Contrasting Environmental Drivers Determine Biodiversity Patterns in Epiphytic Lichen Communities along a European Gradient

 ,
,  , , ,
, , ,  , , , , , , , ,
, , , , , , , , 
Abstract
:
1. Introduction
2. Materials and Methods
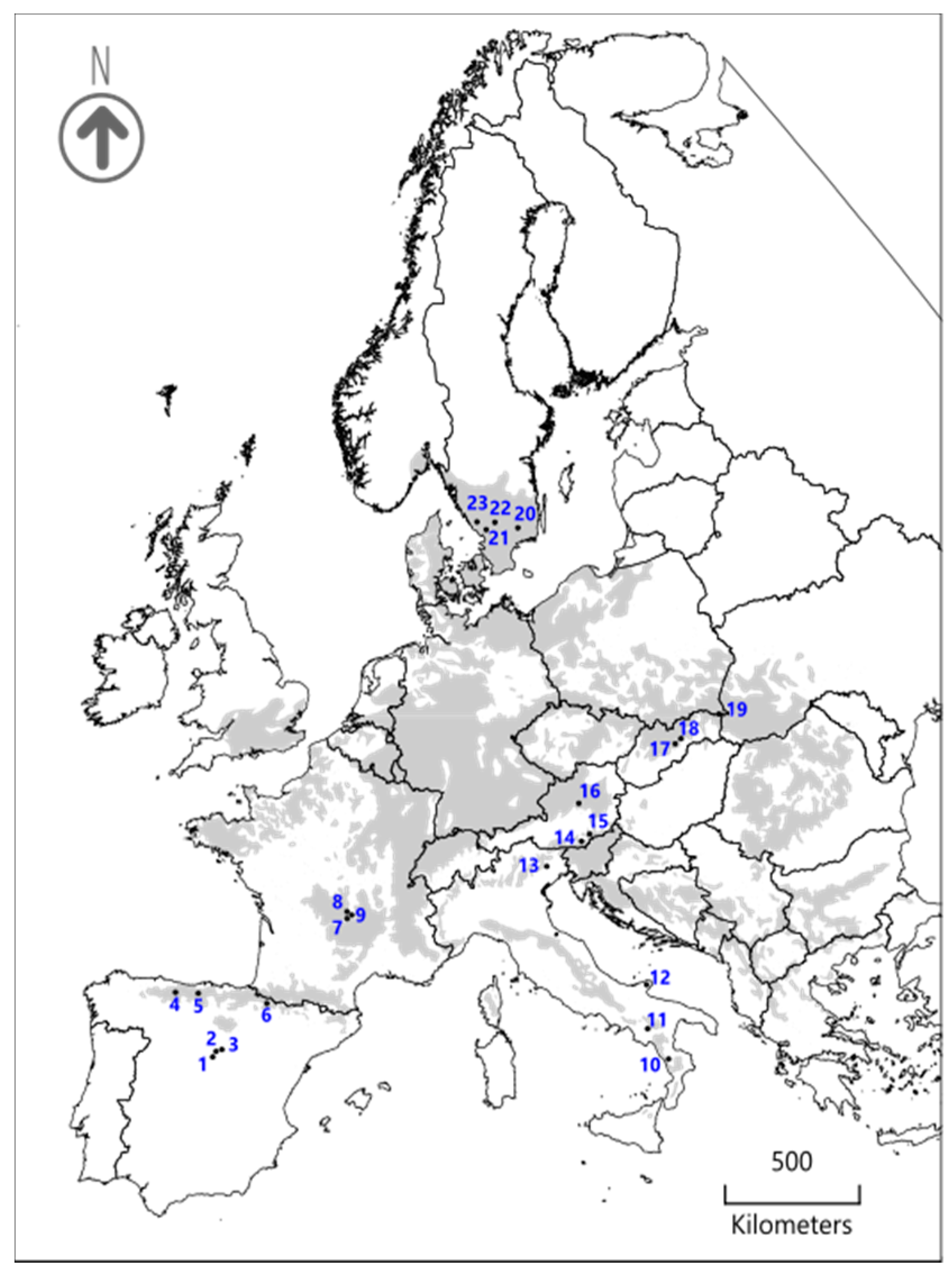
2.1. Study Area and Sampling Design
2.2. Trait Data and Phylogenetic Tree
2.3. Diversity Metrics
2.4. Environmental Drivers
2.5. Data Analyses
2.5.1. Environmental Driver Contribution and Relationships between TD, FD, and PD Determining Biodiversity Patterns across Europe
2.5.2. Contribution of Climatic Drivers Determining Species Composition and Functional Structure of Lichen Communities across Europe
3. Results
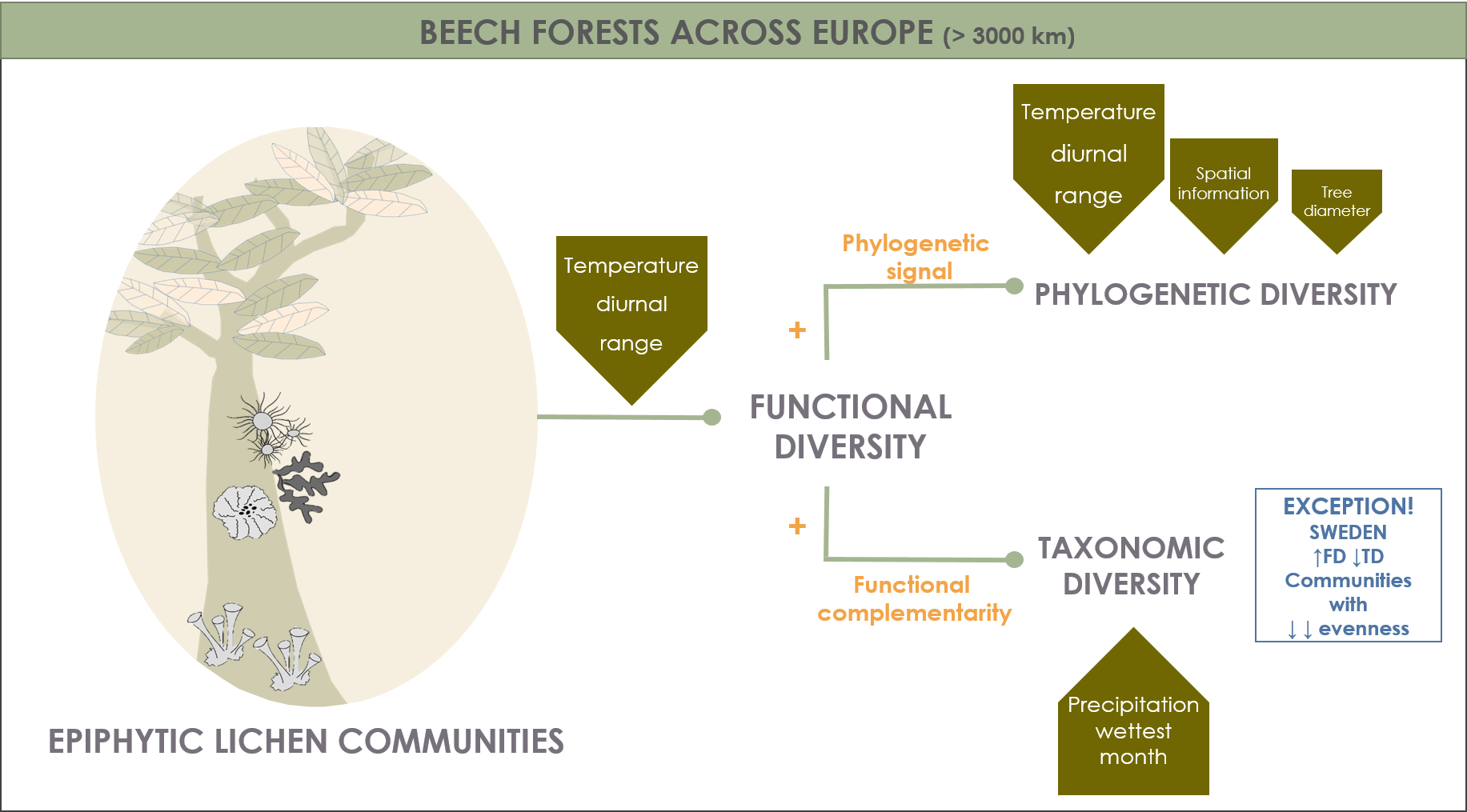
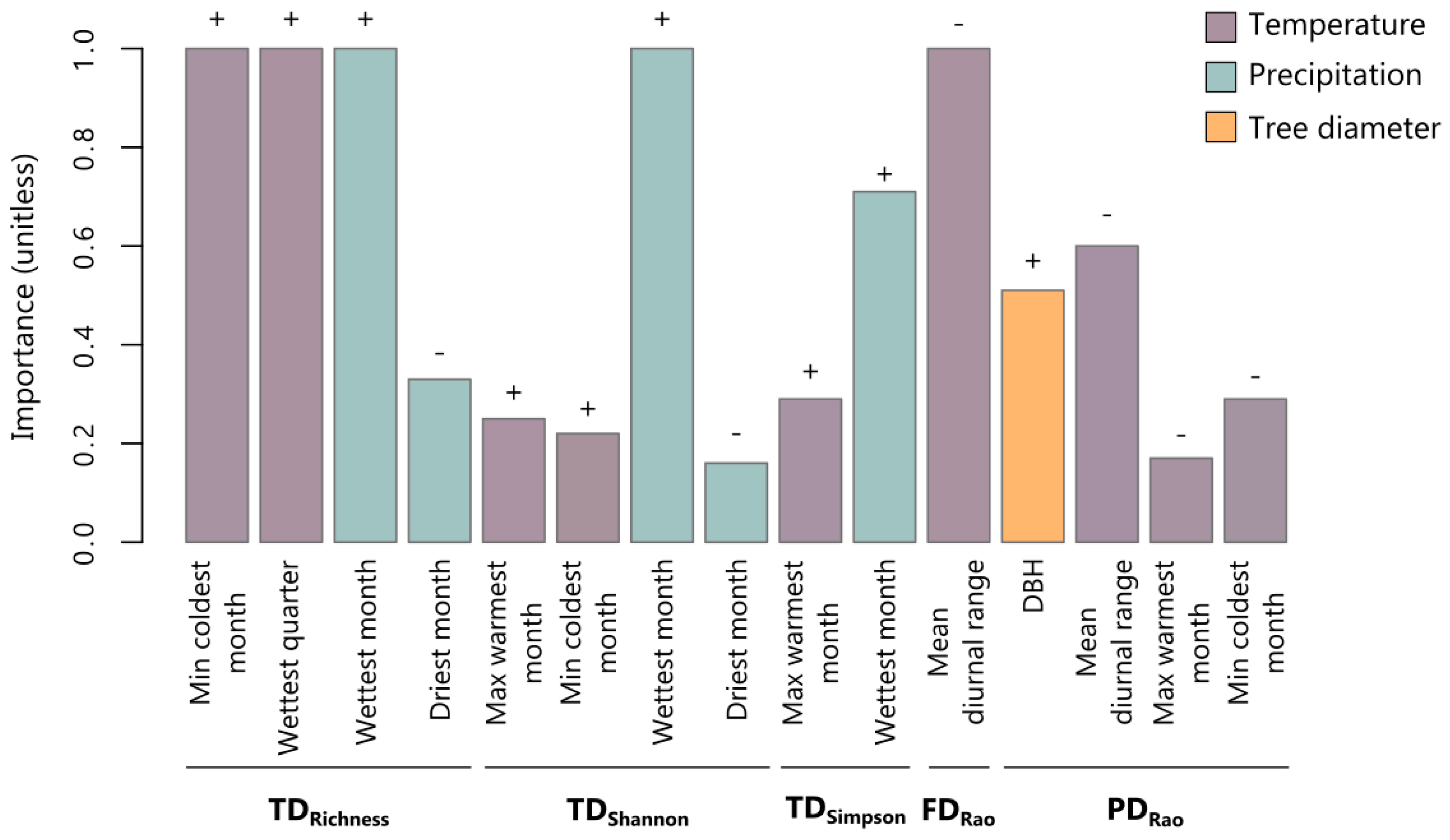
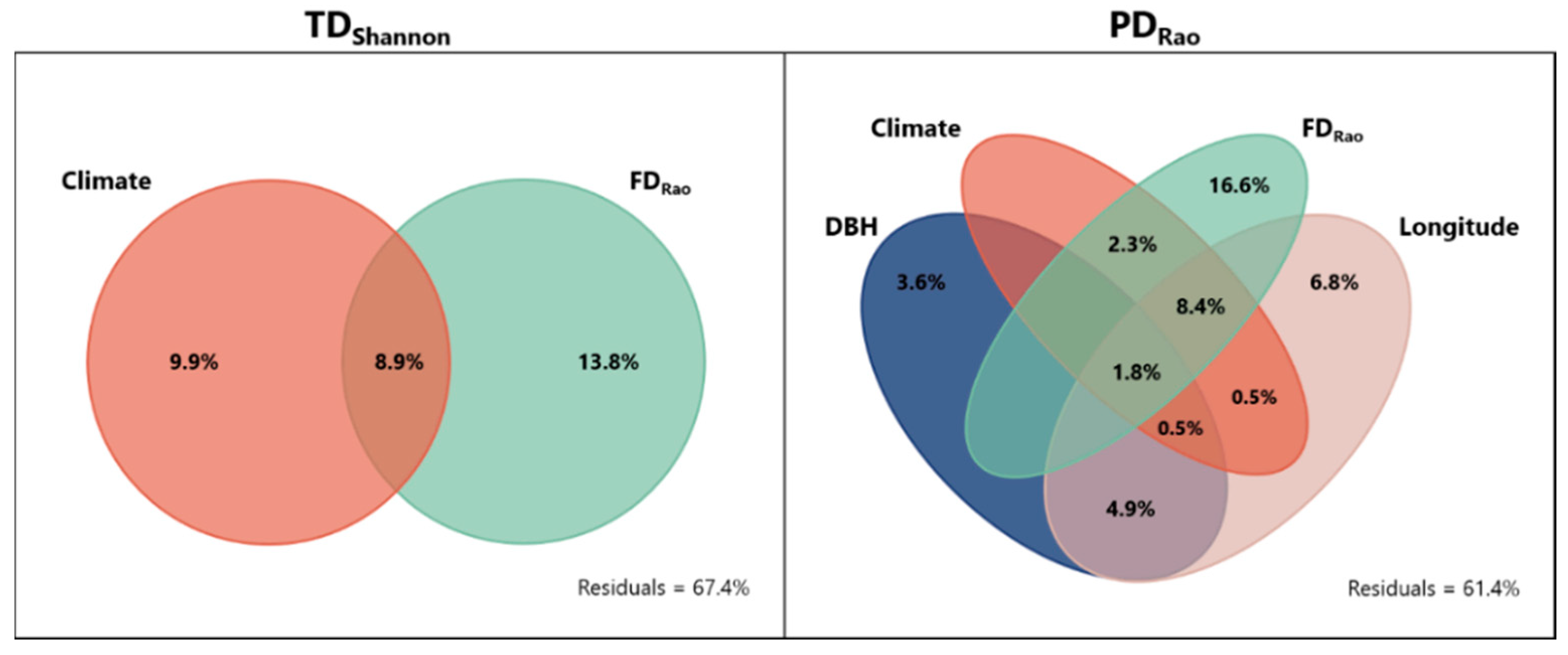
3.1. Environmental Driver Contribution and the Relationships between TD, FD, and PD Determining the Biodiversity Patterns across Europe
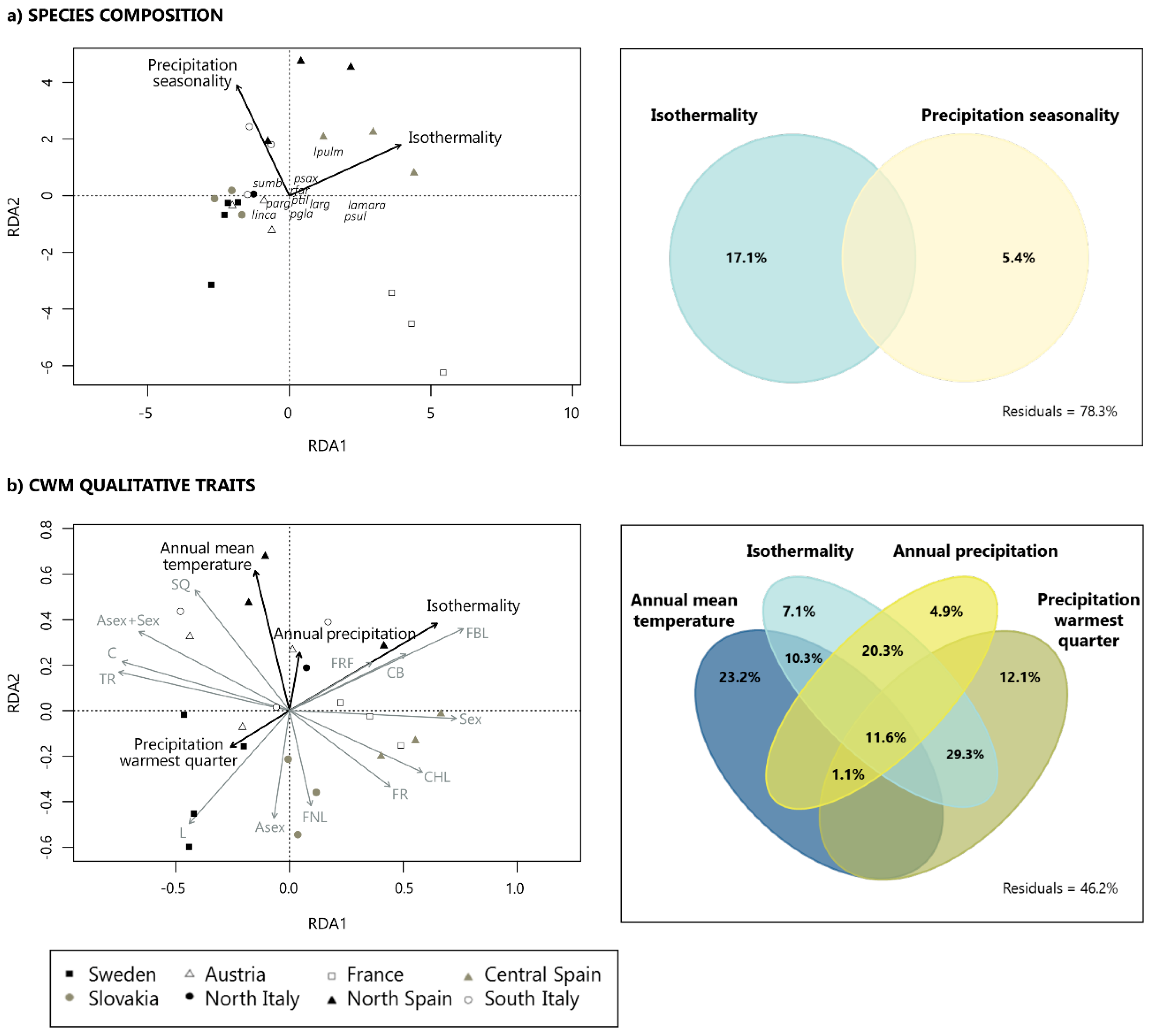
3.2. Contribution of Climatic Drivers Determining Species Composition and Functional Structure of Lichen Communities across Europe
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Mittelbach, G.G. Community Ecology, 1st ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2012; p. 400. [Google Scholar]
- Hurtado, P.; Prieto, M.; Aragón, G.; Escudero, A.; Martínez, I. Critical predictors of functional, phylogenetic and taxonomic diversity are geographically structured in lichen epiphytic communities. J. Ecol. 2019, 107, 2303–2316. [Google Scholar] [CrossRef]
- Hurtado, P.; Matos, P.; Aragón, G.; Branquinho, C.; Prieto, M.; Martínez, I. How much matching there is in functional, phylogenetic and taxonomic optima of epiphytic macrolichen communities along a European climatic gradient? Sci. Total Environ. 2020, 712, 136533. [Google Scholar] [CrossRef]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet, F.; Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Swenson, N.G. The role of evolutionary processes in producing biodiversity patterns, and the interrelationships between taxonomic, functional and phylogenetic biodiversity. Am. J. Bot. 2011, 98, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Purschke, O.; Schmid, B.C.; Sykes, M.T.; Poschlod, P.; Michalski, S.G.; Durka, W.; Kühn, I.; Winter, M.; Prentice, H.C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes. J. Ecol. 2013, 101, 857–866. [Google Scholar] [CrossRef]
- Losos, J.B. Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecol. Lett. 2008, 11, 995–1003. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Fetzer, I.; Johst, K.; Schäwe, R.; Banitz, T.; Harms, H.; Chatzinotas, A. The extent of functional redundancy changes as species’ roles shift in different environments. Proc. Natl. Acad. Sci. USA 2015, 112, 14888–14893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forest, F.; Grenyer, R.; Rouget, M.; Davies, T.J.; Cowling, R.M.; Faith, D.P.; Balmford, A.; Manning, J.C.; Procheş, S.; van der Bank, M.; et al. Preserving the evolutionary potential of floras in biodiversity hotspots. Nature 2007, 445, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.M.; Cadotte, M.W. Unifying measures of biodiversity: Understanding when richness and phylogenetic diversity should be congruent. Divers. Distrib. 2013, 19, 1–10. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Evol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef] [Green Version]
- Leão-Pires, T.A.; Luiz, A.M.; Sawaya, R.J. The complex roles of space and environment in structuring functional, taxonomic and phylogenetic beta diversity of frogs in the Atlantic Forest. PLoS ONE 2018, 13, e0196066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.H.; Lee, C.B. Disentangling the local-scale drivers of taxonomic, phylogenetic and functional diversity in woody plant assemblages along elevational gradients in South Korea. PLoS ONE 2017, 12, e0185763. [Google Scholar] [CrossRef] [Green Version]
- Safi, K.; Cianciaruso, M.V.; Loyola, R.D.; Brito, D.; Armour-Marshall, K.; DinizFilho, J.A.F. Understanding global patterns of mammalian functional and phylogenetic diversity. Philos. Trans. R. Soc. B 2011, 366, 2536–2544. [Google Scholar] [CrossRef]
- Green, T.A.; Sancho, L.G.; Pintado, A. Ecophysiology of desiccation/rehydration cycles in mosses and lichens. In Plant Desiccation Tolerance, 1st ed.; Luttge, U., Beck, E., Pagel, M.D., Eds.; Springer: Berlin, Germany, 2011; p. 386. [Google Scholar]
- Matos, P.; Pinho, P.; Aragón, G.; Martínez, I.; Nunes, A.; Soares, A.M.V.M.; Branquinho, C. Lichen traits responding to aridity. J. Ecol. 2015, 103, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Giordani, P.; Brunialti, G.; Bacaro, G.; Nascimbene, J. Functional traits of epiphytic lichens as potential indicators of environmental conditions in forest ecosystems. Ecol. Indic. 2012, 18, 413–420. [Google Scholar] [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Aragón, G.; Martínez, I.; García, A. Loss of epiphytic diversity along a latitudinal gradient in Southern Europe. Sci. Total Environ. 2012, 426, 188–195. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L. Epiphytic lichen diversity along elevational gradients: Biological traits reveal a complex response to water and energy. J. Biogeogr. 2015, 42, 1222–1232. [Google Scholar] [CrossRef]
- Prieto, M.; Martínez, I.; Aragón, G.; Verdú, M. Phylogenetic and functional structure of lichen communities under contrasting environmental conditions. J. Veg. Sci. 2017, 28, 871–881. [Google Scholar] [CrossRef]
- Nimis, P.L.; Martellos, S. ITALIC—The Information System on Italian Lichens, Version 5.0; Dept. of Biology, University of Trieste: Trieste, Italy, 2017; Available online: http://dryades.units.it/italic (accessed on 20 December 2017).
- Rambold, G.; Elix, J.A.; Heindl-Tenhunen, B.; Köhler, T.; Nash, T.H.; Neubacher, D.; Reichert, W.; Zedda, L.; Triebel, D. LIAS light-Towards the ten thousand species milestone. MycoKeys 2014, 8, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Merinero, S.; Hilmo, O.; Gauslaa, Y. Size is a main driver for hydration traits in cyano- and cephalolichens of boreal rainforest canopies. Fungal Ecol. 2014, 7, 59–66. [Google Scholar] [CrossRef]
- Prieto, M.; Wedin, M. Dating the diversification of the major lineages of Ascomycota (Fungi). PLoS ONE 2013, 8, e65576. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R, 1st ed.; Springer: New York, NY, USA, 2009; p. 574. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K. Multimodel Inference: Understanding AIC Relative Variable Importance Values. 2015. Available online: https://sites.warnercnr.colostate.edu/ (accessed on 18 November 2018).
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Lepš, J.; Smilauer, P. Multivariate Analysis of Ecological Data Using CANOCO, 1st ed.; Cambridge University Press: Cambridge, UK, 2003; p. 282. [Google Scholar]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P.A. Assembly rules, null models, and trait dispersion—New questions front old patterns. Oikos 1995, 74, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M.; et al. Ecological assembly rules in plant communities—Approaches, patterns and prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity (FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial community evenness favours functionality under selective stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef]
- Pausas, J.G.; Verdú, M. The jungle of methods for evaluating phenotypic and phylogenetic structure of communities. Bioscience 2010, 60, 614–625. [Google Scholar] [CrossRef] [Green Version]
- Swenson, N.G.; Enquist, B.J. Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community-wide variation across latitude and elevation. Am. J. Bot. 2007, 94, 451–459. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Davies, T.J.; Regetz, J.; Kembel, S.W.; Cleland, E.E.; Oakley, T.H. Phylogenetic diversity metrics for ecological communities: Integrating species richness, abundance and evolutionary history. Ecol. Lett. 2010, 13, 96–105. [Google Scholar] [CrossRef]
- Riiali, A.; Penttinen, A.; Kuusinen, M. Bayesian mapping of lichens growing on trees. Biom. J. 2001, 43, 717–736. [Google Scholar] [CrossRef]
- Tehler, A.; Irestedt, M. Parallel evolution of lichen growth forms in the family Roccellaceae (Arthoniales, Ascomycota). Cladistics 2007, 23, 432–454. [Google Scholar] [CrossRef]
- Magri, D. Patterns of post-glacial spread and the extent of glacial refugia of European beech (Fagus sylvatica). J. Biogeogr. 2008, 35, 450–463. [Google Scholar] [CrossRef]
- Shipley, B.; de Bello, F.; Cornelissen, H.; Laliberté, E.; Laughlin, D.; Reich, P. Reinforcing foundation stones in trait-based plant ecology. Oecologia 2016, 180, 923–931. [Google Scholar] [CrossRef]
- Mackenzie, T.D.B.; MacDonald, T.M.; Dubois, L.A.; Campbell, D.A. Seasonal changes in temperature and light drive acclimation of photosynthetic physiology and macromolecular content in Lobaria pulmonaria. Planta 2001, 214, 57–66. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FDRao | PDRao | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Linear | Quadratic | Linear | Quadratic | R2 | AICc | ΔAICc | wi | Ip | |
| TDRichness | 0 | 180.1 | 0 | 1 | - | ||||
| TDShannon | 0.30 | −3 | 0 | 1 | 0.58 | ||||
| TDInverse Simpson | 0.27 | 104.6 | 0 | 0.68 | 0.61 | ||||
| 0 | 106.1 | 1.52 | 0.32 | - | |||||
| PDRao | - | - | 0.29 | −20 | 0 | 1 | 0.004 * | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurtado, P.; Prieto, M.; de Bello, F.; Aragón, G.; López-Angulo, J.; Giordani, P.; Díaz-Peña, E.M.; Vicente, R.; Merinero, S.; Košuthová, A.; et al. Contrasting Environmental Drivers Determine Biodiversity Patterns in Epiphytic Lichen Communities along a European Gradient. Microorganisms 2020, 8, 1913. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121913
Hurtado P, Prieto M, de Bello F, Aragón G, López-Angulo J, Giordani P, Díaz-Peña EM, Vicente R, Merinero S, Košuthová A, et al. Contrasting Environmental Drivers Determine Biodiversity Patterns in Epiphytic Lichen Communities along a European Gradient. Microorganisms. 2020; 8(12):1913. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121913
Chicago/Turabian StyleHurtado, Pilar, María Prieto, Francesco de Bello, Gregorio Aragón, Jesús López-Angulo, Paolo Giordani, Eva María Díaz-Peña, Rebeca Vicente, Sonia Merinero, Alica Košuthová, and et al. 2020. "Contrasting Environmental Drivers Determine Biodiversity Patterns in Epiphytic Lichen Communities along a European Gradient" Microorganisms 8, no. 12: 1913. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8121913