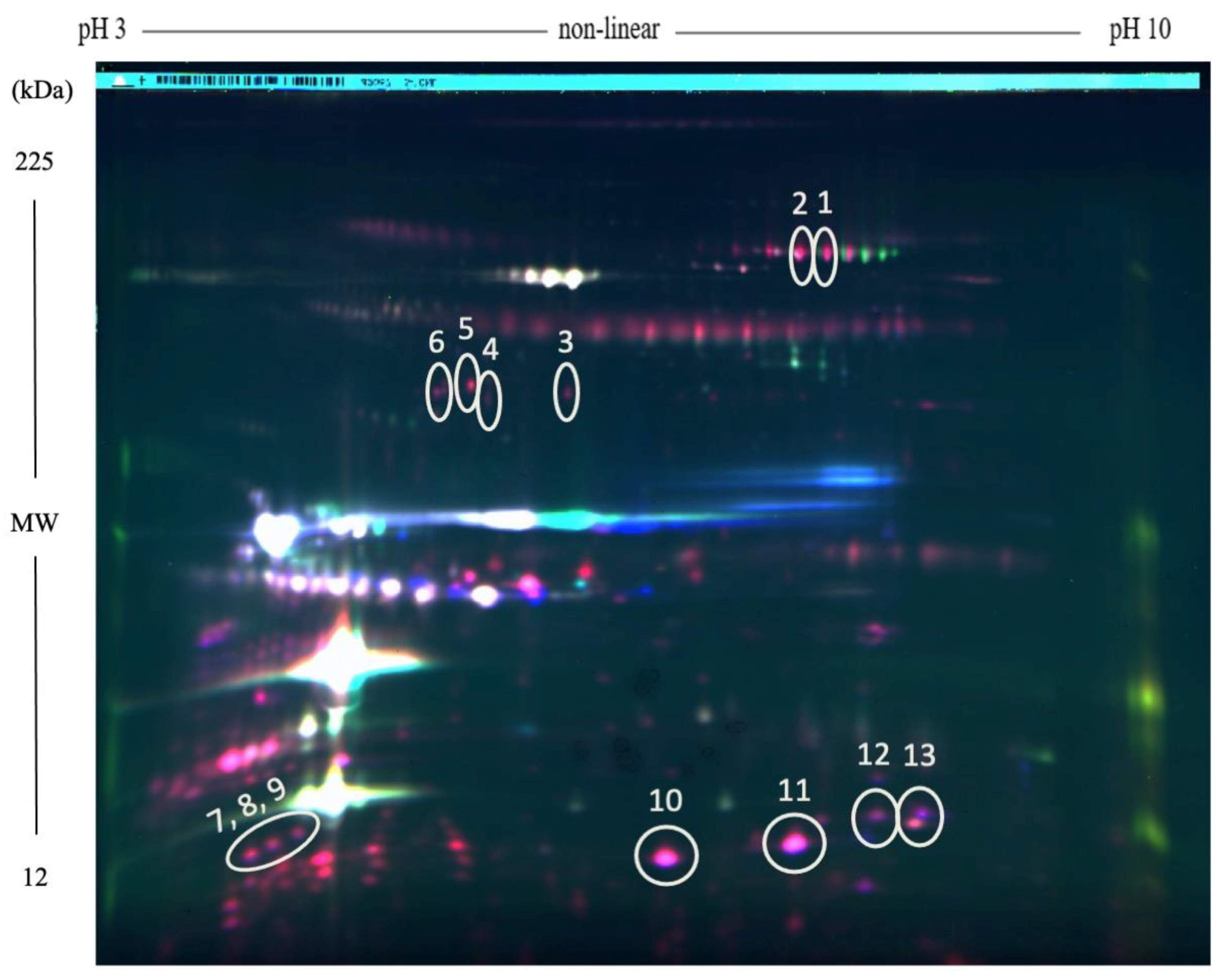{kind=link}
| Function | Protein Name | Accession Number | Gel Spot No. | p-Value | Fold Change Relative to Controls | Mr (kDa) | Unique Peptides | Sequence Coverage (%) |
|---|---|---|---|---|---|---|---|---|
| Host-Defense Proteins | Serotransferrin | Q29443|TRFE_BOVIN | 1 | 0.007 | 3.3 | 77.6 | 3 | 50 |
| Polymeric immunoglobulin receptor | P81265|PIGR_BOVIN | 1 | 0.007 | 3.3 | 82.4 | 12 | 16 | |
| 2 | 0.003 | 5.3 | 82.4 | 13 | 15 | |||
| Fibrinogen gamma-B chain | F1MGU7|F1MGU7_BOVIN | 3 | 0.012 | 3.5 | 50.2 | 22 | 56 | |
| Complement C3 | G3X7A5|G3X7A5_BOVIN | 4 | 0.0001 | 7.5 | 187.1 | 29 | 17.5 | |
| 5 | 0.025 | 6.2 | 187.1 | 43 | 23.7 | |||
| Beta-1,4-galactosyltransferase 1 | P08037|B4GT1_BOVIN | 4 | 0.0001 | 7.5 | 44.8 | 5 | 13.7 | |
| MHC class I antigen | H6V5G4|H6V5G4_BOVIN | 4 | 0.0001 | 7.5 | 38.8 | 6 | 26.3 | |
| Apolipoprotein E | A7YWR0|A7YWR0_BOVIN | 4 | 0.0001 | 7.5 | 35.9 | 4 | 17.7 | |
| Cathepsin B | P07688|CATB_BOVIN | 4 | 0.0001 | 7.5 | 36.6 | 13 | 34.4 | |
| Beta-2-microglobulin | P01888|B2MG_BOVIN | 13 | 0.001 | 20.3 | 13.6 | 5 | 25.4 | |
| Major Milk Proteins | Serum albumin | P02769|ALBU_BOVIN | 3 | 0.012 | 3.5 | 69.2 | 5 | 8.5 |
| 4 | 0.0001 | 7.5 | 69.2 | 27 | 51.6 | |||
| 5 | 0.025 | 6.2 | 69.2 | 15 | 29.8 | |||
| 6 | 0.002 | 11.3 | 69.2 | 11 | 20.8 | |||
| 11 | 0.003 | 13.7 | 69.2 | 7 | 13.7 | |||
| Alpha-S1-casein | P02662|CASA1_BOVIN | 7 | 0.005 | 17.1 | 24.5 | 4 | 32.7 | |
| 8 | 0.002 | 24.7 | 24.5 | 7 | 35 | |||
| 9 | 0.002 | 13.7 | 24.5 | 21 | 48.1 | |||
| Alpha-S2-casein | P02663|CASA2_BOVIN | 6 | 0.002 | 11.3 | 26.0 | 3 | 16.2 | |
| Beta-casein | P02666|CASB_BOVIN | 11 | 0.003 | 13.7 | 25.1 | 5 | 21.4 | |
| 12 | 0.009 | 40.4 | 25.1 | 7 | 25 | |||
| 13 | 0.001 | 20.3 | 25.1 | 6 | 21.9 | |||
| Kappa-casein | P02668|CASK_BOVIN | 4 | 0.0001 | 7.5 | 21.2 | 5 | 30 | |
| 6 | 0.002 | 11.3 | 21.2 | 4 | 25.8 | |||
| Structural and Metabolic proteins | Collagen alpha-1 (I) chain | P02453|CO1A1_BOVIN | 3 | 0.012 | 3.5 | 138.9 | 4 | 3.1 |
| Actin, cytoplasmic 1 | P60712|ACTB_BOVIN | 4 | 0.0001 | 7.5 | 41.7 | 23 | 60 | |
| 5 | 0.025 | 6.2 | 41.7 | 11 | 33.3 | |||
| 6 | 0.002 | 11.3 | 41.7 | 29 | 72.8 | |||
| 7 | 0.005 | 17.1 | 41.7 | 3 | 8.5 | |||
| Actin, cytoplasmic 1 | P60712|ACTB_BOVIN | 10 | 0.002 | 21.9 | 41.7 | 13 | 41.6 | |
| 11 | 0.003 | 13.7 | 41.7 | 6 | 24.3 | |||
| 13 | 0.001 | 20.3 | 41.7 | 6 | 17.9 | |||
| Dystroglycan | F1N7D7|F1N7D7_BOVIN | 4 | 0.0001 | 7.5 | 97.3 | 5 | 7.26 | |
| N-acetylglucosamine-1-phosphotransferase subunit gamma | Q58CS8|GNPTG_BOVIN | 4 | 0.0001 | 7.5 | 33.7 | 5 | 19.3 | |
| Hemoglobin subunit alpha | P01966|HBA_BOVIN | 10 | 0.002 | 21.9 | 15.1 | 4 | 31.7 | |
| Thymosin beta-4 | P62326|TYB4_BOVIN | 10 | 0.002 | 21.9 | 5 | 3 | 54.5 | |
| Histone H2B | F1MUD2|F1MUD2_BOVIN | 10 | 0.002 | 21.9 | 13.9 | 3 | 38.1 | |
| Histone H1.2 | P02253|H12_BOVIN | 10 | 0.002 | 21.9 | 21.3 | 3 | 11.7 | |
| Unknown | Putative uncharacterized protein | A5D7Q2|A5D7Q2_BOVIN | 4 | 0.0001 | 7.5 | 51.6 | 4 | 9.65 |
| Spot Number | Protein Name | Protein Accession Numbers | Unique Peptide Count | Sequence Coverage |
|---|---|---|---|---|
| A | 50S ribosomal protein L19 | RL19_STAA1 | 2 | 17.20% |
| A | Orotate phosphoribosyltransferase | PYRE_STAAC,PYRE_STAAS | 2 | 10.80% |
| A | Immunoglobulin G-binding protein A | SPA_STAA8 | 8 | 23.10% |
| B | ATP-dependent Clp protease ATP-binding subunit ClpL | CLPL_STAAR | 5 | 16.00% |
| B | DNA topoisomerase 4 subunit B | PARE_STAEQ | 5 | 13.30% |
| B | Hyaluronate lyase | HYSA_STAA8 | 7 | 13.40% |
| B | Putative hemin import ATP-binding protein HrtA | HRTA_STAAR | 5 | 24.40% |
| B | Septation ring formation regulator EzrA | EZRA_STAEQ | 5 | 12.60% |
| B | Protein draper | DRPR_DROME | 6 | 16.10% |
| C | DNA-directed RNA polymerase subunit beta (Fragment) | RPOC_STAAU | 5 | 12.10% |
| C | 60 kDa chaperonin | CH60_STAAU | 5 | 20.40% |
| D | Immunoglobulin G-binding protein A | SPA_STAA8 | 10 | 26.60% |
| D | Staphopain B | SSPB_STAAS | 2 | 10.90% |
| D | Transcription termination/antitermination protein NusG | NUSG_STAA8 | 2 | 14.30% |
| D | Lactose phosphotransferase system repressor | LACR_STAA8, LACR_STAAR | 2 | 18.30% |
| D | Acetylglutamate kinase | ARGB_STAAR | 2 | 22.70% |
| Categories | Biological Processes | Count 1 | p-Value 2 |
|---|---|---|---|
| Cellular form | GO:0044085~cellular component biogenesis | 5 | 0.009 |
| GO:0065003~macromolecular complex assembly | 4 | 0.017 | |
| GO:0016043~cellular component organization | 8 | 0.002 | |
| GO:0022607~cellular component assembly | 5 | 0.005 | |
| GO:0009653~anatomical structure morphogenesis | 5 | 0.008 | |
| GO:0043933~macromolecular complex subunit organization | 4 | 0.02 | |
| GO:0051270~regulation of cell motion | 3 | 0.008 | |
| GO:0048856~anatomical structure development | 6 | 0.021 | |
| GO:0048646~anatomical structure formation involved in morphogenesis | 3 | 0.033 | |
| GO:0051651~maintenance of location in cell | 2 | 0.028 | |
| GO:0051235~maintenance of location | 3 | 0.001 | |
| Regulation of cellular process | GO:0032879~regulation of localization | 4 | 0.008 |
| GO:0051179~localization | 9 | 0.011 | |
| GO:0048523~negative regulation of cellular process | 5 | 0.021 | |
| GO:0048519~negative regulation of biological process | 5 | 0.03 | |
| GO:0065008~regulation of biological quality | 5 | 0.029 | |
| GO:0051049~regulation of transport | 3 | 0.037 | |
| GO:0001649~osteoblast differentiation | 2 | 0.038 | |
| Cell Death | GO:0042981~regulation of apoptosis | 4 | 0.017 |
| GO:0043065~positive regulation of apoptosis | 3 | 0.022 | |
| GO:0043068~positive regulation of programmed cell death | 3 | 0.022 | |
| GO:0010942~positive regulation of cell death | 3 | 0.022 | |
| GO:0043067~regulation of programmed cell death | 4 | 0.018 | |
| GO:0010941~regulation of cell death | 4 | 0.018 | |
| Response to stimulus | GO:0009605~response to external stimulus | 4 | 0.019 |
| GO:0050896~response to stimulus | 8 | 0.005 | |
| GO:0006950~response to stress | 6 | 0.006 | |
| GO:0009611~response to wounding | 3 | 0.043 | |
| Angiogenesis | GO:0001944~vasculature development | 3 | 0.022 |
| GO:0001568~blood vessel development | 3 | 0.021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelmegid, S.; Kelton, D.; Caswell, J.; Kirby, G. Proteomic 2D-DIGE Analysis of Milk Whey from Dairy Cows with Staphylococcus aureus Mastitis Reveals Overexpression of Host Defense Proteins. Microorganisms 2020, 8, 1883. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121883
Abdelmegid S, Kelton D, Caswell J, Kirby G. Proteomic 2D-DIGE Analysis of Milk Whey from Dairy Cows with Staphylococcus aureus Mastitis Reveals Overexpression of Host Defense Proteins. Microorganisms. 2020; 8(12):1883. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121883
Chicago/Turabian StyleAbdelmegid, Shaimaa, David Kelton, Jeff Caswell, and Gordon Kirby. 2020. "Proteomic 2D-DIGE Analysis of Milk Whey from Dairy Cows with Staphylococcus aureus Mastitis Reveals Overexpression of Host Defense Proteins" Microorganisms 8, no. 12: 1883. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121883