Differential Response of Mycorrhizal Plants to Tomato bushy stunt virus and Tomato mosaic virus Infection


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Biological Materials
2.2. Viral Affirmation and Quantification
2.3. Disease Severity Evaluation
2.4. Differential Gene Expressions
2.5. Mycorrhiza and Biomass Assessment
3. Results
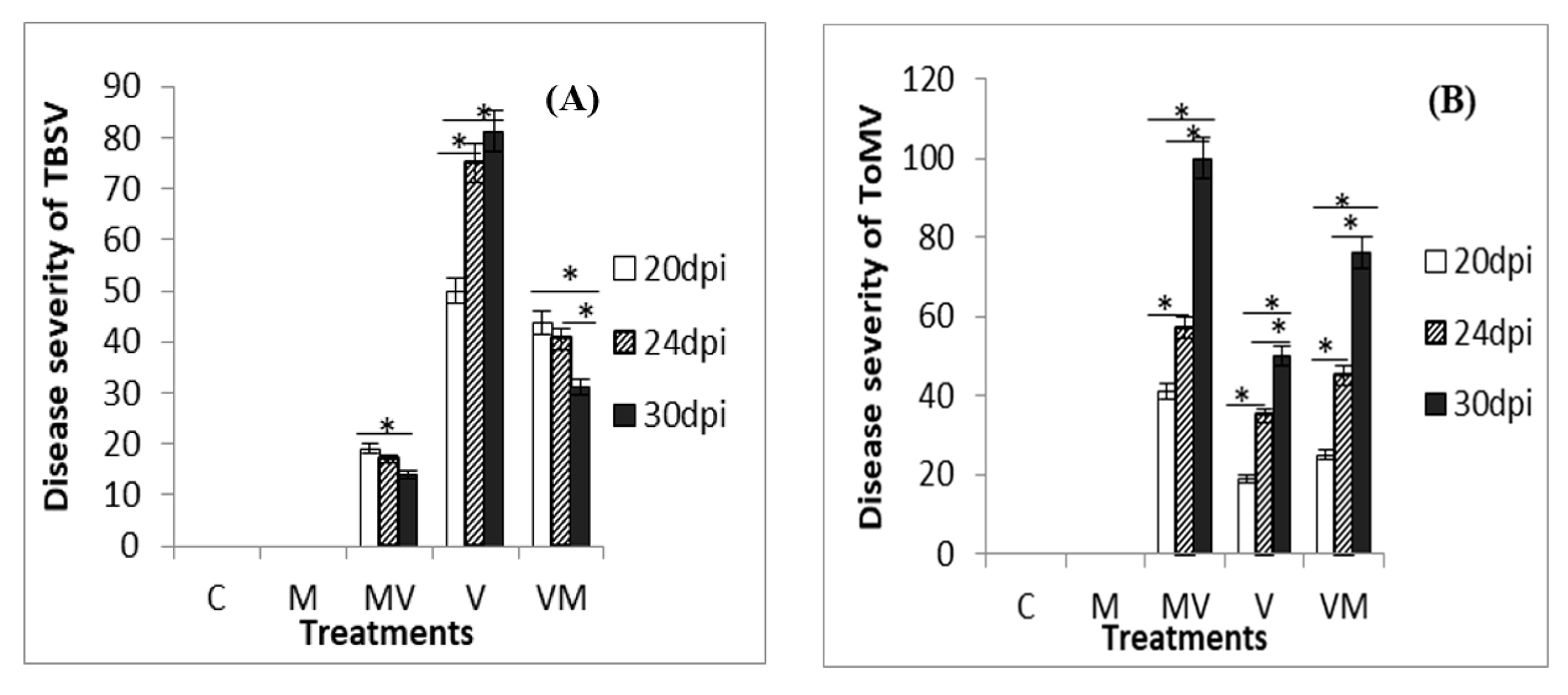
3.1. Phenotypic Responses of Mycorrhizal Plants to VIRAL Infection
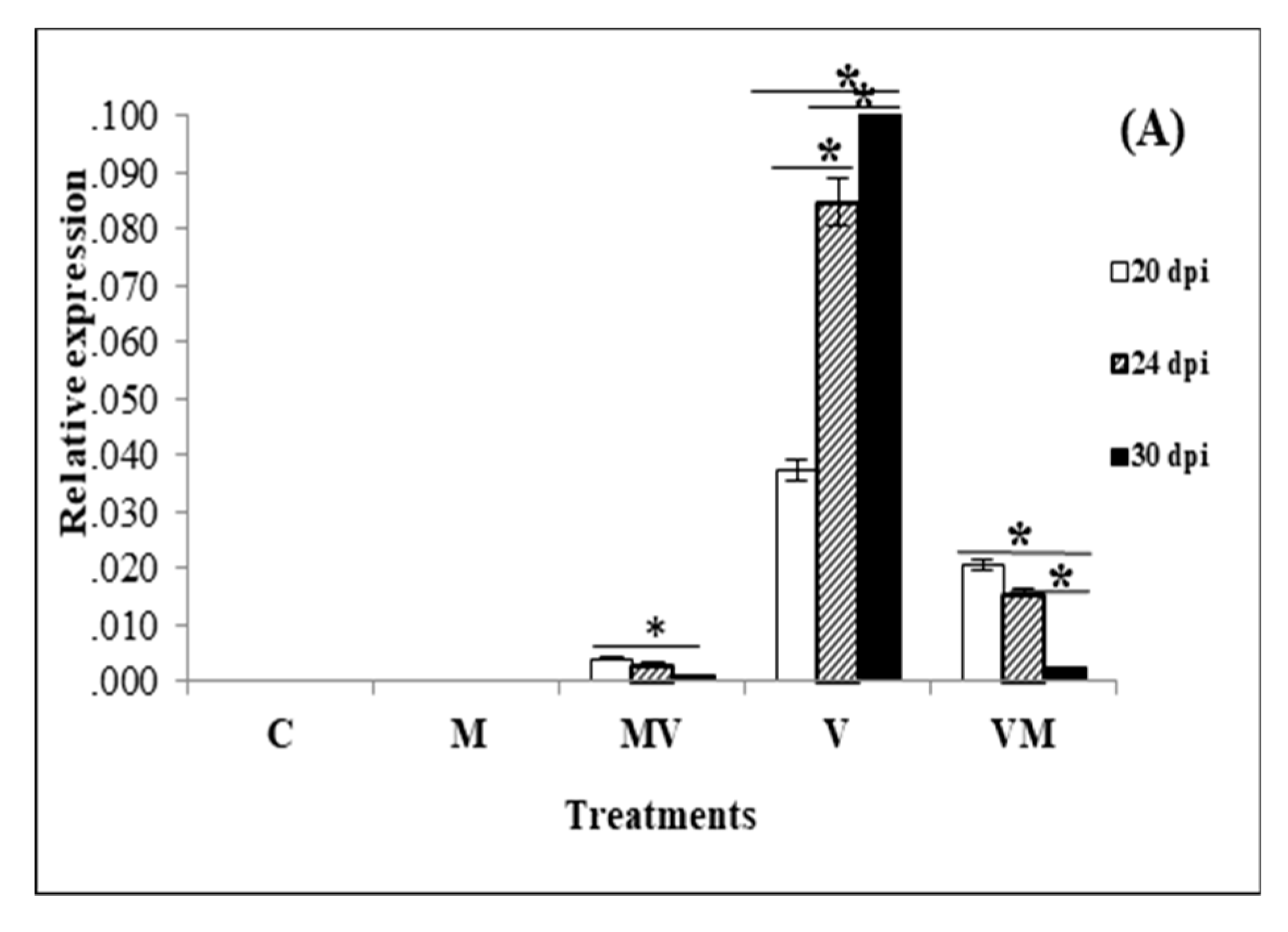
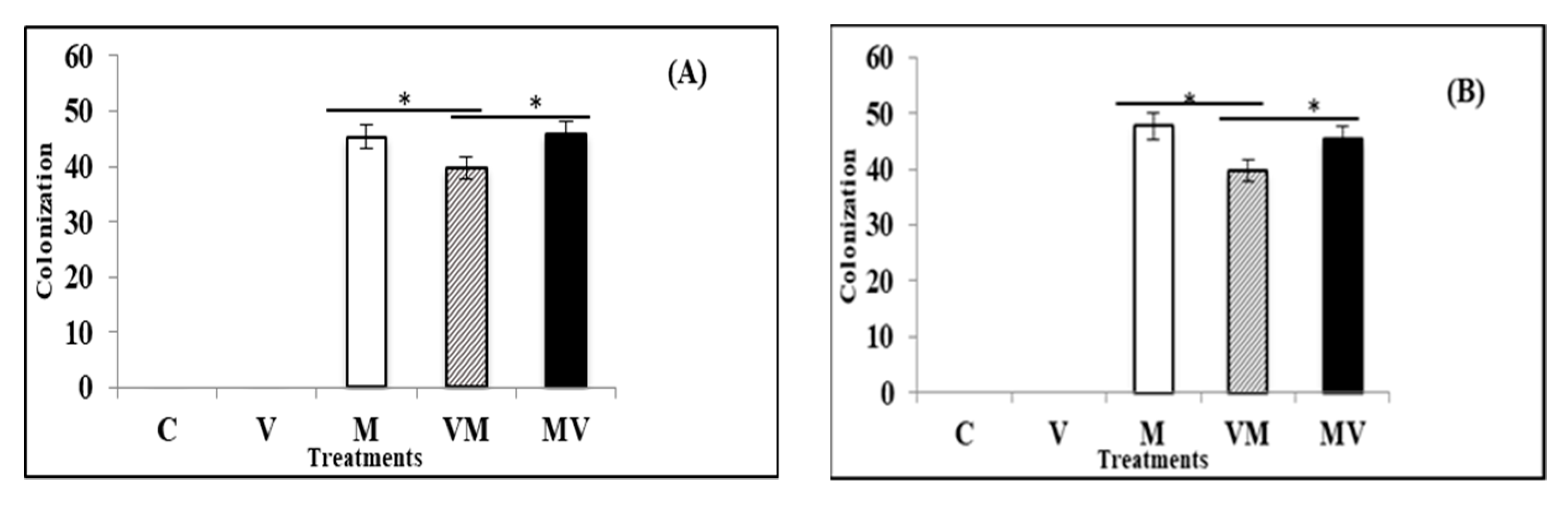
3.2. Effect of Mycorrhiza on the Viral Accumulation
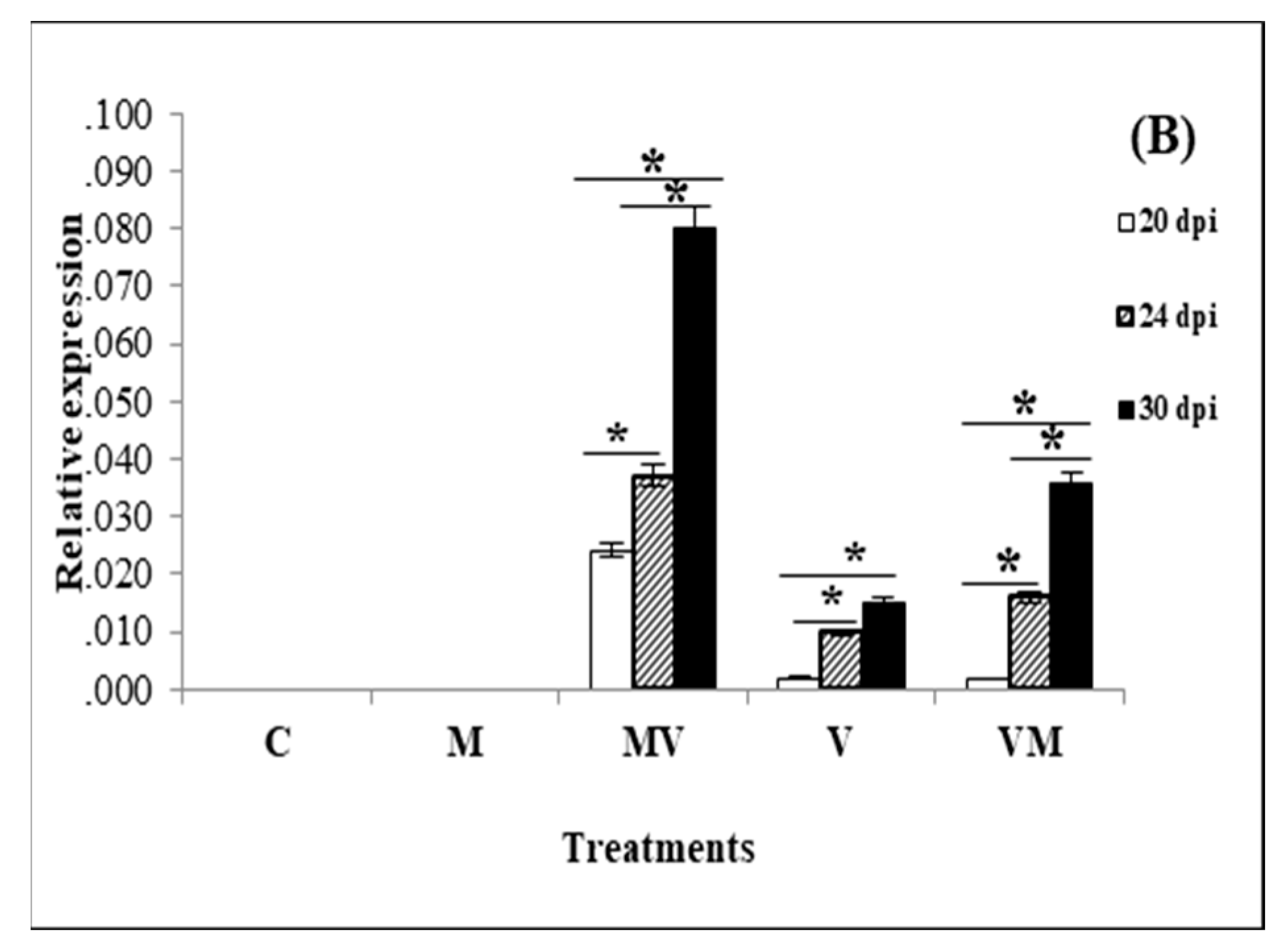
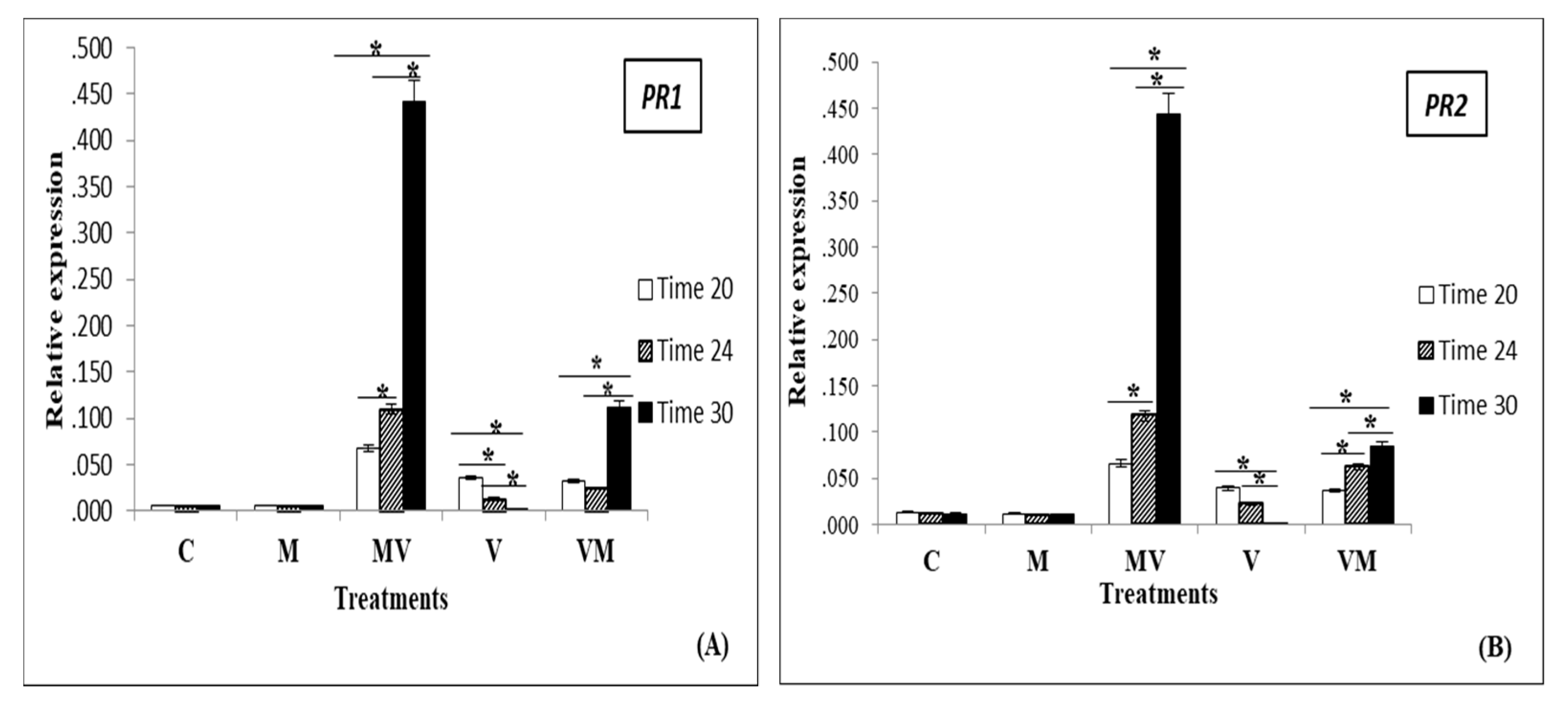
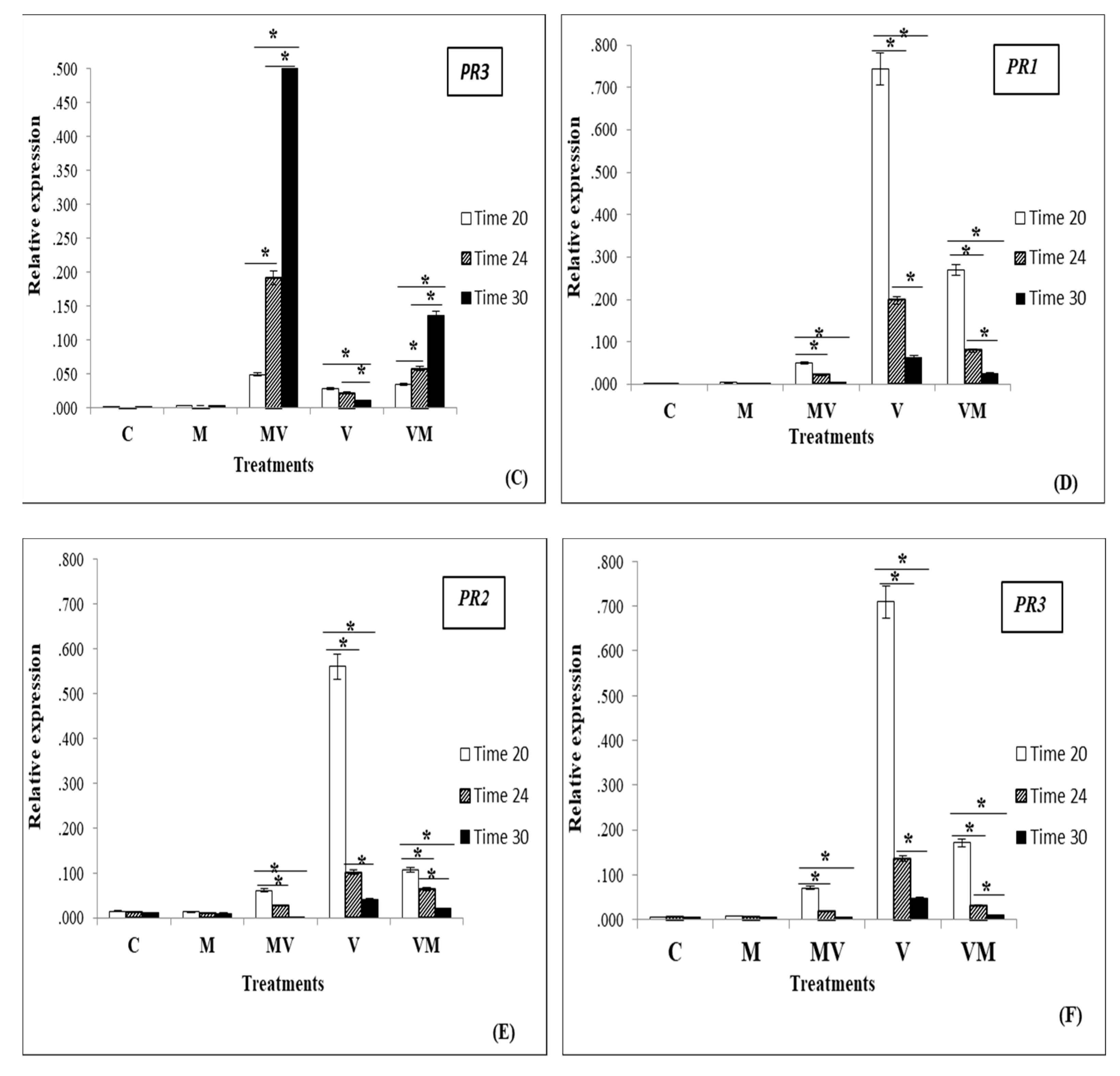
3.3. Regulation of Genes in Mycorrhizal Plants Infected by Viruses
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Augé, R.M.; Toler, H.D.; Saxton, A.M. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: A meta-analysis. Mycorrhiza 2015, 25, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Bona, E.; Scarafoni, A.; Marsano, F.; Boatti, L.; Copetta, A.; Massa, N.; Gamalero, E.; D’Agostino, G.; Cesaro, P.; Cavaletto, M.; et al. Arbuscular mycorrhizal symbiosis affects the grain proteome of Zea mays: A field study. Sci. Rep. 2016, 6, 26439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorilli, V.; Vannini, C.; Ortolani, F.; Garcia-Seco, D.; Chiapello, M.; Novero, M.; Domingo, G.; Terzi, V.; Morcia, C.; Bagnaresi, P.; et al. Omics approaches revealed how arbuscular mycorrhizal symbiosis enhances yield and resistance to leaf pathogen in wheat. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef]
- Fritz, M.; Jakobsen, I.; Lyngkjær, M.F.; Thordal-Christensen, H.; Pons-Kühnemann, J. Arbuscular mycorrhiza reduces susceptibility of tomato to Alternaria solani. Mycorrhiza 2006, 16, 413–419. [Google Scholar] [CrossRef]
- Fiorilli, V.; Catoni, M.; Francia, D.; Cardinale, F.; Lanfranco, L. The arbuscular mycorrhizal symbiosis reduces disease severity in tomato plants infected by Botrytis cinerea. Eur. J. Plant. Pathol. 2011, 93, 237–242. [Google Scholar]
- Pozo, M.J.; Jung, S.C.; López-Ráez, J.A.; Azcón-Aguilar, C. Impact of arbuscular mycorrhizal symbiosis on plant response to biotic stress: The role of plant defence mechanisms. In Arbuscular Mycorrhizas: Physiology and Function; Springer: Berlin/Heidelberg, Germany, 2010; pp. 193–207. [Google Scholar] [CrossRef]
- Liu, J.; Maldonado-Mendoza, I.; Lopez-Meyer, M.; Cheung, F.; Town, C.D.; Harrison, M.J. Arbuscular mycorrhizal symbiosis is accompanied by local and systemic alterations in gene expression and an increase in disease resistance in the shoots. Plant. J. 2007, 50, 529–544. [Google Scholar] [CrossRef]
- García-Chapa, M.; Batlle, A.; Laviña, A.; Camprubí, A.; Estaún, V.; Calvet, C. Tolerance increase to pear decline phytoplasma in mycorrhizal OHF-333 pear rootstock. Acta Hortic. 2004, 657, 437–441. [Google Scholar] [CrossRef]
- Lingua, G.; D’Agostino, G.; Massa, N.; Antosiano, M.; Berta, G. Mycorrhiza-induced differential response to a yellows disease in tomato. Mycorrhiza 2002, 12, 191–198. [Google Scholar] [CrossRef]
- Maffei, G.; Miozzi, L.; Fiorilli, V.; Novero, M.; Lanfranco, L.; Accotto, G.P. The arbuscular mycorrhizal symbiosis attenuates symptom severity and reduces virus concentration in tomato infected by Tomato yellow leaf curl Sardinia virus (TYLCSV). Mycorrhiza 2014, 24, 179–186. [Google Scholar] [CrossRef]
- Jabaji-Hare, S.; Stobbs, L. Electron microscopic examination of tomato roots coinfected with Glomus sp. and tobacco mosaic virus. Phytopathology 1984, 74, 277–279. [Google Scholar] [CrossRef]
- Daft, M.; Okusanya, B. Effect of endogone mycorrhiza on plant growth v. influence of infection on the multiplication of viruses in tomato, petunia and strawberry. New Phytol. 1973, 72, 975–983. [Google Scholar] [CrossRef]
- Miozzi, L.; Catoni, M.; Fiorilli, V.; Mullineaux, P.M.; Accotto, G.P.; Lanfranco, L. Arbuscular mycorrhizal symbiosis limits foliar transcriptional responses to viral infection and favors long-term virus accumulation. Mol. Plant. Microbe Interact. 2011, 24, 1562–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerik, J.; Duffus, J.; Perry, R.; Stenger, D.; Van Maren, A. Etiology of tomato plant decline in the California desert. Phytopathology 1990, 80, 1352–1356. [Google Scholar] [CrossRef]
- Luis-Arteaga, M.; Rodríguez-Cerezo, E.; Fraile, A.; Sáez, E.; García-Arenal, F. Different tomato bushy stunt virus strains that cause disease outbreaks in solanaceous crops in Spain. Phytopathology 1996, 86, 535–542. [Google Scholar] [CrossRef]
- Ohki, T.; Uematsu, S.; Lesemann, D.-E.; Honda, Y.; Tsuda, S.; Fujisawa, I. Characterization of Tomato bushy stunt virus newly isolated from nipplefruit (Solanum mammosum) in Japan. J. Gen. Plant. Pathol. 2005, 71, 74–79. [Google Scholar] [CrossRef]
- Lawrence, C.; Joosten, M.; Tuzun, S. Differential induction of pathogenesis-related proteins in tomato byAlternaria solaniand the association of a basic chitinase isozyme with resistance. Physiol Mol. Plant. Pathol. 1996, 48, 361–377. [Google Scholar] [CrossRef]
- Song, Y.; Chen, D.; Lu, K.; Sun, Z.; Zeng, R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant. Sci. 2015, 6, 786. [Google Scholar] [CrossRef] [Green Version]
- Fiorilli, V.; Catoni, M.; Miozzi, L.; Novero, M.; Accotto, G.P.; Lanfranco, L. Global and cell-type gene expression profiles in tomato plants colonized by an arbuscular mycorrhizal fungus. New Phytol. 2009, 975–987. [Google Scholar] [CrossRef]
- Shaul, O.; Galili, S.; Volpin, H.; Ginzberg, I.; Elad, Y.; Chet, I.; Kapulnik, Y. Mycorrhiza-induced changes in disease severity and PR protein expression in tobacco leaves. Mol. Plant. Microbe Interact. 1999, 12, 1000–1007. [Google Scholar] [CrossRef] [Green Version]
- Scagliusi, S.M.M.; Basu, S.K.; de Gouvea, J.A.; Vega, J. Comparison of two diagnostic methods for evaluation of Sugarcane yellow leaf virus concentration in Brazilian sugarcane cultivars. Funct. Plant Sci. Biotechnol. 2009, 3, 26–30. [Google Scholar]
- Esteban, R.; Balaguer, L.; Manrique, E.; de Casas, R.R.; Ochoa, R.; Fleck, I.; Pintó-Marijuan, M.; Casals, I.; Morales, D.; Jiménez, M.S.; et al. Alternative methods for sampling and preservation of photosynthetic pigments and tocopherols in plant material from remote locations. Photosynth Res. 2020, 101, 77–88. [Google Scholar] [CrossRef]
- Brederode, F.T.; Linthorst, H.J.M.; Bol, J.F. Differential induction of acquired resistance and PR gene expression in tobacco by virus infection, ethephon treatment, UV light and wounding. Plant Mol. Biol. 1991, 17, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.; Ochoa-Corona, R.; Lebas, B.; Timudo, O.; Stewart, F.; Alexander, B. Broad detection and diagnosis of viruses of the genus Tombusvirus by RT-PCR coupled to single strand conformation polymorphism analysis. In Proceedings of the 7th Australasian Plant Virology Workshop, Rottnest Island, Perth, Australia, 8–11 November 2006; p. 48. [Google Scholar]
- Boben, J.; Kramberger, P.; Petrovič, N.; Cankar, K.; Peterka, M.; Štrancar, A.; Ravnikar, M. Detection and quantification of Tomato mosaic virus in irrigation waters. Eur. J. Plant. Pathol. 2007, 118, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.I.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 − ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Friedmann, M.; Lapidot, M.; Cohen, S.; Pilowsky, M. A novel source of resistance to tomato yellow leaf curl virus exhibiting a symptomless reaction to viral infection. J. Am. Soc. Hortic. Sci. 1998, 123, 1004–1007. [Google Scholar] [CrossRef]
- Arunachalam, P.; Radhakrishnan, V.; Mathew, S.; Kumar, P. Reaction of bitter gourd genotypes against distortion mosaic virus. Int. J. Veg. Sci. 2002, 29, 55–57. [Google Scholar]
- Khoshnazar, F.; Eini, O. Response of tomato cultivars to agroinfection with Beet curly top Iran virus. J. Crop Prot. 2016, 5, 473–482. [Google Scholar] [CrossRef]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and vinegar, a simple staining technique for arbuscular-mycorrhizal fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [Green Version]
- Trouvelot, A.; Kough, J.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. Physiological and genetical aspects of mycorrhizae. In Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; pp. 217–221. [Google Scholar]
- Al-Askar, A.; Rashad, Y. Arbuscular mycorrhizal fungi: A biocontrol agent against common. Plant Pathol. J. 2010, 9, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Schouteden, N.; De Waele, D.; Panis, B.; Vos, C.M. Arbuscular mycorrhizal fungi for the biocontrol of plant-parasitic nematodes: A review of the mechanisms involved. Front. Microbiology 2015, 6, 1280. [Google Scholar] [CrossRef] [Green Version]
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular mycorrhizal fungi trigger transcriptional expression of flavonoid and chlorogenic acid biosynthetic pathways genes in tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sharkawy, H.H.; Rashad, Y.M.; Ibrahim, S.A. Biocontrol of stem rust disease of wheat using arbuscular mycorrhizal fungi and Trichoderma spp. Physiol. Mol. Plant Pathol. 2018, 103, 84–91. [Google Scholar] [CrossRef]
- Fakhro, A.; Andrade-Linares, D.R.; von Bargen, S.; Bandte, M.; Büttner, C.; Grosch, R.; Schwarz, D.; Franken, P. Impact of Piriformospora indica on tomato growth and on interaction with fungal and viral pathogens. Mycorrhiza 2010, 20, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Cutt, J.; Klessig, D. Pathogenesis-related proteins. In Genes Involved in Plant Defense. Arch. Virol. 1992, 6, 1529–1541. [Google Scholar]
- Van Loon, L.; Van Strien, E. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Liang, G.H.; Trick, H.N.; Gill, B.S. Pathogenesis-related proteins and their genes in cereals. Plant Cell Tissue Organ Cult. 2001, 64, 93. [Google Scholar] [CrossRef]
- Góngora-Castillo, E.; Ibarra-Laclette, E.; Trejo-Saavedra, D.L.; Rivera-Bustamante, R.F. Transcriptome analysis of symptomatic and recovered leaves of geminivirus-infected pepper (Capsicum annuum). Virol. J. 2012, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, Z.; Teng, K.; Lai, J.; Zhang, Y.; Huang, Y.; Li, Y.; Liang, L.; Wang, Y.; Chu, C.; et al. Up-regulation of LSB1/GDU3 affects geminivirus infection by activating the salicylic acid pathway. Plant J. 2010, 62, 12–23. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Eini, O.; Koolivand, D. Arbuscular mycorrhizal symbiosis enhances virus accumulation and attenuates resistance-related gene expression in tomato plants infected with Beet curly top Iran virus. J. Plant Dis. Prot. 2020, 1–8. [Google Scholar] [CrossRef]
- Sipahioglu, M.H.; Demir, S.; Usta, M.; Akkopru, A. Biological relationship of Potato virus Y and arbuscular mycorrhizal fungus Glomus intraradices in potato. Pest Tech. 2009, 3, 63–66. [Google Scholar]
- Miozzi, L.; Vaira, A.M.; Catoni, M.; Fiorilli, V.; Accotto, G.P.; Lanfranco, L. Arbuscular mycorrhizal symbiosis: Plant friend or foe in the fight against viruses? Front. Microbiol. 2019, 10, 1238. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Control (C) | AMF (M) | AMF + Virus (MV) | Virus (V) | Virus + AMF (VM) |
|---|---|---|---|---|---|
| 0 days | Seed | Seed | Seed | Seed | Seed |
| 30 days | − | +AMF | +AMF | − | − |
| 50 days | − | − | +Virus | +Virus | +Virus |
| 60 days | − | − | − | − | +AMF |
| 70 days | Leaf sampling | Leaf sampling | Leaf sampling | Leaf sampling | Leaf sampling |
| 74 days | Leaf sampling | Leaf sampling | Leaf sampling | Leaf sampling | Leaf sampling |
| 80 days | Leaf sampling | Leaf sampling | Leaf sampling | Leaf sampling | Leaf sampling |
| Gene | Primer Sequence (5′ to 3′) | Accession No. | PCR Product Size |
|---|---|---|---|
| LePR1 | F: 5′-GCCAAGCTATAACTACGCTACCAAC-3′ | DQ159948 | 139 bp |
| R: 5′-GCAAGAAATGAACCACCATCC-3′ | |||
| LePR2 | F: 5′-GGACACCCTTCCGCTACTCTT-3′ | M80604 | 81 bp |
| R: 5′-TGTTCCTGCCCCTCCTTTC-3′ | |||
| LePR3 | F: 5′-AACTATGGGCCATGTGGAAGA-3′ | Z15140 | 81 bp |
| R: 5′-GGCTTTGGGGATTGAGGAG-3′ | |||
| LeUBI3 | F: 5′- TCCATCTCGTGCTCCGTCT-3′ | X58253 | 144 bp |
| R: 5′-GAACCTTTCCAGTGTCATCAACC-3′ |
| TBSV | ToMV | |||
|---|---|---|---|---|
| Treatment | Fresh Weight (g) M ± SD | Dry Weight (g) M ± SD | Fresh Weight (g) M ± SD | Dry Weight (g) M ± SD |
| C | 8.12 ± 0.39 a | 3.1 ± 0.30 a | 8.12 ± 0.39 a | 3.1 ± 0.30 a |
| M | 8.61 ± 0.30 a | 3.4 ± 0.38 a | 8.61 ± 0.30 a | 3.4 ± 0.38 a |
| MV | 7.18 ± 0.34 b | 2.07 ± 0.23 b | 5.48 ± 0.23 b | 1.11 ± 0.15 b |
| V | 5.93 ±0.51 c | 1.06 ± 0.23 c | 7.16 ± 0.49 d | 2.13 ± 0.43 c |
| VM | 7.08 ± 0.28 b | 2.01 ± 0.37 b | 6.13 ± 0.84 b | 1.16 ± 0.24 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khoshkhatti, N.; Eini, O.; Koolivand, D.; Pogiatzis, A.; Klironomos, J.N.; Pakpour, S. Differential Response of Mycorrhizal Plants to Tomato bushy stunt virus and Tomato mosaic virus Infection. Microorganisms 2020, 8, 2038. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122038
Khoshkhatti N, Eini O, Koolivand D, Pogiatzis A, Klironomos JN, Pakpour S. Differential Response of Mycorrhizal Plants to Tomato bushy stunt virus and Tomato mosaic virus Infection. Microorganisms. 2020; 8(12):2038. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122038
Chicago/Turabian StyleKhoshkhatti, Neda, Omid Eini, Davoud Koolivand, Antreas Pogiatzis, John N. Klironomos, and Sepideh Pakpour. 2020. "Differential Response of Mycorrhizal Plants to Tomato bushy stunt virus and Tomato mosaic virus Infection" Microorganisms 8, no. 12: 2038. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122038