Aspergillosis, Avian Species and the One Health Perspective: The Possible Importance of Birds in Azole Resistance

 ,
,  and
and 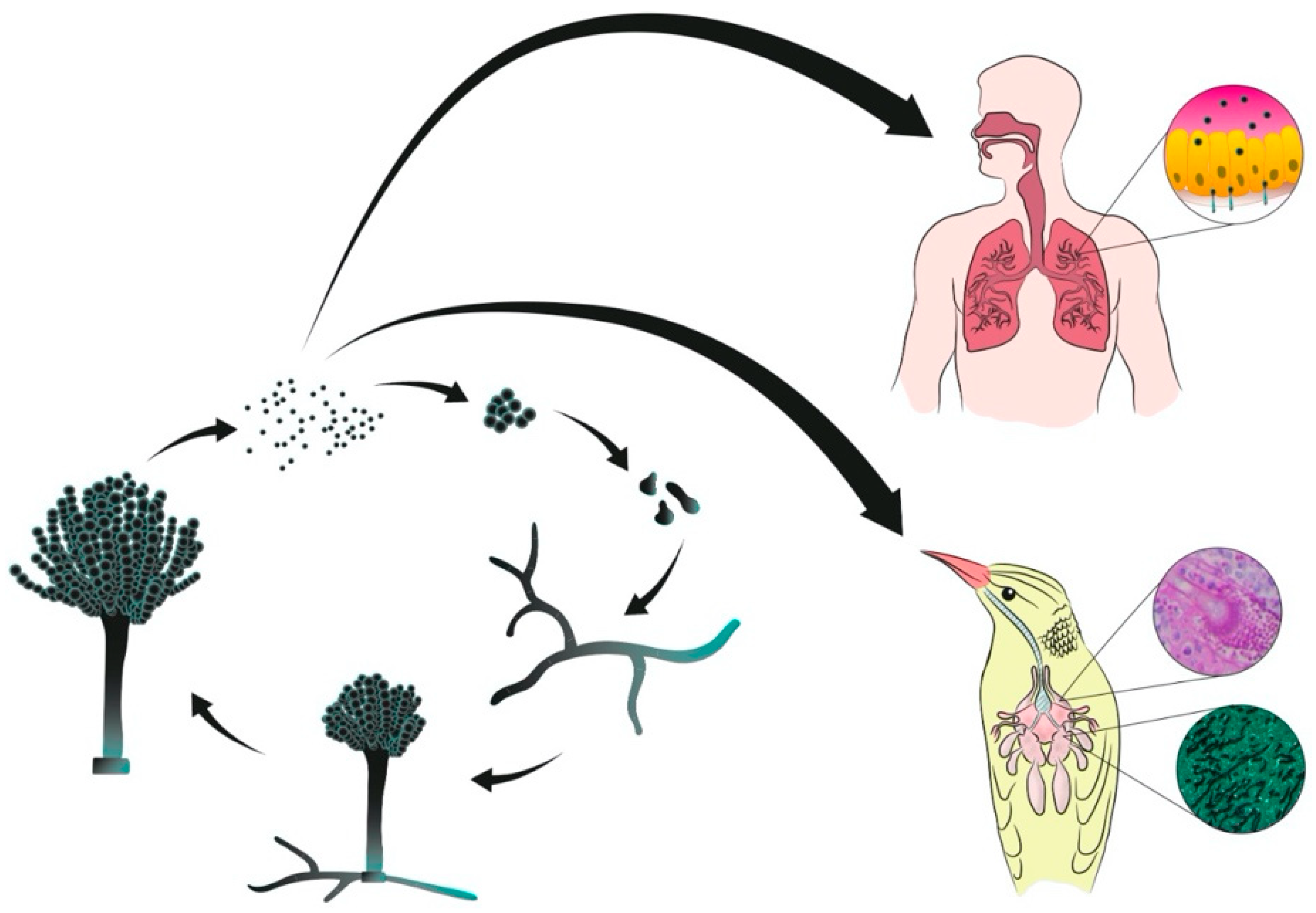{kind=link}
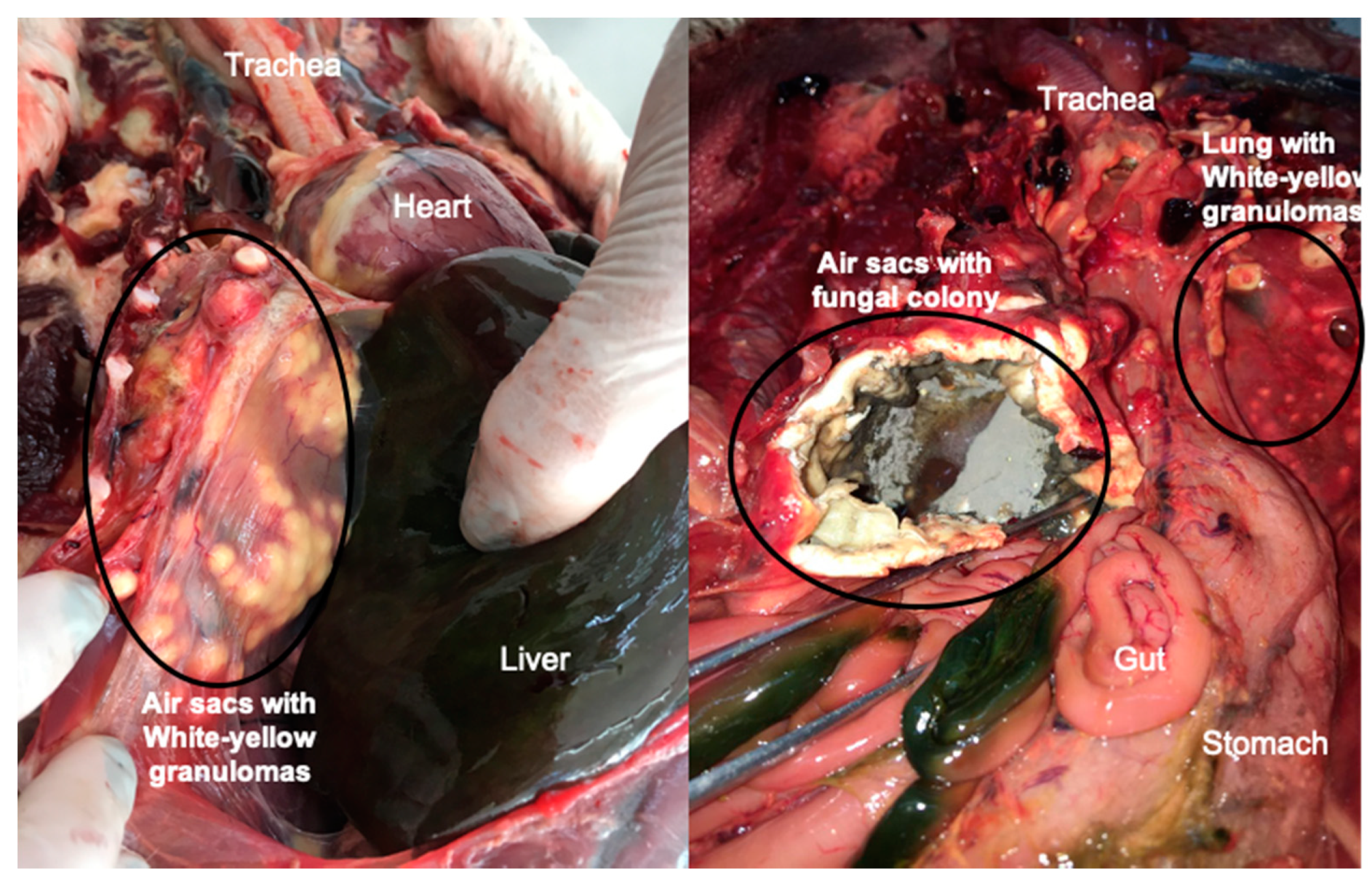{kind=link}
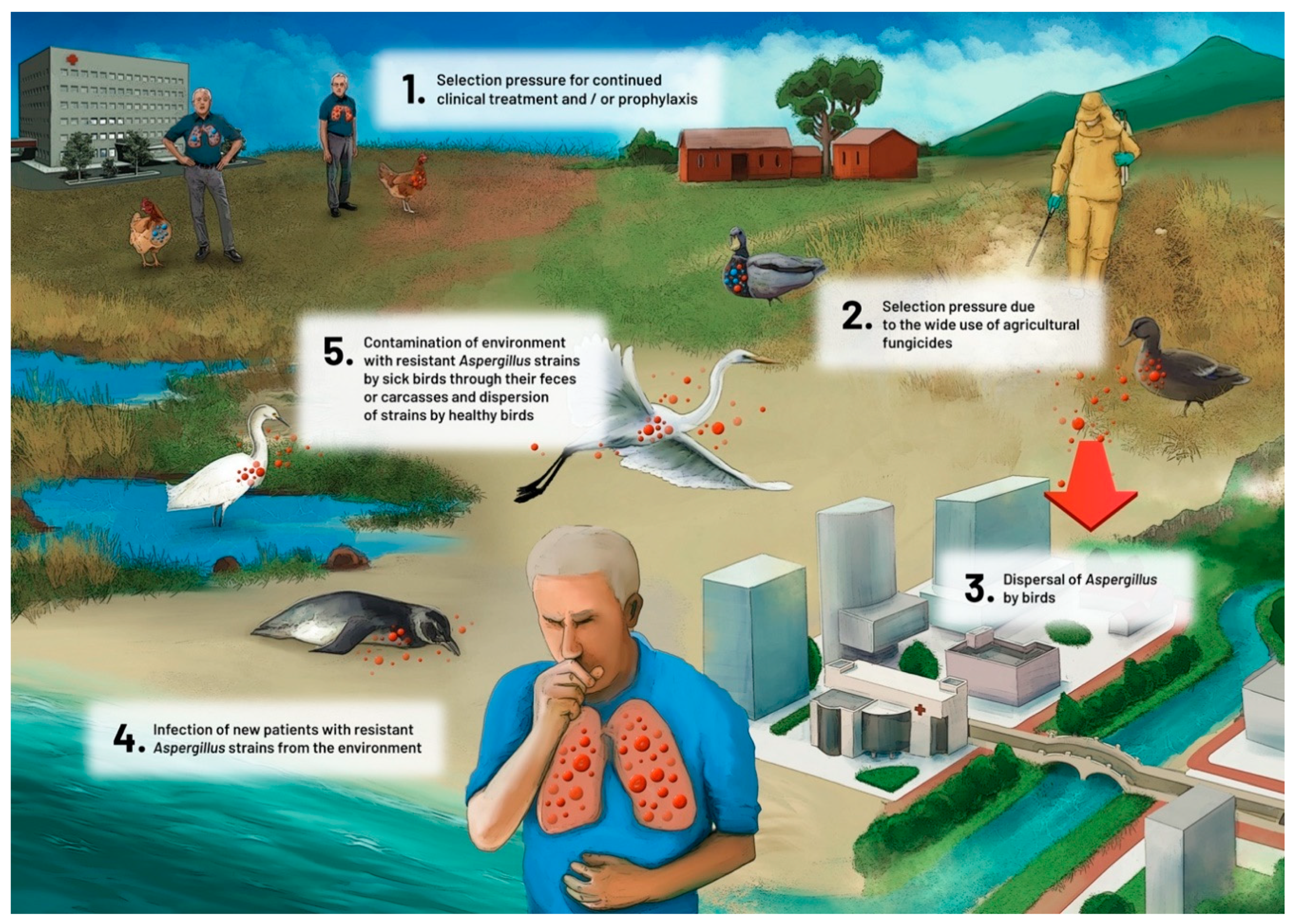{kind=link}
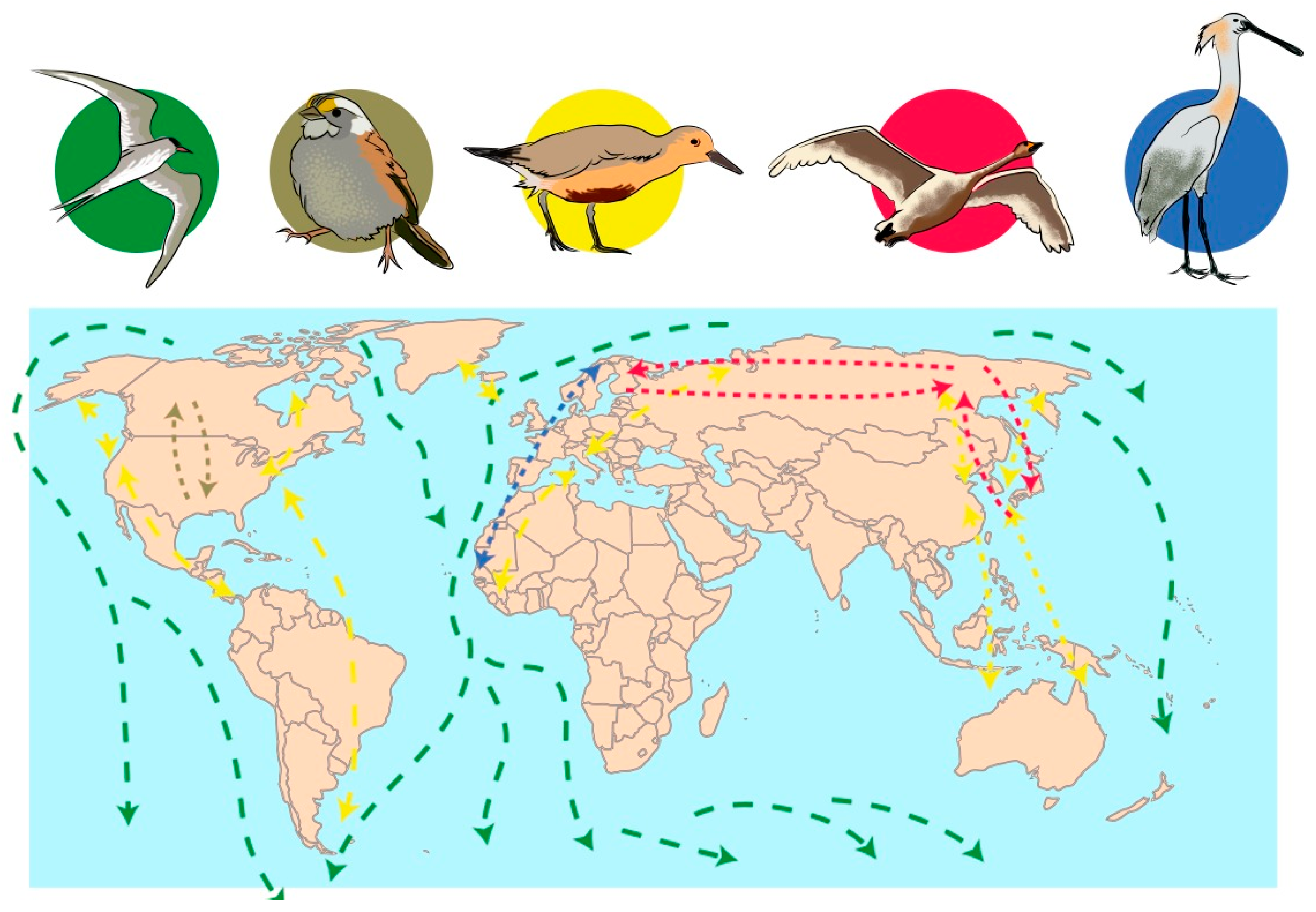{kind=link}
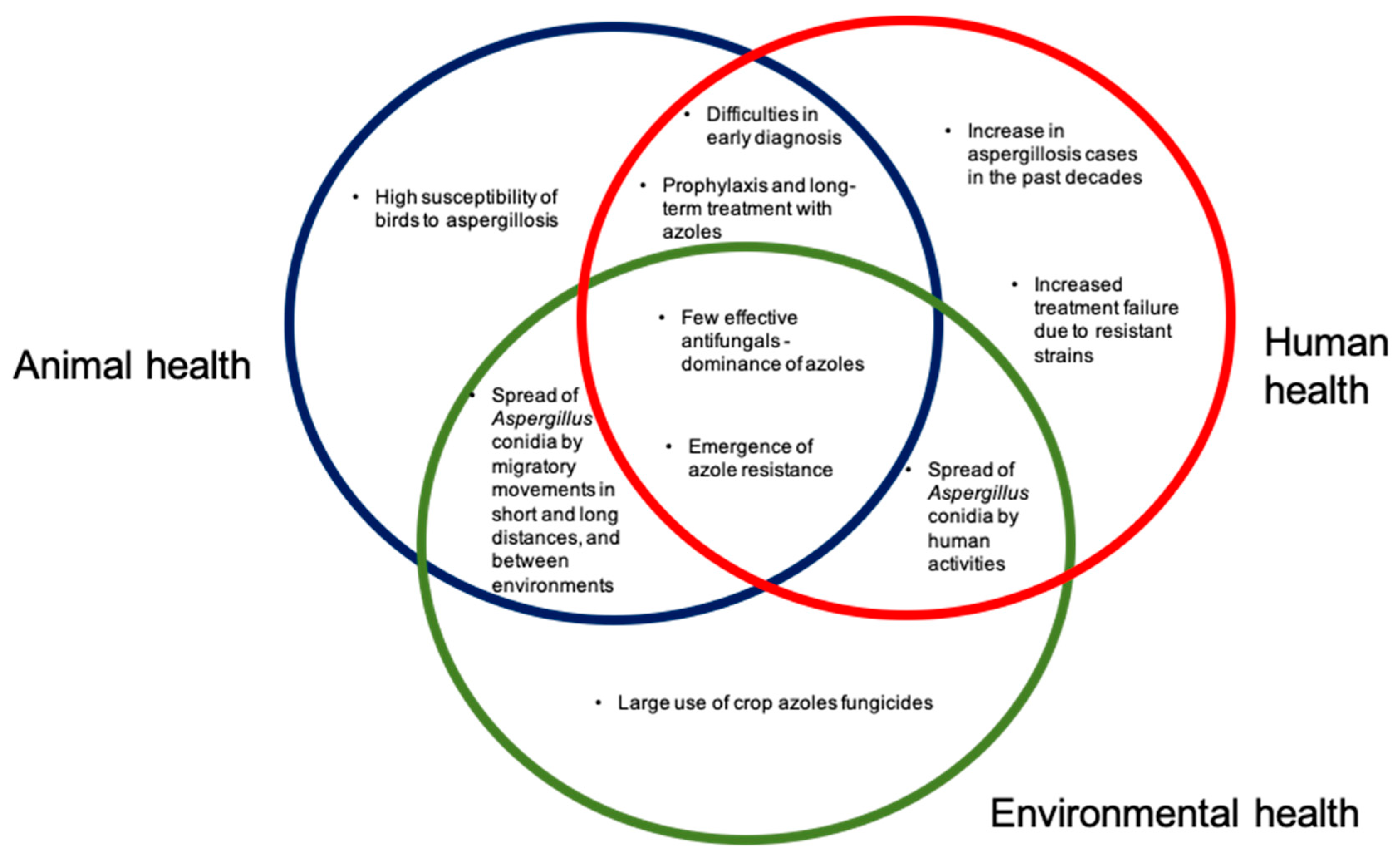{kind=link}
Abstract
:1. Introduction
2. Aspergillus
3. Aspergillosis in Humans
4. Aspergillosis in Avian Species
5. Aspergillus Azole Resistance
6. Main Mechanisms of Azole Resistance
7. The Role of Pesticides in Emerging Aspergillus Azole Resistance
8. Contamination of the Environment with Pesticides
9. Use of Crop Areas and Adjacent Areas by Birds
10. Bird Migration and Their Role in Pathogen Dispersion
11. Why Should We Consider the Role of Birds in Azole Resistance in the One Health Approach?
12. Concluding Remarks and Future Perspectives
- Aspergillus species are ubiquitous fungi, present in many environments, and it is a potential pathogen of importance in animals (we emphasize birds) and humans;
- The emergence of azole resistance in Aspergillus species such as A. fumigatus sensu stricto is a major concern, limiting treatment success;
- The wide use of crop fungicides has an important role in the emergence of azole resistance;
- Avian species are highly susceptible to aspergillosis. Many species have regular migratory movements, moving between different environments, including those where large amounts of pesticides are used, and natural environments, such as estuaries, lagoons and beaches;
- In their movements between different types of environments, such as agricultural fields, natural environments and urban environments, birds may have an important role in the dispersion of Aspergillus isolates, especially resistant strains;
- Considering the characteristics of Aspergillus fungi, the importance of aspergillosis in birds and humans, and the emergence of azole resistance, it is essential to promote more investigations in a One Health approach.
- Avian aspergillosis can be used as an indicator of environmental health in specific countries or regions, including surveillance of the introduction of new resistant strains, changes in resistance rates, and emergence of new mechanisms of azole resistance.
- Regular cleaning of where birds are kept, with non-azole products, aiming at environmental control of the amount of fungal inoculum;
- Avoid contact of people, in groups at risk for aspergillosis, with poultry farms, zoological institutions, avian wildlife rehabilitation centers;
- Search for more efficient early diagnosis techniques for both humans and birds;
- Implementation of antifungal stewardship programs for both humans and birds;
- Search for new antifungal molecules different from those presently used on crops, with different mechanisms of action, and dissimilar to those used in human therapy;
- Surveillance of antifungal susceptibility in Aspergillus strains in environment, birds, and humans;
- Measures to control the dispersion of Aspergillus strains in agricultural products transported by humans from farms to urban areas;
- Surveillance of the main routes of bird migration and correlation with spread of azole resistant isolates;
- Search for new fungal control options for fungal control for crops, such as biological rather than chemical control.
Author Contributions
Funding
Conflicts of Interest
References
- Seyedmousavi, S.; Guillot, J.; Arné, P.; De Hoog, G.S.; Mouton, J.W.; Melchers, W.J.G.; Verweij, P.E. Aspergillus and aspergillosis in wild and domestic animals: A global health concern with parallels to human disease. Med. Mycol. 2015, 53, 765–797. [Google Scholar] [PubMed]
- Marr, K.A.; Patterson, T.; Denning, D. Aspergillosis pathogenesis, clinical manifestations, and therapy. Infect. Dis. Clin. N. Am. 2002, 16, 875–894. [Google Scholar]
- Engel, T.G.P.; Erren, E.; Vanden Driessche, K.S.J.; Melchers, W.J.G.; Reijers, M.H.; Merkus, P.; Verweij, P.E. Aerosol transmission of Aspergillus fumigatus in cystic fibrosis patients in the Netherlands. Emerg. Infect. Dis. 2019, 25, 797–799. [Google Scholar] [PubMed] [Green Version]
- Lemaire, B.; Normand, A.-C.; Forel, J.-M.; Cassir, N.; Piarroux, R.; Ranque, S. Hospitalized patient as source of Aspergillus fumigatus. Emerg. Infect. Dis. 2018, 24, 1524–1527. [Google Scholar]
- Cacciuttolo, E.; Rossi, G.; Nardoni, S.; Legrottaglie, R.; Mani, P. Anatomopathological aspects of avian aspergillosis. Vet. Res. Commun. 2009, 33, 521–527. [Google Scholar] [PubMed]
- Xavier, M.O.; Soares, M.P.; Silva Filho, R.P.; Ruoppolo, V.; Pasqualotto, A.C.; Meireles, M.C.A.; Severo, L.C. Clinical and pathological findings of aspergillosis in Magellanic penguins (Spheniscus magellanicus). Ciência Anim. Bras. 2011, 12, 520–524. [Google Scholar]
- Tang, X.D.; Li, G.H. Treatment of aspergillosis: Clinical practice guidelines of Infectious Diseases Society of America. Chin. J. Infect. Chemother. 2008, 8, 161–166. [Google Scholar]
- Bassetti, M.; Pecori, D.; Della Siega, P.; Corcione, S.; De Rosa, F.G. Current and future therapies for invasive aspergillosis. Pulm. Pharmacol. Ther. 2015, 32, 155–165. [Google Scholar]
- Hyatt, M.W.; Georoff, T.A.; Nollens, H.H.; Wells, R.L.; Clauss, T.M.; Ialeggio, D.M.; Harms, C.A.; Wack, A.N. Voriconazole toxicity in multiple penguin species. J. Zoo Wildl. Med. 2015, 46, 880–888. [Google Scholar]
- Bunskoek, P.E.; Seyedmousavi, S.; Gans, S.J.M.; van Vierzen, P.B.J.; Melchers, W.J.G.; van Elk, C.E.; Mouton, J.W.; Verweij, P.E. Successful treatment of azole-resistant invasive aspergillosis in a bottlenose dolphin with high-dose posaconazole. Med. Mycol. Case Rep. 2017, 16, 16–19. [Google Scholar]
- Bunting, E.M.; Abou-Madi, N.; Cox, S.; Martin-Jimenez, T.; Fox, H.; Kollias, G.V. Evaluation of oral itraconazole administration in captive Humboldt penguins (Spheniscus humboldti). J. Zoo Wildl. Med. 2009, 40, 508–518. [Google Scholar] [PubMed]
- Da Silva Filho, R.P.; Xavier, M.O.; Martins, A.M.; Ruoppolo, V.; Mendoza-Sassi, R.A.; Adornes, A.C.; Cabana, Â.L.; Meireles, M.C.A. Incidence density, proportionate mortality, and risk factors of aspergillosis in Magellanic penguins in a rehabilitation center from Brazil. J. Zoo Wildl. Med. 2015, 46, 667–674. [Google Scholar]
- Ashley, E.S.D.; Lewis, R.; Lewis, J.S.; Martin, C.; Andes, D. Pharmacology of systemic antifungal agents. Clin. Infect. Dis. 2006, 43, S28–S39. [Google Scholar]
- Miles, M.R.; Levy, C.; Morel, W.; Mueller, T.; Steinlage, T.; Van Rij, N.; Frederick, R.D.; Hartman, G.L. International fungicide efficacy trials for the management of soybean rust. Plant Dis. 2007, 91, 1450–1458. [Google Scholar]
- Uppala, S.; Zhou, X.G. Field efficacy of fungicides for management of sheath blight and narrow brown leaf spot of rice. Crop Prot. 2018, 104, 72–77. [Google Scholar]
- Fang, W.; Latgé, J.P. Microbe profile: Aspergillus fumigatus: A saprotrophic and opportunistic fungal pathogen. Microbiology 2018, 164, 1009–1011. [Google Scholar]
- Bennett, J. An overview of the genus Aspergillus. In Aspergillus: Molecular Biology and Genomics; Machida, M., Gomi, K., Eds.; Caister Academic Press: Portland, Oregon, 2010; p. 238. ISBN 9781904455530. [Google Scholar]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar]
- Kocsubé, S.; Perrone, G.; Magistà, D.; Houbraken, J.; Varga, J.; Szigeti, G.; Hubka, V.; Hong, S.B.; Frisvad, J.C.; Samson, R.A. Aspergillus is monophyletic: Evidence from multiple gene phylogenies and extrolites profiles. Stud. Mycol. 2016, 85, 199–213. [Google Scholar]
- Hubka, V.; Dudová, Z.; Kubátová, A.; Frisvad, J.C.; Yaguchi, T.; Horie, Y.; Jurjević, Ž.; Hong, S.B.; Kolařík, M. Taxonomic novelties in Aspergillus section Fumigati: A. tasmanicus sp. nov., induction of sexual state in A. turcosus and overview of related species. Plant Syst. Evol. 2017, 303, 787–806. [Google Scholar]
- Hubka, V.; Barrs, V.; Dudová, Z.; Sklenář, F.; Kubátová, A.; Matsuzawa, T.; Yaguchi, T.; Horie, Y.; Nováková, A.; Frisvad, J.C.; et al. Unravelling species boundaries in the Aspergillus viridinutans complex (Section Fumigati): Opportunistic human and animal pathogens capable of interspecific hybridization. Pers. Mol. Phylogeny Evol. Fungi 2018, 41, 142–174. [Google Scholar]
- Lamoth, F. Aspergillus fumigatus-related species in clinical practice. Front. Microbiol. 2016, 7, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisvad, J.C.; Larsen, T.O. Extrolites of Aspergillus fumigatus and other pathogenic species in Aspergillus section Fumigati. Front. Microbiol. 2016, 6, 1485. [Google Scholar] [CrossRef] [PubMed]
- Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Cuenca-Estrella, M. Antifungal susceptibility profile of cryptic species of Aspergillus. Mycopathologia 2014, 178, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.J.; Barrs, V.R. One-health pathogens in the Aspergillus viridinutans complex. Med. Mycol. 2018, 56. [Google Scholar] [CrossRef]
- Rhodes, J.C. Aspergillus fumigatus: Growth and virulence. Med. Mycol. 2006, 44, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhabhra, R.; Askew, D.S. Thermotolerance and virulence of Aspergillus fumigatus: Role of the fungal nucleolus. Med. Mycol. 2005, 43, 87–93. [Google Scholar] [CrossRef]
- Coleman, J.A.; Jones, A.M.; Collier, L.J.; Richardson, M.D.; Bright-Thomas, R.J. M1 Do climate changes influence environmental Aspergillus fumigatus load at the manchester university NHS foundation trust adult cystic fibrosis centre? Thorax 2019, 74, A235. [Google Scholar]
- GAFFI Global Action Fund For Fungal Infections. Available online: Gaffi.org (accessed on 26 September 2020).
- Pfaller, M.; Wenzel, R. Impact of the changing epidemiology of fungal infections in the 1990s. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 287–291. [Google Scholar] [CrossRef]
- Bitar, D.; Lortholary, O.; Le Strat, Y.; Nicolau, J.; Coignard, B.; Tattevin, P.; Che, D.; Dromer, F. Population_based analysis of invasive fungal infections, France, 2001–2010. Emerg. Infect. Dis. 2014, 20, 1149–1155. [Google Scholar] [CrossRef]
- Maertens, J.; Vrebos, M.; Boogaerts, M. Assessing risk factors for systemic fungal infections. Eur. J. Cancer Care 2001, 10, 56–62. [Google Scholar] [CrossRef]
- Zilberberg, M.D.; Nathanson, B.H.; Harrington, R.; Spalding, J.R.; Shorr, A.F. Epidemiology and outcomes of hospitalizations with invasive aspergillosis in the United States, 2009–2013. Clin. Infect. Dis. 2018, 67, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Marr, K.A.; Carter, R.A.; Boeckh, M.; Martin, P.; Corey, L. Invasive aspergillosis in allogeneic stem cell transplant recipients: Changes in epidemiology and risk factors. Blood 2002, 100, 4358–4366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marr, K.A.; Carter, R.A.; Crippa, F.; Wald, A.; Corey, L. Epidemiology and outcome of mould infections in hematopoietic stem cell transplant recipients. Clin. Infect. Dis. 2002, 34, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, J.L.; Ruoss, S.J.; Bower, N.D.; Lin, M.; Holodniy, M.; Stevens, D.A. Diagnosing invasive fungal disease in critically ill patients. Crit. Rev. Microbiol. 2011, 37, 277–312. [Google Scholar] [CrossRef]
- Crum-Cianflone, N.F. Invasive aspergillosis associated with severe influenza infections. Open Forum Infect. Dis. 2016, 3, 1–8. [Google Scholar] [CrossRef]
- Shah, M.M.; Hsiao, E.I.; Kirsch, C.M.; Gohil, A.; Narasimhan, S.; Stevens, D.A. Invasive pulmonary aspergillosis and influenza co-infection in immunocompetent hosts: Case reports and review of the literature. Diagn. Microbiol. Infect. Dis. 2018, 91, 147–152. [Google Scholar] [CrossRef]
- Koehler, P.; Cornely, O.A.; Bottiger, B.W.; Dusse, F.; Eichenauer, D.A.; Fuchs, F.; Hallek, M.; Jung, N.; Klein, F.; Persigehl, T.; et al. COVID-19 associated pulmonary aspergillosis. Mycoses 2020, 63, 528–534. [Google Scholar] [CrossRef]
- Blaize, M.; Mayaux, J.; Nabet, C.; Lampros, A.; Marcelin, A.-G.; Thellier, M.; Piarroux, R.; Demoule, A.; Fekkar, A. Fatal invasive aspergillosis and coronavirus disease in an immunocompetent patient. Emerg. Infect. Dis. J. 2020, 26, 1636. [Google Scholar] [CrossRef]
- Gangneux, J.P.; Reizine, F.; Guegan, H.; Pinceaux, K.; Le Balch, P.; Prat, E.; Pelletier, R.; Belaz, S.; Le Souhaitier, M.; Le Tulzo, Y.; et al. Is the covid-19 pandemic a good time to include Aspergillus molecular detection to categorize aspergillosis in ICU patients? A monocentric experience. J. Fungi 2020, 6, 105. [Google Scholar] [CrossRef]
- Bartoletti, M.; Pascale, R.; Cricca, M.; Rinaldi, M.; Maccaro, A.; Bussini, L.; Fornaro, G.; Tonetti, T.; Pizzilli, G.; Francalanci, E.; et al. Epidemiology of invasive pulmonary aspergillosis among COVID-19 intubated patients: A prospective study. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Barrs, V.R.; van Doorn, T.M.; Houbraken, J.; Kidd, S.E.; Martin, P.; Pinheiro, M.D.; Richardson, M.; Varga, J.; Samson, R.A. Aspergillus felis sp. nov., an emerging agent of invasive aspergillosis in humans, cats, and dogs. PLoS ONE 2013, 8, e64871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribano, P.; Pelez, T.; Munoz, P.; Bouza, E.; Guinea, J. Is azole resistance in Aspergillus fumigatus a problem in Spain? Antimicrob. Agents Chemother. 2013, 57, 2815–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, S.J. Multi-resistant aspergillosis due to cryptic species. Mycopathologia 2014, 178, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Negri, C.E.; Gonçalves, S.S.; Xafranski, H.; Bergamasco, M.D.; Aquino, V.R.; Castro, P.T.O.; Colombo, A.L. Cryptic and rare Aspergillus species in Brazil: Prevalence in clinical samples and in vitro susceptibility to triazoles. J. Clin. Microbiol. 2014, 52, 3633–3640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Pauw, B.; Walsh, T.J.; Donnelly, J.P.; Stevens, D.A.; Edwards, J.E.; Calandra, T.; Pappas, P.G.; Maertens, J.; Lortholary, O.; Kauffman, C.; et al. Revised definitions of invasive fungal disease from the European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group and the National Institute of Allergy and Infectious Diseases Mycoses Study Group (EORTC/MSG) C. Clin. Infect. Dis. 2008, 46, 1813–1821. [Google Scholar] [CrossRef]
- Denning, D.W.; Page, I.D.; Chakaya, J.; Jabeen, K.; Jude, C.M.; Cornet, M.; Alastruey-Izquierdo, A.; Bongomin, F.; Bowyer, P.; Chakrabarti, A.; et al. Case definition of chronic pulmonary aspergillosis in resource-constrained settings. Emerg. Infect. Dis. 2018, 24, e1–e13. [Google Scholar] [CrossRef]
- Verweij, P.E.; Rijnders, B.J.A.; Brüggemann, R.J.M.; Azoulay, E.; Bassetti, M.; Blot, S.; Calandra, T.; Clancy, C.J.; Cornely, O.A.; Chiller, T.; et al. Review of influenza-associated pulmonary aspergillosis in ICU patients and proposal for a case definition: An expert opinion. Intensive Care Med. 2020, 46, 1524–1535. [Google Scholar] [CrossRef]
- White, P.L.; Dhillon, R.; Cordey, A.; Hughes, H.; Faggian, F.; Soni, S.; Pandey, M.; Whitaker, H.; May, A.; Morgan, M.; et al. A National Strategy to Diagnose Coronavirus Disease 2019–Associated Invasive Fungal Disease in the Intensive Care Unit. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Arné, P.; Thierry, S.; Wang, D.; Deville, M.; Le Loc’h, G.; Desoutter, A.; Féménia, F.; Nieguitsila, A.; Huang, W.; Chermette, R.; et al. Aspergillus fumigatus in poultry. Int. J. Microbiol. 2011, 2011. [Google Scholar] [CrossRef] [Green Version]
- Tell, L.A. Aspergillosis in mammals and birds: Impact on veterinary medicine. Med. Mycol. 2005, 43, 71–73. [Google Scholar] [CrossRef]
- Beernaert, L.A.; Pasmans, F.; Van Waeyenberghe, L.; Haesebrouck, F.; Martel, A. Aspergillus infections in birds: A review. Avian Pathol. 2010, 39, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, M.O.; Soares, M.P.; Meinerz, A.R.M.; Nobre, M.O.; Osório, L.G.; Da Silva Filho, R.P.; Meireles, M.C.A. Aspergillosis: A limiting factor during recovery of captive Magellanic penguins. Braz. J. Microbiol. 2007, 38, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.M.; da Silva Filho, R.P.; Poester, V.R.; Fernandes, C.G.; von Groll, A.; Stevens, D.A.; Sabino, R.; Xavier, M.O. Aspergillosis in albatrosses. Med. Mycol. 2019, 58, 852–855. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.J.; Thompson, P.; Vogelnest, L.; Barrs, V.R. Identification of pathogenic Aspergillus isolates from captive birds in Australia. Med. Mycol. 2018, 56, 1038–1041. [Google Scholar] [CrossRef]
- Sabino, R.; Burco, J.; Valente, J.; Veríssimo, C.; Clemons, K.V.; Stevens, D.A.; Tell, L.A. Molecular identification of clinical and environmental avian Aspergillus isolates. Arch. Microbiol. 2019, 201, 253–257. [Google Scholar] [CrossRef]
- Hof, H.; Kupfahl, C. Gliotoxin in Aspergillus fumigatus: An example that mycotoxins are potential virulence factors. Mycotoxin Res. 2009, 25, 123–131. [Google Scholar] [CrossRef]
- França, M.; Cray, C.; Shivaprasad, H.L. Serologic testing for aspergillosis in commercial broiler chickens and turkeys. Avian Dis. 2012, 56, 160–164. [Google Scholar] [CrossRef]
- Burco, J.D.; Ziccardi, M.H.; Clemons, K.V.; Tell, L.A. Evaluation of plasma (1→3) β-D-glucan concentrations in birds naturally and experimentally infected with Aspergillus fumigatus. Avian Dis. 2012, 56, 183–191. [Google Scholar] [CrossRef]
- Cabana, Â.L.; Xavier, M.O.; Poester, V.; Klafke, G.B.; Filho, P.L.B.; Martins, A.; Filho, R.P.S.; Meireles, M.C.A. Serological monitoring of antibodies for an early diagnosis of aspergillosis in captive penguins. Pesqui. Vet. Bras. 2015, 35, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Cabana, A.L.; Xavier, M.O.; Mendes, J.F.; Teles, A.J.; Martins, A.M.; Silva-Filho, R.P.; Meireles, M.C.A. Applicability of the Platelia EIA® Aspergillus test for the diagnosis of aspergilosis in penguins. Braz. J. Biol. 2018, 79, 169–173. [Google Scholar] [CrossRef]
- Fischer, D.; Van Waeyenberghe, L.; Cray, C.; Gross, M.; Usleber, E.; Pasmans, F.; Martel, A.; Lierz, M. Comparison of diagnostic tools for the detection of aspergillosis in blood samples of experimentally infected falcons. Avian Dis. 2014, 58, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Cray, C.; Watson, T.; Arheart, K.L. Serosurvey and diagnostic application of antibody titers to Aspergillus in avian species. Avian Dis. 2009, 53, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Desoubeaux, G.; Chauvin, D.; Del Carmen Piqueras, M.; Bronson, E.; Bhattacharya, S.K.; Sirpenski, G.; Bailly, E.; Cray, C. Translational proteomic study to address host protein changes during aspergillosis. PLoS ONE 2018, 13, e0200843. [Google Scholar] [CrossRef] [PubMed]
- Hauck, R.; Cray, C.; França, M. Spotlight on avian pathology: Aspergillosis. Avian Pathol. 2020, 49, 115–118. [Google Scholar] [CrossRef]
- Echenique, J.V.Z.; Soares, M.P.; Albano, A.P.N.; Bandarra, P.M.; Schild, A.L. Diseases of wild birds in southern Rio Grande do Sul, Brazil. Pesqui. Vet. Bras. 2020, 40, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Khosravi, A.R.; Shokri, H.; Ziglari, T.; Naeini, A.R.; Mousavi, Z.; Hashemi, H. Outbreak of severe disseminated aspergillosis in a flock of ostrich (Struthio camelus). Mycoses 2008, 51, 557–559. [Google Scholar] [CrossRef]
- Lupo, C.; Le Bouquin, S.; Allain, V.; Balaine, L.; Michel, V.; Petetin, I.; Colin, P.; Chauvin, C. Risk and indicators of condemnation of male turkey broilers in western France, February-July 2006. Prev. Vet. Med. 2010, 94, 240–250. [Google Scholar] [CrossRef]
- Nawrot, U.; Wieliczko, A.; Włodarczyk, K.; Kurzyk, E.; Brillowska-Dąbrowska, A. Low frequency of itraconazole resistance found among Aspergillus fumigatus originating from poultry farms in Southwest Poland. J. Mycol. Med. 2019, 29, 24–27. [Google Scholar] [CrossRef]
- Zafra, R.; Pérez, J.; Pérez-Ecija, R.A.; Borge, C.; Bustamante, R.; Carbonero, A.; Tarradas, C. Concurrent aspergillosis and ascites with high mortality in a farm of growing broiler chickens. Avian Dis. 2008, 52, 711–713. [Google Scholar] [CrossRef]
- Sajid, M.A.; Khan, I.A.; Rauf, U. Aspergillus fumigatus in commercial poultry flocks, a serious threat to poultry industry in Pakistan. J. Anim. Pl. Sci. 2006, 16, 79–81. [Google Scholar]
- Burco, J.D.; Etienne, K.A.; Massey, J.G.; Ziccardi, M.H.; Balajee, S.A. Molecular sub-typing suggests that the environment of rehabilitation centers may be a potential source of Aspergillus fumigatus infecting rehabilitating seabirds. Med. Mycol. 2012, 50, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chege, S.; Howlett, J.; Al Qassimi, M.; Toosy, A.; Kinne, J.; Obanda, V. Opportunistic infection of Aspergillus and bacteria in captive Cape vultures (Gyps coprotheres). Asian Pac. J. Trop. Biomed. 2013, 3, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, G.C.; Rewell, R.E. The incidence of aspergillosis in captive wild birds. J. Comp. Pathol. Ther. 1949, 59, 213-IN17. [Google Scholar] [CrossRef]
- Wolff, P.L.; Petrini, K.R.; Kolmstetter, C. An outbreak of aspergillosis in crested wood partridges ( Rollulus rouloul ). J. Zoo Wildl. Med. 1992, 23, 108–112. [Google Scholar]
- Dykstra, M.J.; Loomis, M.; Reininger, K.; Zombeck, D.; Faucette, T. A comparison of sampling methods for airborne fungal spores during an outbreak of aspergillosis in the forest aviary of the North Carolina Zoological Park. J. Zoo Wildl. Med. 1997, 28, 454–463. [Google Scholar] [PubMed]
- Melo, A.M.; da Silva-Filho, R.P.; Poester, V.R.; von Groll, A.; Fernandes, C.G.; Stevens, D.A.; Sabino, R.; Xavier, M.O. Aspergillosis in free-ranging aquatic birds. Med. Mycol. Case Rep. 2020, 28, 36–38. [Google Scholar] [CrossRef]
- Nardoni, S.; Ceccherelli, R.; Rossi, G.; Mancianti, F. Aspergillosis in Larus cachinnans micaellis: Survey of eight cases. Mycopathologia 2006, 161, 317–321. [Google Scholar] [CrossRef]
- Olias, P.; Gruber, A.D.; Winfried, B.; Hafez, H.M.; Lierz, M. Fungal pneumonia as a major cause of mortality in white stork (Ciconia ciconia) chicks. Avian Dis. Dig. 2010, 5, e36–e37. [Google Scholar]
- Di Somma, A.; Bailey, T.; Silvanose, C.; Garcia-Martinez, C. The use of voriconazole for the treatment of aspergillosis in falcons (Falco species). J. Avian Med. Surg. 2007, 21, 307–316. [Google Scholar] [CrossRef]
- Wang, D.Y.; Gricourt, M.; Arné, P.; Thierry, S.; Seguin, D.; Chermette, R.; Huang, W.Y.; Dannaoui, E.; Botterel, F.; Guillot, J. Mutations in the Cyp51A gene and susceptibility to itraconazole in Aspergillus fumigatus isolated from avian farms in France and China. Poult. Sci. 2014, 93, 12–15. [Google Scholar] [CrossRef]
- Fate, M.A.; Skeeles, J.K.; Beasley, J.N.; Slavik, M.F.; Lapp, N.A.; Shriver, J.W. Efficacy of thiabendazole (Mertect 340-F) in controlling mold in turkey confinement housing. Avian Dis. 1987, 31, 145–148. [Google Scholar] [CrossRef]
- Redmann, T.; Schildger, B. Therapeutic use of enilconazole in broiler chicks with aspergillosis. Dtsch. Tierarztl. Wochenschr. 1989, 96, 15–17. [Google Scholar] [PubMed]
- Williams, C.J.; Brake, J. Evaluation of application methods for control of Aspergillus fumigatus proliferation on the air cell membrane of in ovo injected broiler eggs. Poult. Sci. 2000, 79, 1531–1535. [Google Scholar] [CrossRef] [PubMed]
- Sabino, R.; Carolino, E.; Veríssimo, C.; Martinez, M.; Clemons, K.V.; Stevens, D.A. Antifungal susceptibility of 175 Aspergillus isolates from various clinical and environmental sources. Med. Mycol. 2016, 54, 740–756. [Google Scholar] [CrossRef] [Green Version]
- Howard, S.J.; Arendrup, M.C. Acquired antifungal drug resistance in Aspergillus fumigatus: Epidemiology and detection. Med. Mycol. 2011, 49, 90–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlin, D.S.; Shor, E.; Zhao, Y. Update on antifungal drug resistance. Curr. Clin. Microbiol. Rep. 2015, 2, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W.; Oakley, K.L.; Kelly, S.L.; Stevens, D.A.; Warnock, D.W. Itraconazole resistance in Aspergillus fumigatus. Antimicrob. Agents Chemother. 1997, 41, 1364–1368. [Google Scholar] [CrossRef] [Green Version]
- Mellado, E.; Alcazar-Fuoli, L.; García-Effrón, G.; Alastruey-Izquierdo, A.; Cuenca-Estrella, M.; Rodríguez-Tudela, J.L. New resistance mechanisms to azole drugs in Aspergillus fumigatus and emergence of antifungal drugs-resistant A. fumigatus atypical strains. Med. Mycol. 2006, 44, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, A.R.; Lagrou, K.; Meis, J.F.; Chowdhary, A.; Lockhart, S.R.; Verweij, P.E. Triazole resistance surveillance in Aspergillus fumigatus. Med. Mycol. 2018, 56, S83–S92. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Moreno, C.; Lavergne, R.A.; Hagen, F.; Morio, F.; Meis, J.F.; Le Pape, P. Azole-resistant Aspergillus fumigatus harboring TR 34/L98H, TR 46/Y121F/T289A and TR 53 mutations related to flower fields in Colombia. Sci. Rep. 2017, 7, 45631. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Denardi, L.B.; Melchers, W.J.G.; Zoll, J.; Buil, J.B.; Hagen, F.; Meis, J.F.; Alves, S.H.; Verweij, P.E. First report of azole-resistent Aspergillus fumigatus harboring TR34/L98H and M220R in Brazil. In Proceedings of the 20th Congress of the International Society for Human and Animal Mycology, Amsterdam, The Netherlands, 30 June–4 July 2018. [Google Scholar]
- Prigitano, A.; Esposto, M.C.; Romanò, L.; Auxilia, F.; Tortorano, A.M. Azole-resistant Aspergillus fumigatus in the Italian environment. J. Glob. Antimicrob. Resist. 2019, 16, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Feng, C.L.; Chen, F.; He, Q.; Su, X.; Shi, Y. Triazole resistance in Aspergillus fumigatus clinical isolates obtained in Nanjing, China. Chin. Med. J. 2017, 130, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Abdolrasouli, A.; Petrou, M.A.; Park, H.; Rhodes, J.L.; Rawson, T.M.; Moore, L.S.P.; Donaldson, H.; Holmes, A.H.; Fisher, M.C.; Armstrong-James, D. Surveillance for azole-resistant Aspergillus fumigatus in a centralized diagnostic mycology service, London, United Kingdom, 1998–2017. Front. Microbiol. 2018, 9, 2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, R.H.; Hagen, F.; Astvad, K.M.T.; Tyron, A.; Meis, J.F.; Arendrup, M.C. Azole-resistant Aspergillus fumigatus in Denmark: A laboratory-based study on resistance mechanisms and genotypes. Clin. Microbiol. Infect. 2016, 22, 570.e1–570.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelièvre, L.; Groh, M.; Angebault, C.; Maherault, A.C.; Didier, E.; Bougnoux, M.E. Azole resistant Aspergillus fumigatus: An emerging problem. Med. Mal. Infect. 2013, 43, 139–145. [Google Scholar] [CrossRef]
- Talbot, J.J.; Subedi, S.; Halliday, C.L.; Hibbs, D.E.; Lai, F.; Lopez-Ruiz, F.J.; Harper, L.; Park, R.F.; Cuddy, W.S.; Biswas, C.; et al. Surveillance for azole resistance in clinical and environmental isolates of Aspergillus fumigatus in Australia and cyp51A homology modelling of azole-resistant isolates. J. Antimicrob. Chemother. 2018, 73, 2347–2351. [Google Scholar] [CrossRef] [Green Version]
- Prigitano, A.; Venier, V.; Cogliati, M.; De Lorenzis, G.; Esposto, M.C.; Tortorano, A.M. Azole-resistant Aspergillus fumigatus in the environment of Northern Italy, May 2011 to June 2012. Eurosurveillance 2014, 19, 20747. [Google Scholar] [CrossRef] [Green Version]
- Van der Linden, J.W.M.; Arendrup, M.C.; Warris, A.; Lagrou, K.; Pelloux, H.; Hauser, P.M.; Chryssanthou, E.; Mellado, E.; Kidd, S.E.; Tortorano, A.M.; et al. Prospective multicenter international surveillance of azole resistance in Aspergillus fumigatus. Emerg. Infect. Dis. 2015, 21, 1041–1044. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; Cuenca-Estrella, M.; Mellado, E. Triazole resistance in Aspergillus species: An emerging problem. Drugs 2017, 77, 599–613. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-resistant aspergillosis: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S436–S444. [Google Scholar] [CrossRef] [Green Version]
- Sabino, R.; Veríssimo, C.; Parada, H.; Brandão, J.; Viegas, C.; Carolino, E.; Clemons, K.V.; Stevens, D.A. Molecular screening of 246 Portuguese Aspergillus isolates among different clinical and environmental sources. Med. Mycol. 2014, 52, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhren, J.; Voskuil, W.S.; Boel, C.H.E.; Haas, P.J.A.; Hagen, F.; Meis, J.F.; Kusters, J.G. High prevalence of azole resistance in Aspergillus fumigatus isolates from high-risk patients. J. Antimicrob. Chemother. 2015, 70, 2894–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashu, E.E.; Hagen, F.; Chowdhary, A.; Meis, J.F.; Xu, J. Global population genetic analysis of Aspergillus. mSphere 2017, 2, e00019-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziółkowska, G.; Tokarzewski, S.; Nowakiewicz, A. Drug resistance of Aspergillus fumigatus strains isolated from flocks of domestic geese in Poland. Poult. Sci. 2014, 93, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Spanamberg, A.; Ravazzolo, A.P.; Denardi, L.B.; Hartz, S.A.; Santurio, J.M.; Driemeier, D.; Ferreiro, L. Antifungal susceptibility profile of Aspergillus fumigatus isolates from avian lungs. Pesqui. Veterinária Bras. 2020, 40, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Barber, A.E.; Scheufen, S.; Walther, G.; Kurzai, O.; Schmidt, V. Low rate of azole resistance in cases of avian aspergillosis in Germany. Med. Mycol. 2020. [Google Scholar] [CrossRef]
- Beernaert, L.A.; Pasmans, F.; Van Waeyenberghe, L.; Dorrestein, G.M.; Verstappen, F.; Vercammen, F.; Haesebrouck, F.; Martel, A. Avian Aspergillus fumigatus strains resistant to both itraconazole and voriconazole. Antimicrob. Agents Chemother. 2009, 53, 2199–2201. [Google Scholar] [CrossRef] [Green Version]
- Buil, J.B.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular detection of azole-resistant Aspergillus fumigatus in clinical samples. Front. Microbiol. 2018, 9, 515. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Sharma, C.; Hagen, F.; Meis, J.F. Exploring azole antifungal drug resistance in Aspergillus fumigatus with special reference to resistance mechanisms. Future Microbiol. 2014, 9, 697–711. [Google Scholar] [CrossRef]
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Rijs, A.J.M.M.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Genotype-phenotype complexity of the TR46/Y121F/T289A cyp51A azole resistance mechanism in Aspergillus fumigatus. Fungal Genet. Biol. 2015, 82, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Snelders, E.; Karawajczyk, A.; Schaftenaar, G.; Verweij, P.E.; Melchers, W.J.G. Azole resistance profile of amino acid changes in Aspergillus fumigatus CYP51A based on protein homology modeling. Antimicrob. Agents Chemother. 2010, 54, 2425–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snelders, E.; Karawajczyk, A.; Verhoeven, R.J.A.; Venselaar, H.; Schaftenaar, G.; Verweij, P.E.; Melchers, W.J.G. The structure-function relationship of the Aspergillus fumigatus cyp51A L98H conversion by site-directed mutagenesis: The mechanism of L98H azole resistance. Fungal Genet. Biol. 2011, 48, 1062–1070. [Google Scholar] [CrossRef]
- Ren, J.; Jin, X.; Zhang, Q.; Zheng, Y.; Lin, D.; Yu, Y. Fungicides induced triazole-resistance in Aspergillus fumigatus associated with mutations of TR46/Y121F/T289A and its appearance in agricultural fields. J. Hazard. Mater. 2017, 326, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Van Der Linden, J.W.M.; Camps, S.M.T.; Kampinga, G.A.; Arends, J.P.A.; Debets-Ossenkopp, Y.J.; Haas, P.J.A.; Rijnders, B.J.A.; Kuijper, E.J.; Van Tiel, F.H.; Varga, J.; et al. Aspergillosis due to voriconazole highly resistant Aspergillus fumigatus and recovery of genetically related resistant isolates from domiciles. Clin. Infect. Dis. 2013, 57, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsitsopoulou, A.; Posso, R.; Vale, L.; Bebb, S.; Johnson, E.; White, P.L. Determination of the prevalence of triazole resistance in environmental Aspergillus fumigatus strains isolated in South Wales, UK. Front. Microbiol. 2018, 9, 1395. [Google Scholar] [CrossRef] [PubMed]
- Morio, F.; Aubin, G.G.; Danner-Boucher, I.; Haloun, A.; Sacchetto, E.; Garcia-Hermoso, D.; Bretagne, S.; Miegeville, M.; Le Pape, P. High prevalence of triazole resistance in Aspergillus fumigatus, especially mediated by TR/L98H, in a French cohort of patients with cystic fibrosis. J. Antimicrob. Chemother. 2012, 67, 1870–1873. [Google Scholar] [CrossRef] [Green Version]
- Dauchy, C.; Bautin, N.; Nseir, S.; Reboux, G.; Wintjens, R.; Le Rouzic, O.; Sendid, B.; Viscogliosi, E.; Le Pape, P.; Arendrup, M.C.; et al. Emergence of Aspergillus fumigatus azole resistance in azole-naïve patients with chronic obstructive pulmonary disease and their homes. Indoor Air 2018, 28, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Bowyer, P.; Moore, C.B.; Rautemaa, R.; Denning, D.W.; Richardson, M.D. Azole antifungal resistance today: Focus on Aspergillus. Curr. Infect. Dis. Rep. 2011, 13, 485–491. [Google Scholar] [CrossRef]
- Camps, S.M.T.; Dutilh, B.E.; Arendrup, M.C.; Rijs, A.J.M.M.; Snelders, E.; Huynen, M.A.; Verweij, P.E.; Melchers, W.J.G. Discovery of a hapE mutation that causes azole resistance in Aspergillus fumigatus through whole genome sequencing and sexual crossing. PLoS ONE 2012, 7, e50034. [Google Scholar] [CrossRef] [Green Version]
- Hortschansky, P.; Misslinger, M.; Mörl, J.; Gsaller, F.; Bromley, M.J.; Brakhage, A.A.; Groll, M.; Haas, H.; Huber, E.M. Structural basis of HapEP88L-linked antifungal triazole resistance in Aspergillus fumigatus. Life Sci. Alliance 2020, 3. [Google Scholar] [CrossRef] [PubMed]
- Wel, X.; Chen, P.; Gao, R.; Li, Y.; Zhang, A.; Liu, F.; Lu, L. Screening and characterization of a non-cyp51A mutation in an Aspergillus fumigatus cox10 strain conferring azole resistance. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Rybak, J.M.; Ge, W.; Wiederhold, N.P.; Parker, J.E.; Kelly, S.L.; Rogers, P.D.; Fortwendel, J.R. Mutations in hmg1, challenging the paradigm of clinical triazole resistance in Aspergillus fumigatus. MBio 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, C.; Nelson-Sathi, S.; Singh, A.; Radhakrishna Pillai, M.; Chowdhary, A. Genomic perspective of triazole resistance in clinical and environmental Aspergillus fumigatus isolates without cyp51A mutations. Fungal Genet. Biol. 2019, 132, 103265. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.G.; Bromley, M.; Buied, A.; Moore, C.B.; Rajendran, R.; Rautemaa, R.; Ramage, G.; Denning, D.W.; Bowyer, P. The cdr1B efflux transporter is associated with non-cyp51a-mediated itraconazole resistance in Aspergillus fumigatus. J. Antimicrob. Chemother. 2013, 68, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Diekema, D.; Moye-Rowley, W.S. Contributions of Aspergillus fumigatus ATP-binding cassette transporter proteins to drug resistance and virulence. Eukaryot. Cell 2013, 12, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slaven, J.W.; Anderson, M.J.; Sanglard, D.; Dixon, G.K.; Bille, J.; Roberts, I.S.; Denning, D.W. Increased expression of a novel Aspergillus fumigatus ABC transporter gene, atrF, in the presence of itraconazole in an itraconazole resistant clinical isolate. Fungal Genet. Biol. 2002, 36, 199–206. [Google Scholar] [CrossRef]
- Da Silva Ferreira, M.E.; Luiz Capellaro, J.; Dos Reis Marques, E.; Malavazi, I.; Perlin, D.; Park, S.; Anderson, J.B.; Colombo, A.L.; Arthington-Skaggs, B.A.; Goldman, M.H.S.; et al. In vitro evolution of itraconazole resistance in Aspergillus fumigatus involves multiple mechanisms of resistance. Antimicrob. Agents Chemother. 2004, 48, 4405–4413. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, A.M.; Goldman, G.H.; Park, S.; Marras, S.A.E.; Delmas, G.; Oza, U.; Lolans, K.; Dudley, M.N.; Mann, P.A.; Perlin, D.S. Multiple resistance mechanisms among Aspergillus fumigatus mutants with high-level resistance to itraconazole. Antimicrob. Agents Chemother. 2003, 47, 1719–1726. [Google Scholar] [CrossRef] [Green Version]
- Meneau, I.; Coste, A.T.; Sanglard, D. Identification of Aspergillus fumigatus multidrug transporter genes and their potential involvement in antifungal resistance. Med. Mycol. 2016, 54, 616–627. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Ferreira, M.E.; Malavazi, I.; Savoldi, M.; Brakhage, A.A.; Goldman, M.H.S.; Kim, H.S.; Nierman, W.C.; Goldman, G.H. Transcriptome analysis of Aspergillus fumigatus exposed to voriconazole. Curr. Genet. 2006, 50, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Mowat, E.; Lang, S.; Williams, C.; McCulloch, E.; Jones, B.; Ramage, G. Phase-dependent antifungal activity against Aspergillus fumigatus developing multicellular filamentous biofilms. J. Antimicrob. Chemother. 2008, 62, 1281–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Singh, S. Biofilm formation by Aspergillus fumigatus. Med. Mycol. 2015, 52, 2–9. [Google Scholar]
- Rajendran, R.; Mowat, E.; McCulloch, E.; Lappin, D.F.; Jones, B.; Lang, S.; Majithiya, J.B.; Warn, P.; Williams, C.; Ramage, G. Azole resistance of Aspergillus fumigatus biofilms is partly associated with efflux pump activity. Antimicrob. Agents Chemother. 2011, 55, 2092–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- 138. Snelders, E.; Huis In’t Veld, R.A.G.; Rijs, A.J.M.M.; Kema, G.H.J.; Melchers, W.J.G.; Verweij, P.E. Possible environmental origin of resistance of Aspergillus fumigatus to medical triazoles. Appl. Environ. Microbiol. 2009, 75, 4053–4057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snelders, E.; Camps, S.M.T.; Karawajczyk, A.; Schaftenaar, G.; Kema, G.H.J.; van der Lee, H.A.; Klaassen, C.H.; Melchers, W.J.G.; Verweij, P.E. Triazole fungicides can induce cross-resistance to medical triazoles in Aspergillus fumigatus. PLoS ONE 2012, 7, e31801. [Google Scholar] [CrossRef] [Green Version]
- Jeanvoine, A.; Rocchi, S.; Bellanger, A.P.; Reboux, G.; Millon, L. Azole-resistant Aspergillus fumigatus: A global phenomenon originating in the environment? Med. Mal. Infect. 2019, 50, 389–395. [Google Scholar] [CrossRef]
- Van der Linden, J.W.M.; Snelders, E.; Kampinga, G.A.; Rijnders, B.J.A.; Mattsson, E.; Debets-Ossenkopp, Y.J.; Kuijper, E.J.; van Tiel, F.H.; Melchers, W.J.G.; Verweij, P.E. Clinical implications of azole resistance in Aspergillus fumigatus, The Netherlands, 2007–2009. Emerg. Infect. Dis. 2011, 17, 1846–1854. [Google Scholar] [CrossRef]
- Berger, S.; El Chazli, Y.; Babu, A.F.; Coste, A.T. Azole resistance in Aspergillus fumigatus: A consequence of antifungal use in agriculture? Front. Microbiol. 2017, 8, 1024. [Google Scholar] [CrossRef] [Green Version]
- Lavergne, R.A.; Chouaki, T.; Hagen, F.; Toublanc, B.; Dupont, H.; Jounieaux, V.; Meis, J.F.; Morio, F.; Le Pape, P. Home environment as a source of life-threatening azole-resistant Aspergillus fumigatus in immunocompromised patients. Clin. Infect. Dis. 2017, 64, 76–78. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, E.; Maertens, J.; Schoemans, H.; Lagrou, K. Azole-resistant Aspergillus fumigatus due to TR46/Y121F/T289A mutation emerging in Belgium, July 2012. Eurosurveillance 2012, 17, 3–5. [Google Scholar]
- Chowdhary, A.; Sharma, C.; Kathuria, S.; Hagen, F.; Meis, J.F. Azole-resistant Aspergillus fumigatus with the environmental TR46/Y121F/T289A mutation in India. J. Antimicrob. Chemother. 2014, 69, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Sewell, T.R.; Zhu, J.; Rhodes, J.; Hagen, F.; Meis, J.F.; Fisher, M.C.; Jombart, T. Nonrandom distribution of azole resistance across the global population of Aspergillus fumigatus. MBio 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, N.; He, Y.; Yao, S.; Zhang, H.; Ren, J.; Fang, H.; Yu, Y. Tebuconazole induces triazole-resistance in Aspergillus fumigatus in liquid medium and soil. Sci. Total Environ. 2019, 648, 1237–1243. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Yao, S.; Zhang, H.; Wang, S.; Jin, X.; Lin, D.; Fang, H.; Yu, Y. Mutation in cyp51A and high expression of efflux pump gene of Aspergillus fumigatus induced by propiconazole in liquid medium and soil. Environ. Pollut. 2020, 256, 113385. [Google Scholar] [CrossRef] [PubMed]
- Pardo, E.; Marín, S.; Sanchis, V.; Ramos, A.J. Impact of relative humidity and temperature on visible fungal growth and OTA production of ochratoxigenic Aspergillus ochraceus isolates on grapes. Food Microbiol. 2005, 22, 383–389. [Google Scholar] [CrossRef]
- Nalley, L.; Tsiboe, F.; Durand-Morat, A.; Shew, A.; Thoma, G. Economic and environmental impact of rice blast pathogen (Magnaporthe oryzae) alleviation in the United States. PLoS ONE 2016, 11, e0167295. [Google Scholar] [CrossRef]
- Kandel, Y.R.; Mueller, D.S.; Hart, C.E.; Bestor, N.R.C.; Bradley, C.A.; Ames, K.A.; Giesler, L.J.; Wise, K.A. Analyses of yield and economic response from foliar fungicide and insecticide applications to soybean in the North Central United States. Plant Health Prog. 2016, 17, 232–238. [Google Scholar] [CrossRef] [Green Version]
- Rosburg, A.; Menapace, L. Factors influencing corn fungicide treatment decisions. J. Agric. Resour. Econ. 2018, 43, 151–171. [Google Scholar]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef]
- SINDVEG Ministério da Agricultura Pecuária e Abastecimento. Available online: http://www.agricultura.gov.br/assuntos/camaras-setoriais-tematicas/documentos/camaras-tematicas/insumos-agropecuarios/anos-anteriores/mercado-de-defensivos-72.pdf/view (accessed on 18 July 2019).
- Schermerhorn, P.G.; Golden, P.E.; Krynitsky, A.J.; Leimkuehler, W.M. Determination of 22 triazole compounds including parent fungicides and metabolites in apples, peaches, flour, and water by liquid chromatography/tandem mass spectrometry. J. AOAC Int. 2005, 88, 1491–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 years of research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of azole-resistant Aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, 3–7. [Google Scholar] [CrossRef]
- Wang, C.; Wu, Q.; Wu, C.; Wang, Z. Application of dispersion–solidification liquid–liquid microextraction for the determination of triazole fungicides in environmental water samples by high-performance liquid chromatography. J. Hazard. Mater. 2011, 185, 71–76. [Google Scholar] [CrossRef]
- Branchet, P.; Cadot, E.; Fenet, H.; Sebag, D.; Ngatcha, B.N.; Borrell-Estupina, V.; Ngoupayou, J.R.N.; Kengne, I.; Braun, J.J.; Gonzalez, C. Polar pesticide contamination of an urban and peri-urban tropical watershed affected by agricultural activities (Yaoundé, Center Region, Cameroon). Environ. Sci. Pollut. Res. 2018, 25, 17690–17715. [Google Scholar] [CrossRef]
- Wu, T.; Wright, K.; Hurst, S.F.; Morrison, C.J. Enhanced extracellular production of aspartyl proteinase, a virulence factor, by Candida albicans isolates following growth in subinhibitory concentrations of fluconazole. Antimicrob. Agents Chemother. 2000, 44, 1200–1208. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rodríguez, P.; Soto-Gómez, D.; Paradelo, M.; López-Periago, J.E. Concentration levels of new-generation fungicides in throughfall released by foliar wash-off from vineyards. J. Environ. Manag. 2017, 203, 467–475. [Google Scholar] [CrossRef]
- Smalling, K.L.; Reilly, T.J.; Sandstrom, M.W.; Kuivila, K.M. Occurrence and persistence of fungicides in bed sediments and suspended solids from three targeted use areas in the United States. Sci. Total Environ. 2013, 447, 179–185. [Google Scholar] [CrossRef]
- Mimbs, W.H.; Cusaac, J.P.W.; Smith, L.M.; McMurry, S.T.; Belden, J.B. Occurrence of current-use fungicides and bifenthrin in Rainwater Basin wetlands. Chemosphere 2016, 159, 275–281. [Google Scholar] [CrossRef]
- Caldas, S.S.; Arias, J.L.O.; Rombaldi, C.; Mello, L.L.; Cerqueira, M.B.R.; Martins, A.F.; Primel, E.G. Occurrence of pesticides and PPCPs in surface and drinking water in southern Brazil: Data on 4-year monitoring. J. Braz. Chem. Soc. 2019, 30, 71–80. [Google Scholar] [CrossRef]
- Parejo, M.; Gutiérrez, J.S.; Navedo, J.G.; Soriano-Redondo, A.; Abad-Gómez, J.M.; Villegas, A.; Corbacho, C.; Sánchez-Guzmán, J.M.; Masero, J.A. Day and night use of habitats by northern pintails during winter in a primary rice-growing region of Iberia. PLoS ONE 2019, 14, e0220400. [Google Scholar] [CrossRef] [PubMed]
- Somers, C.M.; Morris, R.D. Birds and wine grapes: Foraging activity causes small-scale damage patterns in single vineyards. J. Appl. Ecol. 2002, 39, 511–523. [Google Scholar] [CrossRef]
- Navedo, J.G.; Hahn, S.; Parejo, M.; Abad-Gómez, J.M.; Gutiérrez, J.S.; Villegas, A.; Sánchez-Guzmán, J.M.; Masero, J.A. Unravelling trophic subsidies of agroecosystems for biodiversity conservation: Food consumption and nutrient recycling by waterbirds in Mediterranean rice fields. Sci. Total Environ. 2015, 511, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Vallon, M.; Dietzen, C.; Laucht, S.; Ludwigs, J.D. Focal species candidates for pesticide risk assessment in European rice fields: A review. Integr. Environ. Assess. Manag. 2018, 14, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Acosta, M.; Mugica, L.; Blanco, D.; López-Lanús, B.; Dias, R.A.; Doodnath, L.W.; Hurtado, J. Birds of rice fields in the Americas. Waterbirds Int. J. Waterbird Biol. 2010, 33, 105–122. [Google Scholar] [CrossRef]
- Fujioka, M.; Lee, S.D.; Kurechi, M.; Yoshida, H. Bird Use of Rice Fields in Korea and Japan. Waterbirds Int. J. Waterbird Biol. 2010, 33, 8–29. [Google Scholar] [CrossRef]
- Sundar, K.S.G.; Subramanya, S. Bird use of rice fields in the Indian Subcontinent. Waterbirds Int. J. Waterbird Biol. 2010, 33, 44–70. [Google Scholar] [CrossRef]
- Wymenga, E.; Zwarts, L. Use of rice fields by birds in West Africa. Waterbirds 2010, 33, 97–104. [Google Scholar]
- Hannay, M.B.; Boulanger, J.R.; Curtis, P.D.; Eaton, R.A.; Hawes, B.C.; Leigh, D.K.; Rossetti, C.A.; Steensma, K.M.M.; Lindell, C.A. Bird species and abundances in fruit crops and implications for bird management. Crop Prot. 2019, 120, 43–49. [Google Scholar] [CrossRef]
- Wilcoxon, C.A.; Ward, M.P.; Walk, J.W. Bird occupancy on cover crop fields during their spring migration in the midwest corn belt region. In Proceedings of the Private Landscapes, Public Responsibilities: 77th Midwest Fish & Wildlife Conference, Lincoln, Nebraska, 5–7 February 2017. [Google Scholar]
- Kragten, S. Shift in crop preference during the breeding season by Yellow Wagtails Motacilla flava flava on arable farms in The Netherlands. J. Ornithol. 2011, 152, 751–757. [Google Scholar] [CrossRef] [Green Version]
- Rajashekara, S.; Venkatesha, M.G. Insectivorous bird communities of diverse agro-ecosystems in the Bengaluru region, India. J. Entomol. Zool. Stud. 2014, 2, 142–155. [Google Scholar]
- Dias, R.A.; Maurício, G.N.; Bugoni, L. Birds of the Patos Lagoon Estuary and adjacent coastal waters, southern Brazil: Species assemblages and conservation implications. Mar. Biol. Res. 2017, 13, 108–120. [Google Scholar] [CrossRef]
- Najdenski, H.; Dimova, T.; Zaharieva, M.M.; Nikolov, B.; Petrova-Dinkova, G.; Dalakchieva, S.; Popov, K.; Hristova-Nikolova, I.; Zehtindjiev, P.; Peev, S.; et al. Migratory birds along the mediterranean—Black sea flyway as carriers of zoonotic pathogens. Can. J. Microbiol. 2018, 64, 915–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubálek, Z. An annotated checklist of pathogenic microorganisms associated with migratory birds. J. Wildl. Dis. 2004, 40, 639–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramey, A.M.; Reeves, A.B.; Donnelly, T.; Poulson, R.L.; Stallknecht, D.E. Introduction of Eurasian-origin influenza a(H8N4) virus into north America by migratory birds. Emerg. Infect. Dis. 2018, 24, 1950–1953. [Google Scholar] [CrossRef]
- Lickfett, T.M.; Clark, E.; Gehring, T.M.; Alm, E.W. Detection of Influenza A viruses at migratory bird stopover sites in Michigan, USA. Infect. Ecol. Epidemiol. 2018, 8, 1474709. [Google Scholar] [CrossRef] [Green Version]
- Rappole, J.H.; Derrickson, S.R.; Hubalek, Z. Migratory birds and spread of west nile virus in the western hemisphere. Emerg. Infect. Dis. 2000, 6, 319–328. [Google Scholar] [CrossRef]
- Ramey, A.M.; DeLiberto, T.J.; Berhane, Y.; Swayne, D.E.; Stallknecht, D.E. Lessons learned from research and surveillance directed at highly pathogenic influenza A viruses in wild birds inhabiting North America. Virology 2018, 518, 55–63. [Google Scholar] [CrossRef]
- Al-Yasiri, M.H.; Normand, A.C.; L’Ollivier, C.; Lachaud, L.; Bourgeois, N.; Rebaudet, S.; Piarroux, R.; Mauffrey, J.F.; Ranque, S. Opportunistic fungal pathogen Candida glabrata circulates between humans and yellow-legged gulls. Sci. Rep. 2016, 6, 36157. [Google Scholar] [CrossRef] [Green Version]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the emergence of Candida auris: Climate change, azoles, swanps, and birds. MBio 2019, 10, e01397-19. [Google Scholar] [CrossRef] [Green Version]
- Young, E.A.; Cornish, T.E.; Little, S.E. Concomitant mycotic and verminous pneumonia in a blue jay from Georgia. J. Wildl. Dis. 1998, 34, 625–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, S.H.; Chmura, A.; Converse, K.; Kilpatrick, A.M.; Patel, N.; Lammers, E.; Daszak, P. Aquatic bird disease and mortality as an indicator of changing ecosystem health. Mar. Ecol. Prog. Ser. 2007, 352, 299–309. [Google Scholar] [CrossRef]
- Small, C.; Nicholls, R.J. A global analysis of human settlement in coastal zones. J. Coast. Res. 2003, 19, 584–599. [Google Scholar]
- Lerner, H.; Berg, C. The concept of health in One Health and some practical implications for research and education: What is One Health? Infect. Ecol. Epidemiol. 2015, 5, 25300. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, P.M.; Kock, R.; Kachani, M.; Kunkel, R.; Thomas, J.; Gilbert, J.; Wallace, R.; Blackmore, C.; Wong, D.; Karesh, W.; et al. Toward proof of concept of a one health approach to disease prediction and control. Emerg. Infect. Dis. 2013, 19, e130265. [Google Scholar] [CrossRef]
- Chowdhary, A.; Meis, J.F. Emergence of azole resistant Aspergillus fumigatus and One Health: Time to implement environmental stewardship. Environ. Microbiol. 2018, 20, 1299–1301. [Google Scholar] [CrossRef] [Green Version]
- Verweij, P.E.; Lucas, J.A.; Arendrup, M.C.; Bowyer, P.; Brinkmann, A.J.F.; Denning, D.W.; Dyer, P.S.; Fisher, M.C.; Geenen, P.L.; Gisi, U.; et al. The One Health problem of azole resistance in Aspergillus fumigatus: Current insights and future research agenda. Fungal Biol. Rev. 2020, 34, 202–214. [Google Scholar] [CrossRef]
- Miskiewicz, A.; Kowalczyk, P.; Oraibi, S.M.; Cybulska, K.; Misiewicz, A. Bird feathers as potential sources of pathogenic microorganisms: A new look at old diseases. Antonie van Leeuwenhoek. Int. J. Gen. Mol. Microbiol. 2018, 111, 1493–1507. [Google Scholar] [CrossRef] [Green Version]
- Garcia, N.M.; Del Negro, G.M.; Heins-Vaccari, E.M.; de Melo, N.T.; de Assis, C.M.; da Lacaz, C.S. Paracoccidioides brasiliensis, a new sample isolated from feces of a penguin (Pygoscelis adeliae). Rev. Inst. Med. Trop. Sao Paulo 1993, 35, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Meyers, J.P.; Sallaberry, A.M.; Ortiz, E.; Castro, G.; Gordon, L.J.; Maron, J.L.; Schick, C.T.; Tablio, E.; Antas, P.; Below, T. Migration routes of new world sanderlings (Calidris alba). Biol. Sci. Fac. Publ. 1990, 107, 172–180. [Google Scholar]
- Poultry Working Groups of the RUMA Alliance. Responsible Use of Antimicrobials in Poultry Production; Poultry Working Groups of the RUMA Alliance: Welwyn, UK, 2005. [Google Scholar]
- Risi, E.; Thary, V.; Arné, P.; Deville, M.; Lahitte, J.D. De Aspergillosis of seabirds in captivity. In Proceedings of the 7th International Effects of Oil and Wildlife Conference, Hamburg, Germany, 14–16 October 2003; pp. 14–16. [Google Scholar]
- Wang, D.Y. Genetic Diversity and Antifungal Susceptibility of Aspergillus spp. Isolates from Avian Farms in Guangxi, China. Ph.D. Thesis, Université de Guangxi, Guangxi, China, 2012. [Google Scholar]
- Wang, C.; Wang, J.; Su, W.; Gao, S.; Luo, J.; Zhang, M.; Xie, L.; Liu, S.; Liu, X.; Chen, Y.; et al. Relationship between domestic and wild birds in live poultry market and a novel human H7N9 virus in China. J. Infect. Dis. 2014, 209, 34–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabil, N.M.; Erfan, A.M.; Tawakol, M.M.; Haggag, N.M.; Naguib, M.M.; Samy, A. Wild birds in live birds markets: Potential reservoirs of enzootic avian influenza viruses and antimicrobial resistant enterobacteriaceae in northern Egypt. Pathogens 2020, 9, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melo, A.M.; Stevens, D.A.; Tell, L.A.; Veríssimo, C.; Sabino, R.; Xavier, M.O. Aspergillosis, Avian Species and the One Health Perspective: The Possible Importance of Birds in Azole Resistance. Microorganisms 2020, 8, 2037. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122037
Melo AM, Stevens DA, Tell LA, Veríssimo C, Sabino R, Xavier MO. Aspergillosis, Avian Species and the One Health Perspective: The Possible Importance of Birds in Azole Resistance. Microorganisms. 2020; 8(12):2037. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122037
Chicago/Turabian StyleMelo, Aryse Martins, David A. Stevens, Lisa A. Tell, Cristina Veríssimo, Raquel Sabino, and Melissa Orzechowski Xavier. 2020. "Aspergillosis, Avian Species and the One Health Perspective: The Possible Importance of Birds in Azole Resistance" Microorganisms 8, no. 12: 2037. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122037