Rumen and Hindgut Bacteria Are Potential Indicators for Mastitis of Mid-Lactating Holstein Dairy Cows

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experiment Design
2.3. Sample Collection and Analysis
2.4. DNA Extraction and Sequencing
2.5. Sequence Analyses
2.6. Statistical Analyses
3. Results
3.1. Performance and Rumen Fermentation
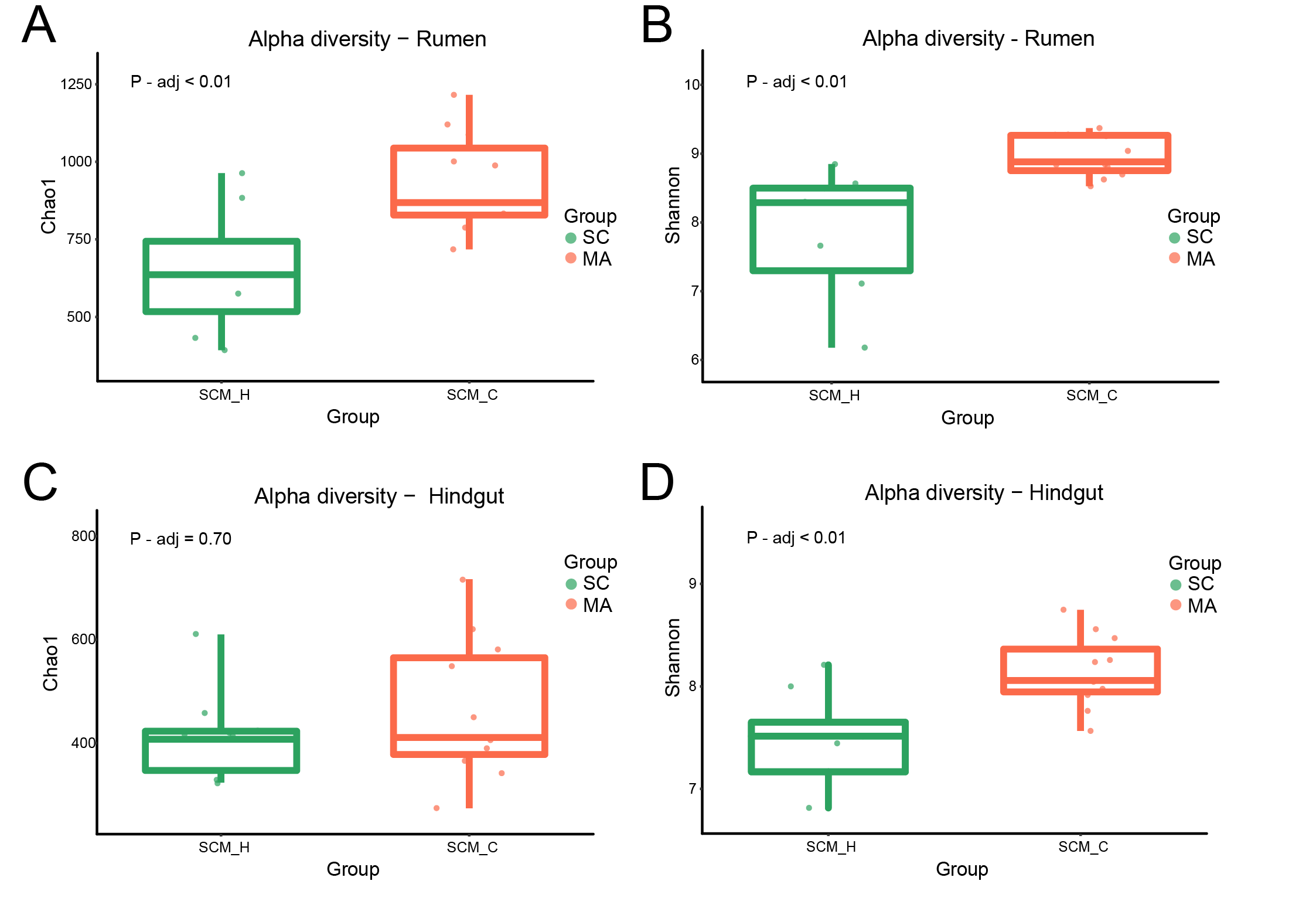
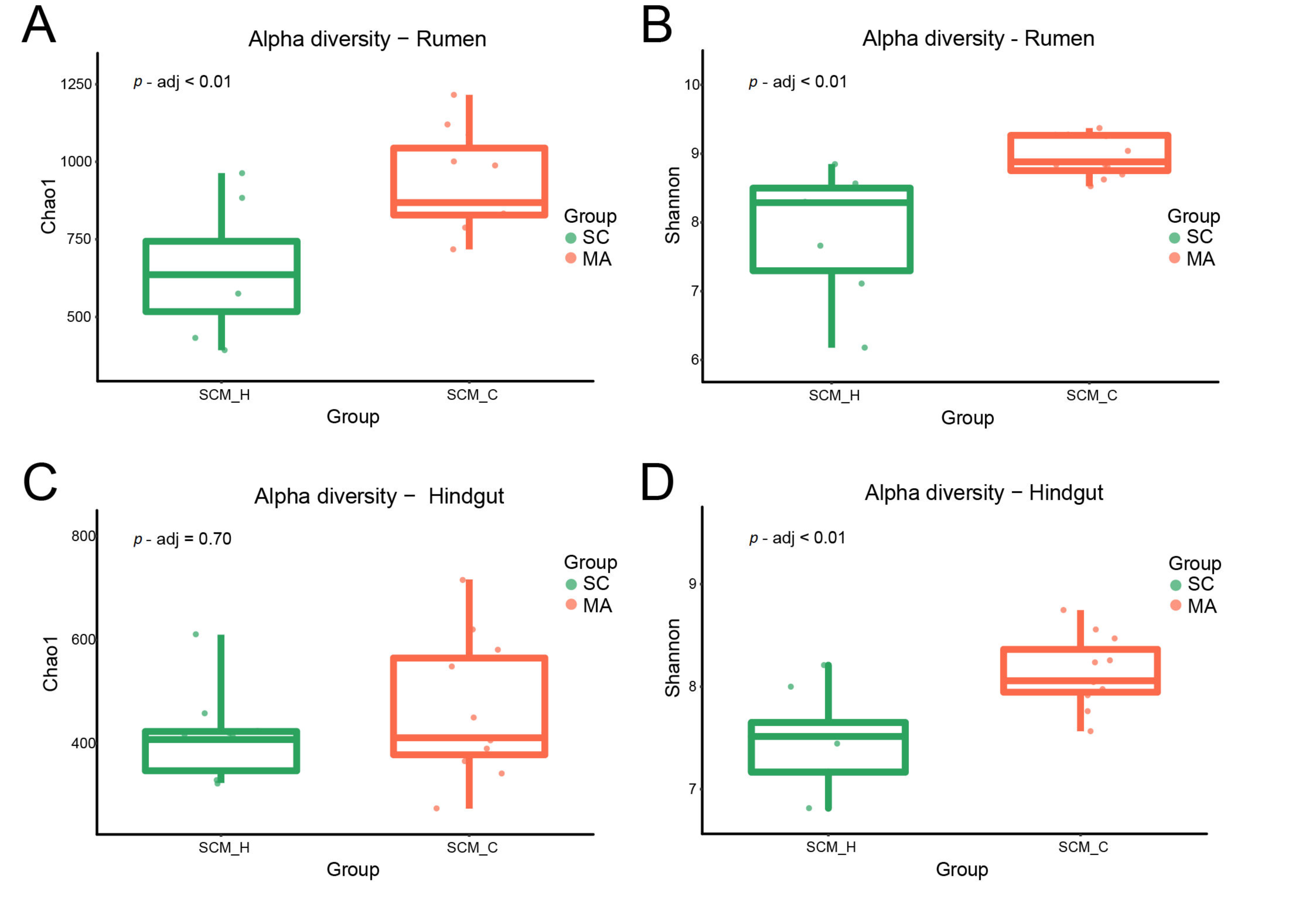
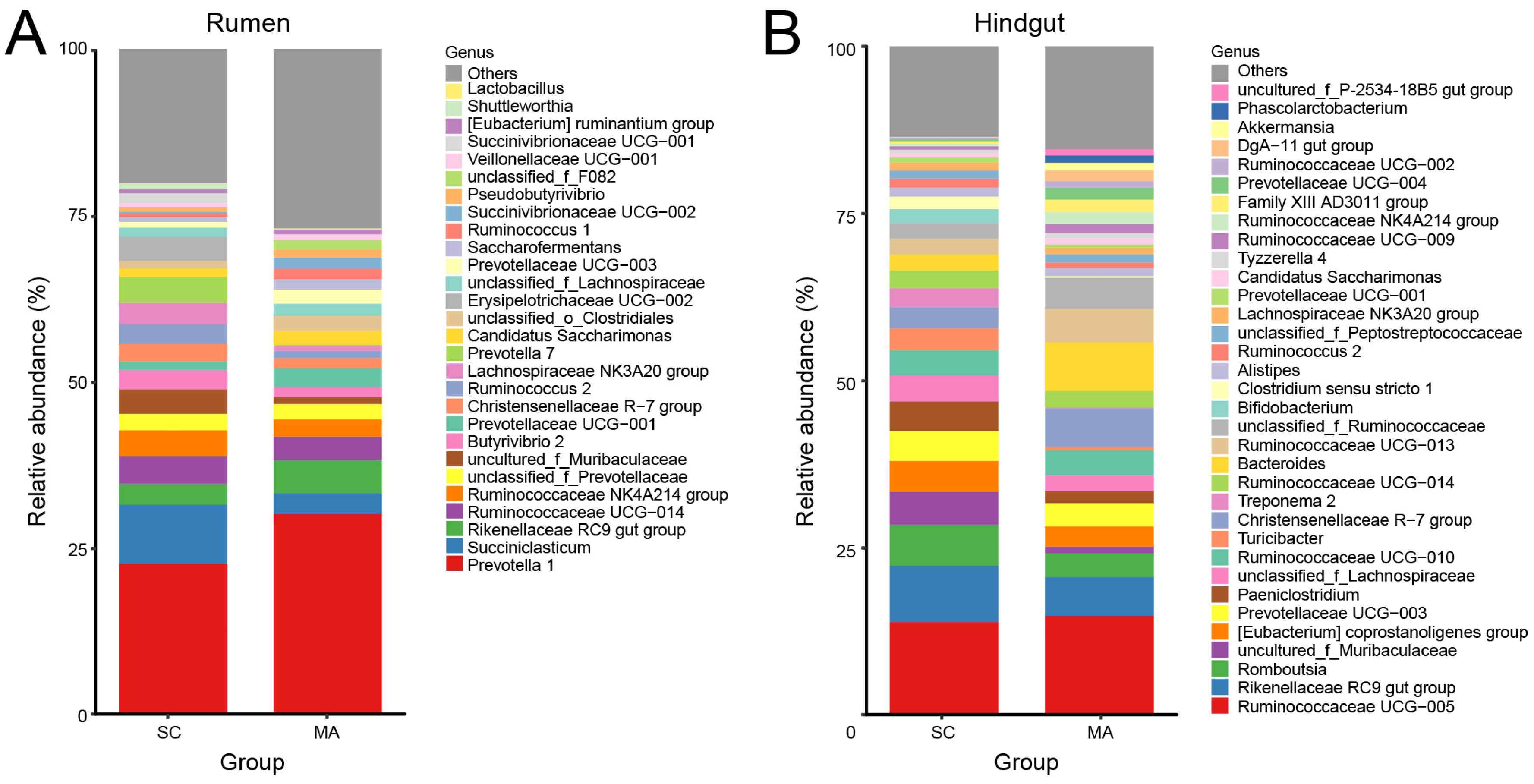
3.2. Rumen and Hindgut Bacteria Communities
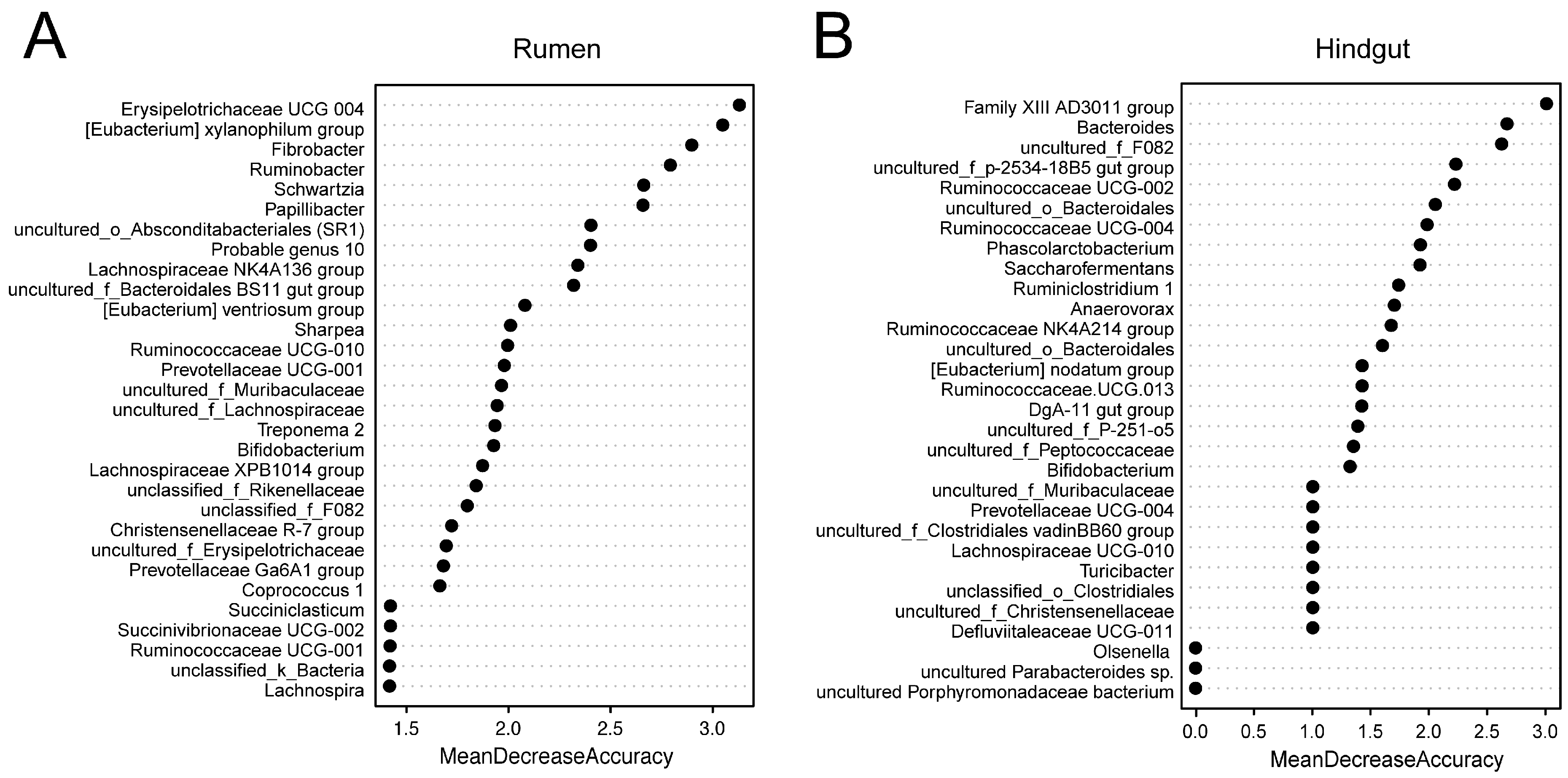
3.3. Random Forest Models of Observed Rumen and Hindgut Bacterial Genera
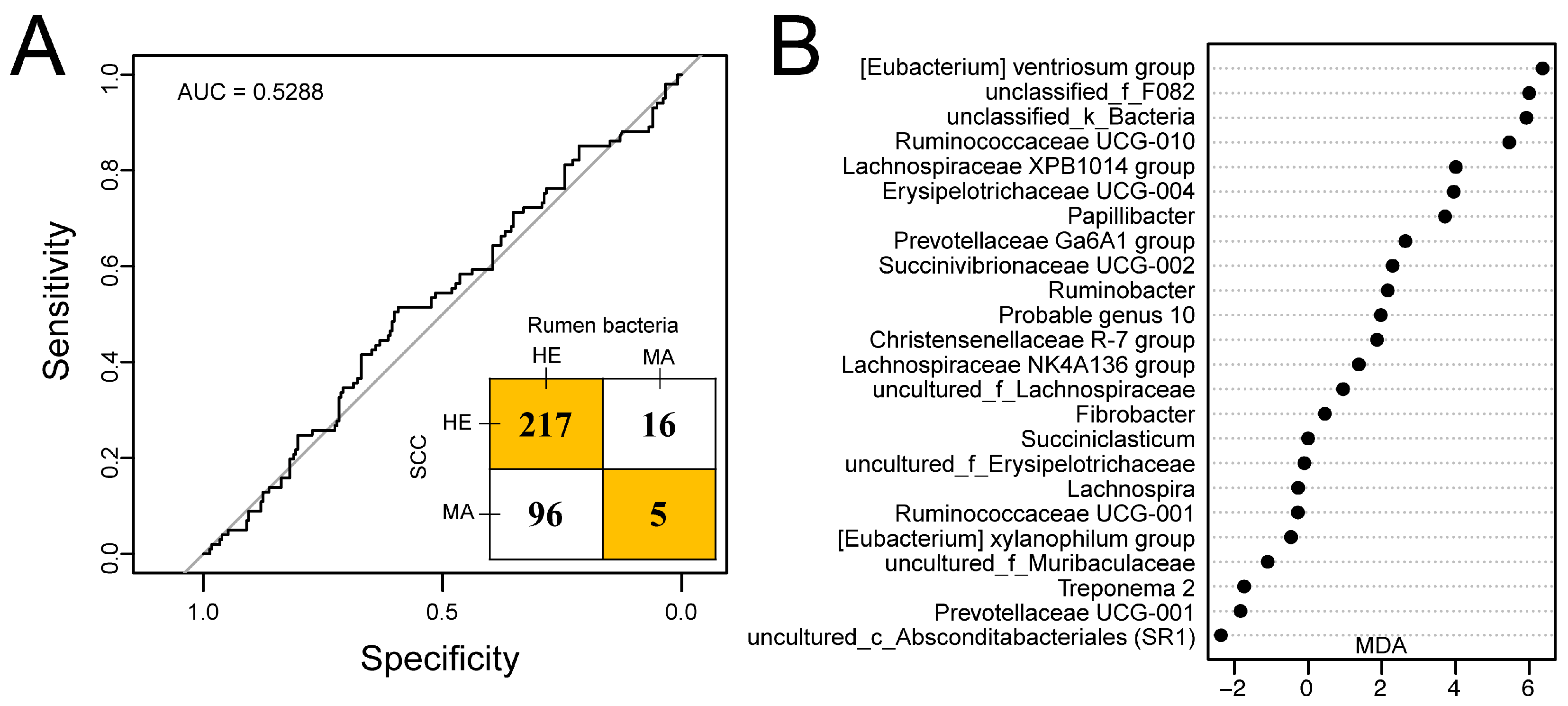
3.4. Predicting Mastitis Using Rumen Bacteria
4. Discussion
4.1. Differences Between Cows with High SCC
4.2. Random Forest Model and Potential Biomarker
4.3. Comparison of SCC and Rumen Bacteria Identification for Mastitis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Verteramo, C.L.J.; Tauer, L.W.; Gröhn, Y.T.; Smith, R.L. Mastitis risk effect on the economic consequences of paratuberculosis control in dairy cattle: A stochastic modeling study. PLoS ONE 2019, 14, e0217888. [Google Scholar] [CrossRef] [PubMed]
- Huijps, K.; Hogeveen, H.; Lam, T.J.; Oude Lansink, A.G. Costs and efficacy of management measures to improve udder health on Dutch dairy farms. J. Dairy Sci. 2010, 93, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, S.; Virchow, F.; Torgerson, P.R.; Bischoff, M.; Biner, B.; Hartnack, S.; Rüegg, S.R. Test characteristics of milk amyloid A ELISA, somatic cell count, and bacteriological culture for detection of intramammary pathogens that cause subclinical mastitis. J. Dairy Sci. 2017, 100, 7419–7426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezzat, A.M.; Quintela, B.M.; Böhme, K.; Fernández-No, I.; Caamaño-Antelo, S.; Calo-Mata, P.; Barros-Velázquez, J. The Immunology of Mammary Gland of Dairy Ruminants between Healthy and Inflammatory Conditions. J. Vet. Med. 2014, 2014, 659801. [Google Scholar]
- Rainard, P.; Foucras, G.; Boichard, D.; Rupp, R. Invited review: Low milk somatic cell count and susceptibility to mastitis. J. Dairy Sci. 2018, 101, 6703–6714. [Google Scholar] [CrossRef] [Green Version]
- Alhussien, M.N.; Dang, A.K. Milk somatic cells, factors influencing their release, future prospects, and practical utility in dairy animals: An overview. Vet. World 2018, 11, 562–577. [Google Scholar] [CrossRef]
- Viguier, C.; Arora, S.; Gilmartin, N.; Welbeck, K.; O’Kennedy, R. Mastitis detection: Current trends and future perspectives. Trends Biotechnol. 2009, 27, 486–493. [Google Scholar] [CrossRef]
- Reksen, O.; Sølverød, L.; Østerås, O. Relationships between milk culture results and composite milk somatic cell counts in Norwegian dairy cattle. J. Dairy Sci. 2008, 91, 3102–3113. [Google Scholar] [CrossRef]
- Sah, K.; Karki, P.; Shrestha, R.D.; Sigdel, A.; Adesogan, A.T.; Dahl, G.E. MILK Symposium review: Improving control of mastitis in dairy animals in Nepal*. J. Dairy Sci. 2020, 103, 9740–9747. [Google Scholar] [CrossRef]
- Shaheen, M.; Tantary, H.A.; Nabi, S.U. A Treatise on Bovine Mastitis: Disease and Disease Economics, Etiological Basis, Risk Factors, Impact on Human Health, Therapeutic Management, Prevention and Control Strategy. Adv. Dairy Res. 2015, 4, 1. [Google Scholar]
- Chakraborty, S.; Dhama, K.; Tiwari, R.; Iqbal, Y.M.; Khurana, S.K.; Khandia, R.; Munjal, A.; Munuswamy, P.; Kumar, M.A.; Singh, M.; et al. Technological interventions and advances in the diagnosis of intramammary infections in animals with emphasis on bovine population-a review. Vet. Q. 2019, 39, 76–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poore, G.D.; Kopylova, E.; Zhu, Q.; Carpenter, C.; Fraraccio, S.; Wandro, S.; Kosciolek, T.; Janssen, S.; Metcalf, J.; Song, S.J.; et al. Microbiome analyses of blood and tissues suggest cancer diagnostic approach. Nature 2020, 579, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; McClure, M.; Rorie, R.; Wang, X.; Chai, J.; Wei, X.; Lai, S.; Zhao, J. The vaginal and fecal microbiomes are related to pregnancy status in beef heifers. J. Anim. Sci. Biotechnol. 2019, 10, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.; Villot, C.; Renaud, D.; Skidmore, A.; Chevaux, E.; Steele, M.; Guan, L.L. Linking perturbations to temporal changes in diversity, stability, and compositions of neonatal calf gut microbiota: Prediction of diarrhea. ISME J. 2020, 14, 2223–2235. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Guo, J.; Zhao, C.; Jiang, P.; Maimai, T.; Yanyi, L.; Cao, Y.; Fu, Y.; Zhang, N. The gut microbiota contributes to the development of Staphylococcus aureus-induced mastitis in mice. ISME J. 2020, 14, 1897–1910. [Google Scholar] [CrossRef]
- Ma, C.; Sun, Z.; Zeng, B.; Huang, S.; Zhao, J.; Zhang, Y.; Su, X.; Xu, J.; Wei, H.; Zhang, H. Cow-to-mouse fecal transplantations suggest intestinal microbiome as one cause of mastitis. Microbiome 2018, 6, 200–217. [Google Scholar] [CrossRef]
- Zhong, Y.; Xue, M.; Liu, J. Composition of Rumen Bacterial Community in Dairy Cows With Different Levels of Somatic Cell Counts. Front. Microbiol. 2018, 9, 3217. [Google Scholar] [CrossRef] [Green Version]
- Addis, M.F.; Tanca, A.; Uzzau, S.; Oikonomou, G.; Bicalho, R.C.; Moroni, P. The bovine milk microbiota: Insights and perspectives from -omics studies. Mol. Biosyst. 2016, 12, 2359–2372. [Google Scholar] [CrossRef] [Green Version]
- Laporte, M.F.; Paquin, P. Near-infrared analysis of fat, protein, and casein in cow’s milk. J. Agric. Food Chem. 1999, 47, 2600–2605. [Google Scholar] [CrossRef]
- Shen, J.S.; Chai, Z.; Song, L.J.; Liu, J.X.; Wu, Y.M. Insertion depth of oral stomach tubes may affect the fermentation parameters of ruminal fluid collected in dairy cows. J. Dairy Sci. 2012, 95, 5978–5984. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.L.; Liu, J.X.; Ye, J.A.; Wu, Y.M.; Guo, Y.Q. Effect of tea saponin on rumen fermentation in vitro. Anim. Feed Sci. Technol. 2005, 120, 333–339. [Google Scholar] [CrossRef]
- Li, M.; Penner, G.B.; Hernandez-Sanabria, E.; Oba, M.; Guan, L.L. Effects of sampling location and time, and host animal on assessment of bacterial diversity and fermentation parameters in the bovine rumen. J. Appl. Microbiol. 2009, 107, 1924–1934. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, M.; Sun, H.; Wu, X.; Guan, L.L.; Liu, J. Assessment of Rumen Microbiota from a Large Dairy Cattle Cohort Reveals the Pan and Core Bacteriomes Contributing to Varied Phenotypes. Appl. Environ. Microbiol. 2018, 84, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.Z.; Xue, M.; Guan, L.L.; Liu, J. A collection of rumen bacteriome data from 334 mid-lactation dairy cows. Sci. Data 2019, 6, 180301. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Richoux, R.; Boutinaud, M.; Martin, P.; Gagnaire, V. Role of somatic cells on dairy processes and products: A review. Dairy Sci. Technol. 2014, 94, 517–538. [Google Scholar] [CrossRef] [Green Version]
- Kayano, M.; Itoh, M.; Kusaba, N.; Hayashiguchi, O.; Kida, K.; Tanaka, Y.; Kawamoto, K.; Gröhn, Y.T. Associations of the first occurrence of pathogen-specific clinical mastitis with milk yield and milk composition in dairy cows. J. Dairy Res. 2018, 85, 309–316. [Google Scholar] [CrossRef]
- Wall, S.K.; Hernández-Castellano, L.E.; Ahmadpour, A.; Bruckmaier, R.M.; Wellnitz, O. Differential glucocorticoid-induced closure of the blood-milk barrier during lipopolysaccharide- and lipoteichoic acid-induced mastitis in dairy cows. J. Dairy Sci. 2016, 99, 7544–7553. [Google Scholar] [CrossRef] [Green Version]
- Kester, H.J.; Sorter, D.E.; Hogan, J.S. Activity and milk compositional changes following experimentally induced Streptococcus uberis bovine mastitis. J. Dairy Sci. 2015, 98, 999–1004. [Google Scholar] [CrossRef] [Green Version]
- Pant, K.; Saraya, A.; Venugopal, S.K. Oxidative stress plays a key role in butyrate-mediated autophagy via Akt/mTOR pathway in hepatoma cells. Chem. Biol. Interact. 2017, 273, 99–106. [Google Scholar] [CrossRef]
- Claesson, M.J.; Jeffery, I.B.; Conde, S.; Power, S.E.; O’Connor, E.M.; Cusack, S.; Harris, H.M.B.; Coakley, M.; Lakshminarayanan, B.; O’Sullivan, O.; et al. Gut microbiota composition correlates with diet and health in the elderly. Nature 2012, 488, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda-Varas, P.; Proudfoot, K.L.; Weary, D.M.; von Keyserlingk, M.A.G. Changes in behaviour of dairy cows with clinical mastitis. Appl. Anim. Behav. Sci. 2016, 175, 8–13. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palm, N.W.; de Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W.; et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef] [Green Version]
- Dinh, D.M.; Volpe, G.E.; Duffalo, C.; Bhalchandra, S.; Tai, A.K.; Kane, A.V.; Wanke, C.A.; Ward, H.D. Intestinal microbiota, microbial translocation, and systemic inflammation in chronic HIV infection. J. Infect. Dis. 2015, 211, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gylswyk, N.O.; Hippe, H.; Rainey, F.A. Schwartzia succinivorans gen. nov., sp. nov., another ruminal bacterium utilizing succinate as the sole energy source. Int. J. Syst. Bacteriol. 1997, 47, 155–159. [Google Scholar] [CrossRef]
- Indugu, N.; Vecchiarelli, B.; Baker, L.D.; Ferguson, J.D.; Vanamala, J.K.P.; Pitta, D.W. Comparison of rumen bacterial communities in dairy herds of different production. BMC Microbiol. 2017, 17, 190. [Google Scholar] [CrossRef]
- Campbell, J.H.; O’Donoghue, P.; Campbell, A.G.; Schwientek, P.; Sczyrba, A.; Woyke, T.; Söll, D.; Podar, M. UGA is an additional glycine codon in uncultured SR1 bacteria from the human microbiota. Proc. Natl. Acad. Sci. USA 2013, 110, 5540–5545. [Google Scholar] [CrossRef] [Green Version]
- Hongoh, Y.; Sharma, V.K.; Prakash, T.; Noda, S.; Taylor, T.D.; Kudo, T.; Sakaki, Y.; Toyoda, A.; Hattori, M.; Ohkuma, M. Complete genome of the uncultured Termite Group 1 bacteria in a single host protist cell. Proc. Natl. Acad. Sci. USA 2008, 105, 5555–5560. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Haake, S.K.; Mannon, P.; Lemon, K.P.; Waldron, L.; Gevers, D.; Huttenhower, C.; Izard, J. Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome Biol. 2012, 13, R42. [Google Scholar] [CrossRef] [Green Version]
- Griffen, A.L.; Beall, C.J.; Campbell, J.H.; Firestone, N.D.; Kumar, P.S.; Yang, Z.K.; Podar, M.; Leys, E.J. Distinct and complex bacterial profiles in human periodontitis and health revealed by 16S pyrosequencing. ISME J. 2012, 6, 1176–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falentin, H.; Rault, L.; Nicolas, A.; Bouchard, D.S.; Lassalas, J.; Lamberton, P.; Aubry, J.M.; Marnet, P.G.; Le Loir, Y.; Even, S. Bovine Teat Microbiome Analysis Revealed Reduced Alpha Diversity and Significant Changes in Taxonomic Profiles in Quarters with a History of Mastitis. Front. Microbiol. 2016, 7, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, M.T.; Kenny, D.J.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Ruminococcus gnavus, a member of the human gut microbiome associated with Crohn’s disease, produces an inflammatory polysaccharide. Proc. Natl. Acad. Sci. USA 2019, 116, 12672–12677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Tailford, L.E.; Owen, C.D.; Walshaw, J.; Crost, E.H.; Hardy-Goddard, J.; Le Gall, G.; de Vos, W.M.; Taylor, G.L.; Juge, N. Discovery of intramolecular trans-sialidases in human gut microbiota suggests novel mechanisms of mucosal adaptation. Nat. Commun. 2015, 6, 7624. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Duan, Y.; Xu, Z.; Lyu, N.; Liu, F.; Liang, S.; Zhu, B. An examination of data from the American Gut Project reveals that the dominance of the genus Bifidobacterium is associated with the diversity and robustness of the gut microbiota. Microbiol. Open 2019, 8, e939. [Google Scholar] [CrossRef] [PubMed]
- Rupp, R.; Beaudeau, F.; Boichard, D. Relationship between milk somatic-cell counts in the first lactation and clinical mastitis occurrence in the second lactation of French Holstein cows. Prev. Vet. Med. 2000, 46, 99–111. [Google Scholar] [CrossRef]
- Suriyasathaporn, W.; Schukken, Y.H.; Nielen, M.; Brand, A. Low somatic cell count: A risk factor for subsequent clinical mastitis in a dairy herd. J. Dairy Sci. 2000, 83, 1248–1255. [Google Scholar] [CrossRef]
- Lipkens, Z.; Piepers, S.; De Visscher, A.; De Vliegher, S. Evaluation of test-day milk somatic cell count information to predict intramammary infection with major pathogens in dairy cattle at drying off. J. Dairy Sci. 2019, 102, 4309–4321. [Google Scholar] [CrossRef]
- Pers-Kamczyc, E.; Zmora, P.; Cieślak, A.; Szumacher, S.M. Development of nucleic acid based techniques and possibilities of their application to rumen microbial ecology research. J. Anim. Feed Sci. 2011, 20, 315–337. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Nguyen, V.T.; Suzuki, T.; Yamada, K.; Kudo, K.; Hikita, C.; Le, V.P.; Nguyen, M.C.; Yoshida, N. Network analysis and functional estimation of the microbiome reveal the effects of cashew nut shell liquid feeding on methanogen behaviour in the rumen. Micro. Biotech. 2020. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | SC 1 | MA 2 | SEM | p-Value |
|---|---|---|---|---|
| Parity | 1.89 | 2.18 | 0.21 | 0.42 |
| Days in milk | 165.67 | 166.27 | 5.33 | 0.88 |
| Somatic cell counts, 103/mL | 2892 | 2169 | 637.0 | 0.30 |
| Milk yield, kg/d | 21.51a | 9.82 b | 2.08 | <0.01 |
| Protein, % | 3.80 | 3.60 | 0.08 | 0.12 |
| Fat, % | 3.60 | 4.32 | 0.16 | 0.09 |
| Lactose, % | 4.22 a | 3.26 b | 0.21 | 0.04 |
| Milk urea nitrogen, mg/dL | 15.06 a | 7.44 b | 0.97 | <0.01 |
| Item | SC 1 | MA 2 | SEM | p-Value |
|---|---|---|---|---|
| Rumen pH | 6.63 | 6.56 | 0.06 | 0.69 |
| Ammonia nitrogen, mg/dL | 6.24 | 7.11 | 0.46 | 0.46 |
| Total volatile fatty acid, mmol/L | 85.10 | 83.96 | 3.97 | 0.37 |
| Molar proportion, mmol/100 mmol | ||||
| Acetate (A) | 64.67 b | 70.38 a | 0.77 | <0.01 |
| Propionate (P) | 19.30 | 17.48 | 0.45 | 0.10 |
| Butyrate | 11.86 a | 9.00 b | 0.46 | <0.01 |
| Isobutyrate | 1.08 | 0.86 | 0.07 | 0.13 |
| Valerate | 1.39 a | 1.10 b | 0.05 | 0.01 |
| Isovalerate | 1.69 a | 1.17 b | 0.11 | 0.02 |
| A:P ratio | 3.41 b | 4.04 a | 0.12 | 0.01 |
| Item | SC 1 | MA 2 | SEM | p-Value |
|---|---|---|---|---|
| Erysipelotrichaceae UCG-004 | 0.07 b | 0.30 a | 0.04 | <0.01 |
| [Eubacterium] xylanophilum group | 0.01 b | 0.17 a | 0.03 | <0.01 |
| Fibrobacter | 0.04 b | 0.33 a | 0.05 | <0.01 |
| Ruminobacter | 0.03 b | 0.53 a | 0.12 | <0.01 |
| Schwartzia | 0.74 a | 0.09 b | 0.12 | <0.01 |
| Papillibacter | 0.02 b | 0.39 a | 0.06 | <0.01 |
| uncultured_o_Absconditabacteriales (SR1) | 0.25 b | 0.86 a | 0.11 | <0.01 |
| probable Genus 10 | 0.07 b | 0.36 a | 0.05 | <0.01 |
| Lachnospiraceae NK4A136 group | 0.11 b | 0.41 a | 0.07 | <0.01 |
| uncultured_f_Bacteroidales BS11 gut group | 0.24 b | 0.65 a | 0.11 | 0.02 |
| [Eubacterium] ventriosum group | 0.05 b | 0.40 a | 0.06 | <0.01 |
| Sharpea | 0.17 | nd | 0.04 | - |
| Ruminococcaceae UCG-010 | 0.13 b | 0.61 a | 0.08 | <0.01 |
| Prevotellaceae UCG-001 | 1.26 b | 2.77 a | 0.28 | <0.01 |
| uncultured_f_Muribaculaceae | 0.52 | 0.71 | 0.13 | 0.65 |
| uncultured_f_Lachnospiraceae | 0.19 b | 0.45 a | 0.05 | <0.01 |
| Treponema 2 | 0.24 b | 1.03 a | 0.15 | <0.01 |
| Bifidobacterium | 0.48 a | 0.01 b | 0.11 | <0.01 |
| Lachnospiraceae XPB1014 group | 0.55 | 0.73 | 0.09 | 0.15 |
| unclassified_f_Rikenellaceae | 0.04 b | 0.43 a | 0.13 | <0.01 |
| unclassified_f_F082 | 0.27 b | 1.19 a | 0.16 | <0.01 |
| Christensenellaceae R-7 group | 2.69 | 1.54 | 0.41 | 0.54 |
| uncultured_f_Erysipelotrichaceae | 0.04 b | 0.33 a | 0.06 | <0.01 |
| Prevotellaceae Ga6A1 group | 0.12 b | 0.32 a | 0.04 | 0.01 |
| Coprococcus 1 | 0.13 | 0.04 | 0.02 | 0.25 |
| Succiniclasticum | 8.91 | 3.09 | 1.21 | 0.15 |
| Succinivibrionaceae UCG-002 | 0.24 b | 1.68 a | 0.35 | 0.01 |
| Ruminococcaceae UCG-001 | 0.05 | 0.25 | 0.05 | 0.07 |
| unclassified_k_Bacteria | 0.04 | 0.08 | 0.01 | 0.02 |
| Lachnospira | 0.47 a | 0.36 b | 0.11 | 0.02 |
| Item | SC 1 | MA 2 | SEM | p-Value |
|---|---|---|---|---|
| Family XIII AD3011 group | 0.36 b | 1.79 a | 0.19 | <0.01 |
| Bacteroides | 2.38 b | 7.29 a | 0.70 | <0.01 |
| uncultured_f_F082 | 0.02 | 0.54 | 0.07 | <0.01 |
| uncultured_f_p-2534-18B5 gut group | nd | 0.95 | 0.25 | - |
| Ruminococcaceae UCG-002 | 0.19 b | 0.98 a | 0.11 | <0.01 |
| uncultured_o_ Bacteroidales | 0.07 b | 0.59 a | 0.07 | <0.01 |
| Prevotellaceae UCG-004 | 0.34 b | 1.80 a | 0.24 | <0.01 |
| Phascolarctobacterium | 0.06 b | 1.09 a | 0.13 | <0.01 |
| Saccharofermentans | 0.01 b | 0.23 a | 0.03 | <0.01 |
| Ruminiclostridium 1 | 0.01 b | 0.22 a | 0.03 | <0.01 |
| Anaerovorax | nd | 0.28 | 0.04 | - |
| Ruminococcaceae NK4A214 group | 0.41 b | 1.84 a | 0.17 | <0.01 |
| uncultured_o_Bacteroidales | 0.01 | 0.05 | 0.02 | 0.76 |
| [Eubacterium] nodatum group | 0.04 b | 0.27 a | 0.04 | 0.01 |
| Ruminococcaceae UCG-013 | 2.37 b | 5.00 a | 0.44 | <0.01 |
| DgA-11 gut group | 0.08 b | 1.61 a | 0.21 | <0.01 |
| uncultured_f_P-251-o5 | nd | 0.25 | 0.03 | - |
| uncultured_f_Peptococcaceae | 0.03 b | 0.18 a | 0.02 | <0.01 |
| Bifidobacterium | 2.06 | nd | 0.50 | - |
| unclassified_f_Muribaculaceae | 0.42 a | 0.13 b | 0.06 | 0.02 |
| Prevotellaceae UCG-004 | 0.34 b | 1.80 a | 0.24 | <0.01 |
| uncultured_f_Clostridiales vadinBB60 group | 0.19 | 0.03 | 0.03 | 0.07 |
| Lachnospiraceae UCG-010 | 0.09 b | 0.63 a | 0.07 | <0.01 |
| Turicibacter | 3.29 a | 0.53 b | 0.48 | <0.01 |
| unclassified_o_Clostridiales | 0.12 b | 0.54 a | 0.06 | <0.01 |
| uncultured_f_Christensenellaceae | nd | 0.11 | 0.02 | - |
| Defluviitaleaceae UCG-011 | 0.08 b | 0.17 a | 0.02 | 0.01 |
| Olsenella | 0.14 | 0.34 | 0.07 | 0.20 |
| uncultured Parabacteroides sp. | 0.32 | nd | 0.14 | - |
| uncultured Porphyromonadaceae bacterium | 0.63 a | 0.08 b | 0.10 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Y.; Xue, M.-Y.; Sun, H.-Z.; Valencak, T.G.; Guan, L.L.; Liu, J. Rumen and Hindgut Bacteria Are Potential Indicators for Mastitis of Mid-Lactating Holstein Dairy Cows. Microorganisms 2020, 8, 2042. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122042
Zhong Y, Xue M-Y, Sun H-Z, Valencak TG, Guan LL, Liu J. Rumen and Hindgut Bacteria Are Potential Indicators for Mastitis of Mid-Lactating Holstein Dairy Cows. Microorganisms. 2020; 8(12):2042. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122042
Chicago/Turabian StyleZhong, Yifan, Ming-Yuan Xue, Hui-Zeng Sun, Teresa G. Valencak, Le Luo Guan, and Jianxin Liu. 2020. "Rumen and Hindgut Bacteria Are Potential Indicators for Mastitis of Mid-Lactating Holstein Dairy Cows" Microorganisms 8, no. 12: 2042. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8122042