Agroforestry Management Systems Drive the Composition, Diversity, and Function of Fungal and Bacterial Endophyte Communities in Theobroma Cacao Leaves

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Design
2.2. Sampling
2.3. Surface Sterilization and DNA Extraction
2.4. Amplification and Sequencing of 16S rRNA Genes
2.5. Amplification and Sequencing of the Fungal ITS Region
2.6. Processing of Bacterial and Fungal Datasets
2.7. Data Analysis
2.8. Core Community and Correlation-Based Indicator Species Analysis
2.9. Functional Predictions
2.10. Nucleotide Sequence Accession Numbers
3. Results
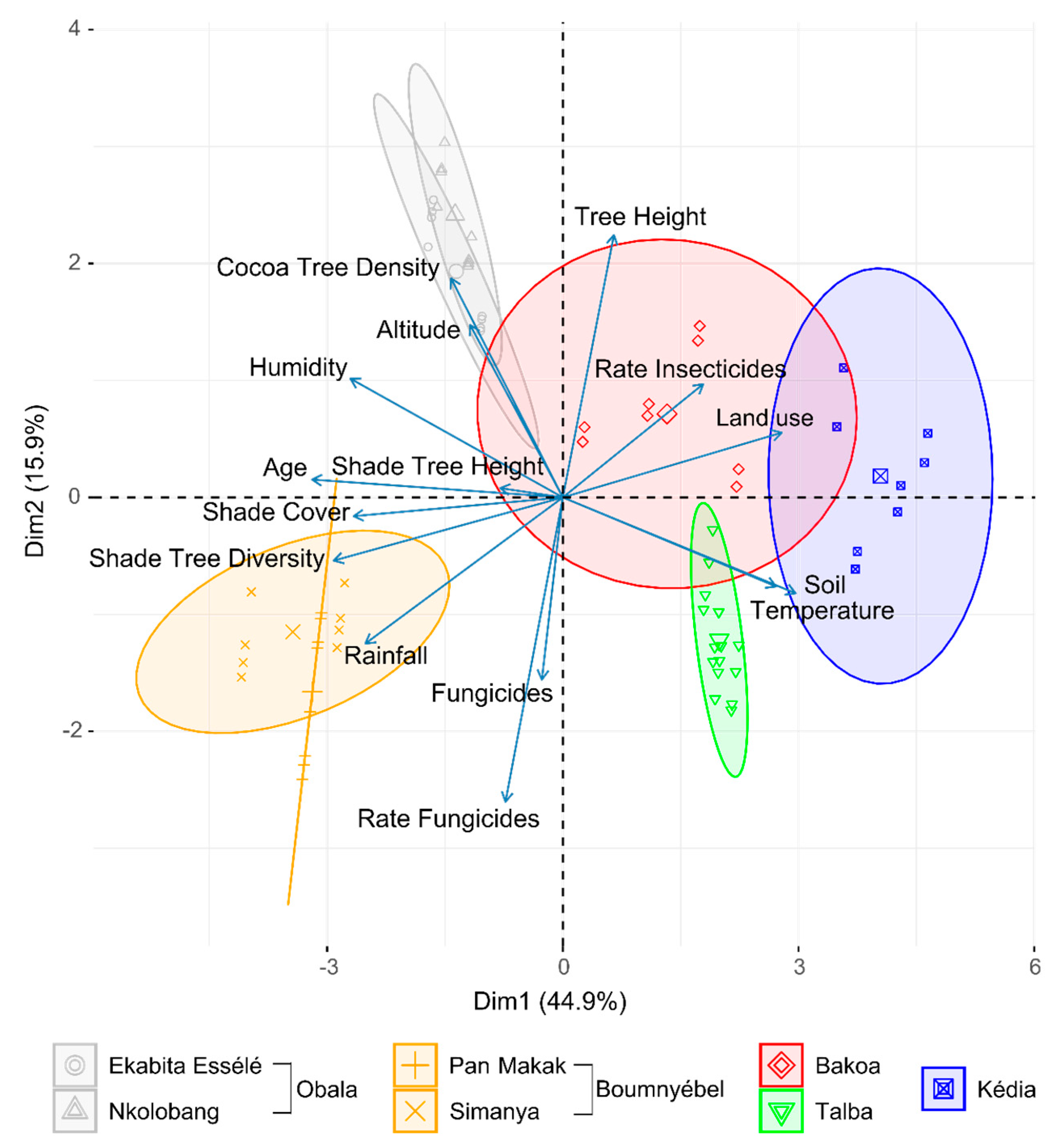
3.1. General Characteristics of the Investigated Sites
3.2. Sequence Characteristics
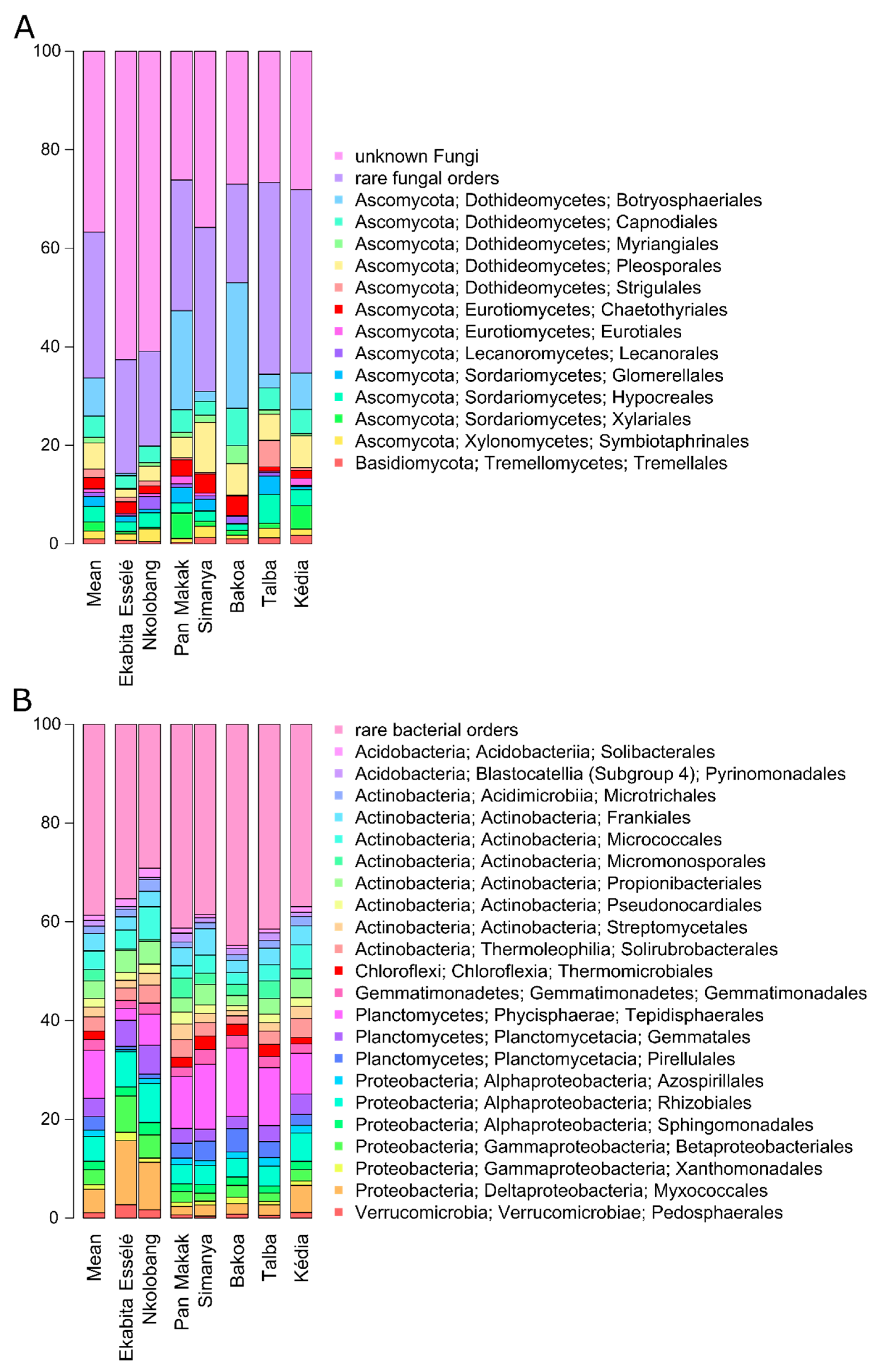
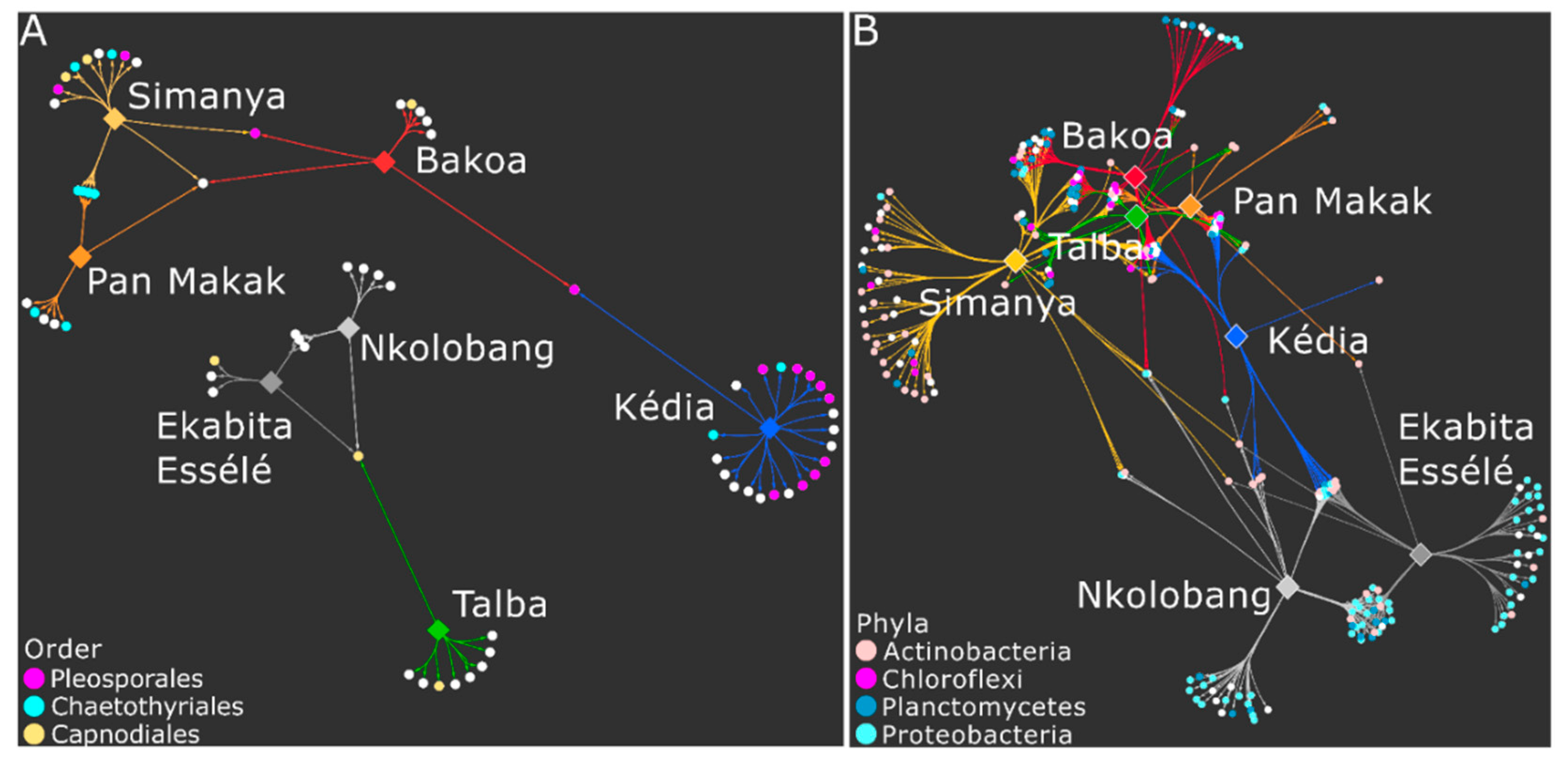
3.3. Foliar Endophyte Communities
3.4. Endophyte Diversity and Community Composition Per Site
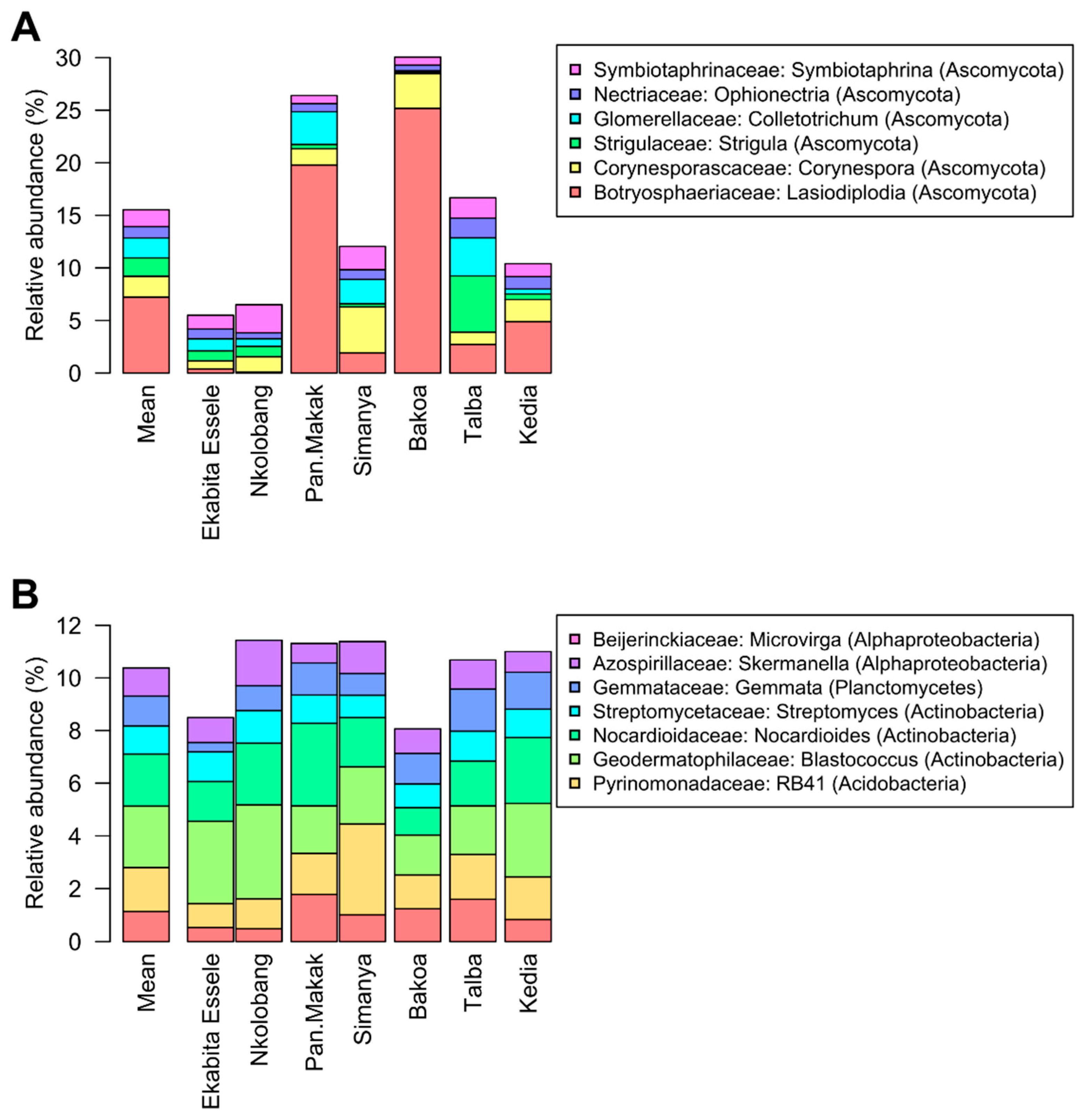
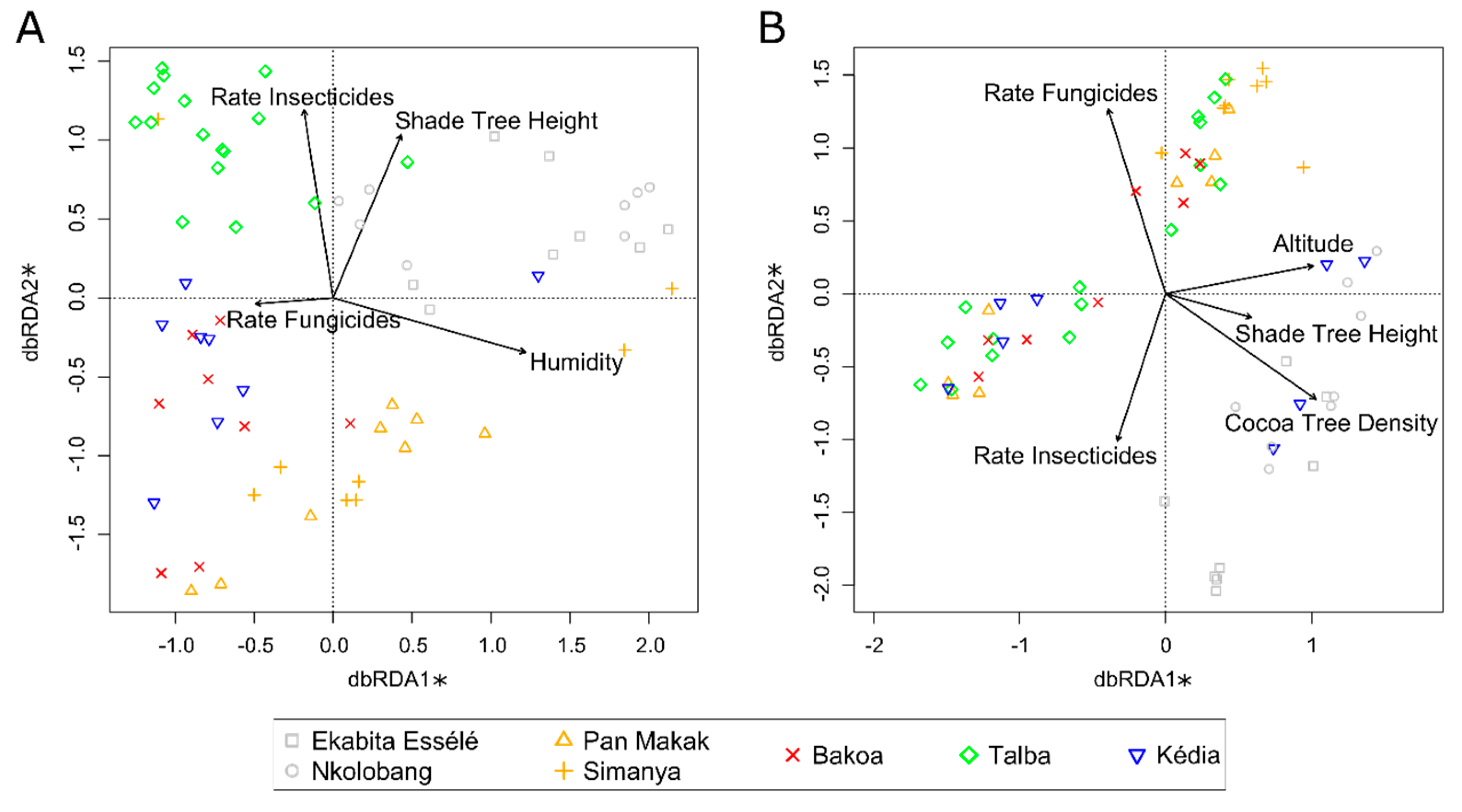
3.5. Bacterial and Fungal Taxa Associated with Agroforestry System Type
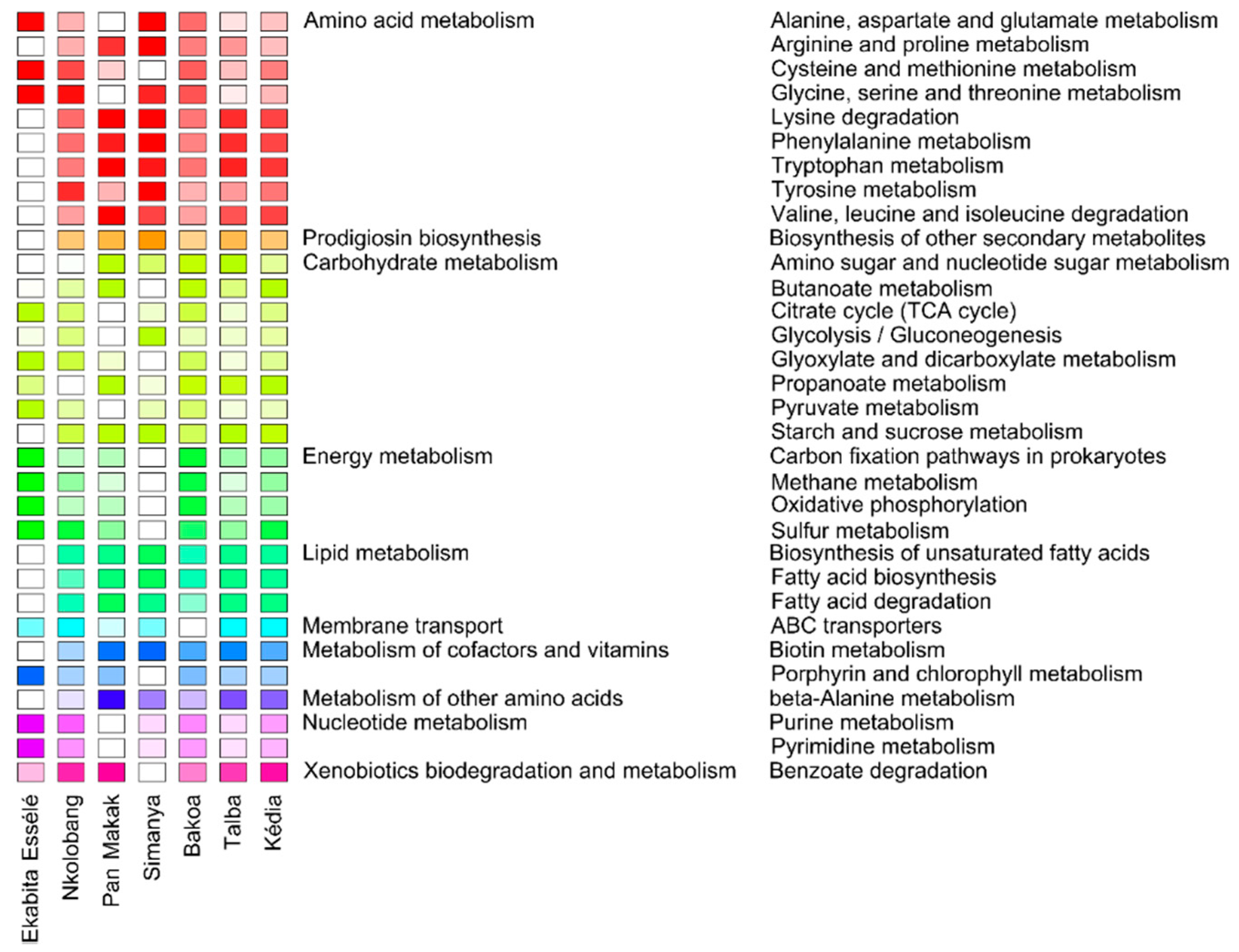
3.6. Fungal Functionality Differs between the Agroforest Management Systems
3.7. Predicted Functional Profiles of Bacterial Endophytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rice, R.A.; Greenberg, R. Cacao cultivation and the conservation of biological diversity. AMBIO J. Hum. Environ. 2000, 29, 167–173. [Google Scholar] [CrossRef]
- Clough, Y.; Faust, H.; Tscharntke, T. Cacao boom and bust: Sustainability of agroforests and opportunities for biodiversity conservation. Conserv. Lett. 2009, 2, 197–205. [Google Scholar] [CrossRef]
- Wade, A.S.; Asase, A.; Hadley, P.; Mason, J.; Ofori-Frimpong, K.; Preece, D.; Spring, N.; Norris, K. Management strategies for maximizing carbon storage and tree species diversity in cocoa-growing landscapes. Agric. Ecosyst. Environ. 2010, 138, 324–334. [Google Scholar] [CrossRef]
- Bisseleua, D.H.B.; Missoup, A.D.; Vidal, S. Biodiversity Conservation, Ecosystem Functioning, and Economic Incentives under Cocoa Agroforestry Intensification Conservación de Biodiversidad, Funcionamiento del Ecosistema e Incentivos Económicos en la Intensificación de la Agroforestería de Cacao. Conserv. Biol. 2009, 23, 1176–1184. [Google Scholar] [CrossRef]
- Bos, M.M.; Steffan-Dewenter, I.; Tscharntke, T. Shade tree management affects fruit abortion, insect pests and pathogens of cacao. Agric. Ecosyst. Environ. 2007, 120, 201–205. [Google Scholar] [CrossRef]
- Bisseleua, H.B.D.; Fotio, D.; Yede, A.D.M.; Vidal, S. Shade Tree Diversity, Cocoa Pest Damage, Yield Compensating Inputs and Farmers’ Net Returns in West Africa. PLoS ONE 2013, 8, e56115. [Google Scholar] [CrossRef]
- Bisseleua, D.H.B.; Vidal, S. Plant biodiversity and vegetation structure in traditional cocoa forest gardens in southern Cameroon under different management. Biodivers. Conserv. 2008, 17, 1821–1835. [Google Scholar] [CrossRef] [Green Version]
- Deheuvels, O.; Rousseau, G.X.; Quiroga, G.S.; Franco, M.D.; Cerda, R.; Mendoza, S.J.V.; Somarriba, E. Biodiversity is affected by changes in management intensity of cocoa-based agroforests. Agrofor. Syst. 2014, 88, 1081–1099. [Google Scholar] [CrossRef]
- Clough, Y.; Barkmann, J.; Juhrbandt, J.; Kessler, M.; Wanger, T.C.; Anshary, A.; Buchori, D.; Cicuzza, D.; Darras, K.; Putra, D.D.; et al. Combining high biodiversity with high yields in tropical agroforests. Proc. Natl. Acad. Sci. USA 2011, 108, 8311–8316. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttila, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Wemheuer, F.; Kaiser, K.; Karlovsky, P.; Daniel, R.; Vidal, S.; Wemheuer, B. Bacterial endophyte communities of three agricultural important grass species differ in their response towards management regimes. Sci. Rep. 2017, 7, 40914. [Google Scholar] [CrossRef] [PubMed]
- Hanada, R.E.; Pomella, A.W.V.; Costa, H.S.; Bezerra, J.L.; Loguercio, L.L.; Pereira, J.O. Endophytic fungal diversity in Theobroma cacao (cacao) and T. grandiflorum (cupuaçu) trees and their potential for growth promotion and biocontrol of black-pod disease. Fungal Biol. 2010, 114, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Saucedo-García, A.; Anaya, A.L.; Espinosa-García, F.; Gonzalez, M. Diversity and Communities of Foliar Endophytic Fungi from Different Agroecosystems of Coffea arabica L. in Two Regions of Veracruz, Mexico. PLoS ONE 2014, 9, e98454. [Google Scholar] [CrossRef]
- Granzow, S.; Kaiser, K.; Wemheuer, B.; Pfeiffer, B.; Daniel, R.; Vidal, S.; Wemheuer, F. The Effects of Cropping Regimes on Fungal and Bacterial Communities of Wheat and Faba Bean in a Greenhouse Pot Experiment Differ between Plant Species and Compartment. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejía, L.C.; Rojas, E.I.; Maynard, Z.; Bael, S.V.; Arnold, A.E.; Hebbar, P.; Samuels, G.J.; Robbins, N.; Herre, E.A. Endophytic fungi as biocontrol agents of Theobroma cacao pathogens. Biol. Control 2008, 46, 4–14. [Google Scholar] [CrossRef]
- Christian, N.; Herre, E.A.; Clay, K. Foliar endophytic fungi alter patterns of nitrogen uptake and distribution in Theobroma cacao. New Phytol. 2019, 222, 1573–1583. [Google Scholar] [CrossRef]
- Leite, H.A.C.; Silva, A.B.; Gomes, F.P.; Gramacho, K.P.; Faria, J.C.; de Souza, J.T.; Loguercio, L.L. Bacillus subtilis and Enterobacter cloacae endophytes from healthy Theobroma cacao L. trees can systemically colonize seedlings and promote growth. Appl. Microbiol. Biotechnol. 2013, 97, 2639–2651. [Google Scholar] [CrossRef]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant–endophyte–herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 108, 1. [Google Scholar]
- Arnold, A.E.; Mejia, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [Green Version]
- Melnick, R.L.; Suárez, C.; Bailey, B.A.; Backman, P.A. Isolation of endophytic endospore-forming bacteria from Theobroma cacao as potential biological control agents of cacao diseases. Biol. Control 2011, 57, 236–245. [Google Scholar] [CrossRef]
- Vaast, P.; Somarriba, E. Trade-offs between crop intensification and ecosystem services: The role of agroforestry in cocoa cultivation. Agrofor. Syst. 2014, 88, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Bowers, J.H.; Bailey, B.A.; Hebbar, P.K.; Sanogo, S.; Lumsden, R.D. The impact of plant diseases on world chocolate production. Plant Health Prog. 2001, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Marelli, J.-P.; Guest, D.I.; Bailey, B.A.; Evans, H.C.; Brown, J.K.; Junaid, M.; Barreto, R.W.; Lisboa, D.O.; Puig, A.S. Chocolate Under Threat from Old and New Cacao Diseases. Phytopathology 2019, 109, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- Wemheuer, B.; Thomas, T.; Wemheuer, F. Fungal Endophyte Communities of Three Agricultural Important Grass Species Differ in Their Response Towards Management Regimes. Microorganisms 2019, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.; Souza, R.; Lima, T.; Cavalcanti, M. Endophytic fungal diversity in coffee leaves (Coffea arabica) cultivated using organic and conventional crop management systems. Mycosphere 2014, 5, 523–530. [Google Scholar] [CrossRef]
- Christian, N.; Sullivan, C.; Visser, N.D.; Clay, K. Plant Host and Geographic Location Drive Endophyte Community Composition in the Face of Perturbation. Microbial. Ecol. 2016, 72, 621–632. [Google Scholar] [CrossRef]
- Wemheuer, F.; Wemheuer, B.; Kretzschmar, D.; Pfeiffer, B.; Herzog, S.; Daniel, R.; Vidal, S. Impact of grassland management regimes on bacterial endophyte diversity differs with grass species. Lett. Appl. Microbiol. 2016, 62, 323–329. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Wemheuer, B.; Wemheuer, F. Assessing bacterial and fungal diversity in the plants endosphere. In Metagenomics—Methods and Protocols, 2nd ed.; Streit, W., Daniel, R., Eds.; Humana Press: Totowa, NJ, USA, 2016. [Google Scholar]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Updating the 97% identity threshold for 16S ribosomal RNA OTUs. Bioinformatics 2018, 34, 2371–2375. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 16. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Ogle, D.H. FSA: Fisheries Stock Analysis. R Package Version 0.8.14. 2017. Available online: https://cran.r-project.org/web/packages/FSA/index.html (accessed on 25 August 2017).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.4.-4. 2017. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 25 August 2017).
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2016, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- Marcon, E.; Herault, B. Entropart: An R Package to Measure and Partition Diversity. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Tichy, L.; Chytry, M. Statistical determination of diagnostic species for site groups of unequal size. J. Veg. Sci. 2006, 17, 809–818. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: A R-based tool for the rapid prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene marker gene sequences. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Solis, M.J.L.; Dela Cruz, T.E.; Schnittler, M.; Unterseher, M. The diverse community of leaf-inhabiting fungal endophytes from Philippine natural forests reflects phylogenetic patterns of their host plant species Ficus benjamina, F. elastica and F. religiosa. Mycoscience 2016, 57, 96–106. [Google Scholar] [CrossRef]
- Wemheuer, F.; Wemheuer, B.; Daniel, R.; Vidal, S. Deciphering bacterial and fungal endophyte communities in leaves of two maple trees with green islands. Sci. Rep. 2019, 9, 14183. [Google Scholar] [CrossRef]
- Gazis, R.; Chaverri, P. Diversity of fungal endophytes in leaves and stems of wild rubber trees (Hevea brasiliensis) in Peru. Fungal Ecol. 2010, 3, 240–254. [Google Scholar] [CrossRef]
- Siddique, A.B.; Khokon, A.; Unterseher, M. What do we learn from cultures in the omics age? High-throughput sequencing and cultivation of leaf- inhabiting endophytes from beech (Fagus sylvatica L.) revealed complementary community composition but similar correlations with local habitat conditions. MycoKeys 2017, 20, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; Hardy, G.E.S.; Burgess, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley Region of Western Australia. Eur. J. Plant Pathol. 2011, 130, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Jakuschkin, B.; Fievet, V.; Schwaller, L.; Fort, T.; Robin, C.; Vacher, C. Deciphering the pathobiome: Intra-and interkingdom interactions involving the pathogen Erysiphe alphitoides. Microb. Ecol. 2016, 72, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Da Siva Fonseca, E.; Peixoto, R.S.; Rosado, A.S.; de Carvalho Balieiro, F.; Tiedje, J.M.; da Costa Rachid, C.T.C. The Microbiome of Eucalyptus Roots under Different Management Conditions and Its Potential for Biological Nitrogen Fixation. Microb. Ecol. 2018, 75, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Griffin, E.A.; Harrison, J.G.; Kembel, S.W.; Carrell, A.A.; Joseph Wright, S.; Carson, W.P. Plant host identity and soil macronutrients explain little variation in sapling endophyte community composition: Is disturbance an alternative explanation? J. Ecol. 2019, 107, 1876–1889. [Google Scholar] [CrossRef]
- Viaene, T.; Langendries, S.; Beirinckx, S.; Maes, M.; Goormachtig, S. Streptomyces as a plant’s best friend? FEMS Microb. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. Antonie Van Leeuwenhoek 2015, 108, 267–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, M.K.; Arnold, A.E.; Johnson, N.C. Factors influencing communities of foliar fungal endophytes in riparian woody plants. Fungal Ecol. 2013, 6, 365–378. [Google Scholar] [CrossRef]
- Köberl, M.; Dita, M.; Martinuz, A.; Staver, C.; Berg, G. Agroforestry leads to shifts within the gammaproteobacterial microbiome of banana plants cultivated in Central America. Front. Microbiol. 2015, 6, 91. [Google Scholar]
- Helander, M.; Wäli, P.; Kuuluvainen, T.; Saikkonen, K. Birch leaf endophytes in managed and natural boreal forests. Can. J. For. Res. 2006, 36, 3239–3245. [Google Scholar] [CrossRef]
- Zimmerman, N.B.; Vitousek, P.M. Fungal endophyte communities reflect environmental structuring across a Hawaiian landscape. Proc. Natl. Acad. Sci. USA 2012, 109, 13022–13027. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Herre, E.A. Canopy cover and leaf age affect colonization by tropical fungal endophytes: Ecological pattern and process in Theobroma cacao (Malvaceae). Mycologia 2003, 95, 388–398. [Google Scholar] [CrossRef]
- Moulas, C.; Petsoulas, C.; Rousidou, K.; Perruchon, C.; Karas, P.; Karpouzas, D.G. Effects of systemic pesticides imidacloprid and metalaxyl on the phyllosphere of pepper plants. Biomed. Res. Int. 2013, 2013, 969750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, I.; Friberg, H.; Steinberg, C.; Persson, P. Fungicide effects on fungal community composition in the wheat phyllosphere. PLoS ONE 2014, 9, e111786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerkenaar, A.; Sijpesteijn, A.K. Antifungal activity of metalaxyl and furalaxyl. Pest. Biochem. Physiol. 1981, 15, 71–78. [Google Scholar] [CrossRef]
- Wilson, D. Ecology of woody plant endophytes. In Microbial Endophytes; Bacon, C.W., White, J.F., Eds.; Marcel Dekker, Inc: New York, NY, USA, 2000; pp. 389–420. [Google Scholar]
- Scholtysik, A.; Unterseher, M.; Otto, P.; Wirth, C. Spatio-temporal dynamics of endophyte diversity in the canopy of European ash (Fraxinus excelsior). Mycol. Prog. 2012, 12. [Google Scholar] [CrossRef]
- Rodriguez, R.; White, J., Jr.; Arnold, A.E.; Redman, A.R.A. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W. Host species identity, site and time drive temperate tree phyllosphere bacterial community structure. Microbiome 2016, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Elsas, J.D.V. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef]
- Card, S.; Johnson, L.; Teasdale, S.; Caradus, J. Deciphering endophyte behaviour: The link between endophyte biology and efficacious biological control agents. FEMS Microbiol. Ecol. 2016, 92, fiw114. [Google Scholar] [CrossRef] [Green Version]
- Coutinho, I.B.L.; Freire, F.C.O.; Lima, C.S.; Lima, J.S.; Gonçalves, F.J.T.; Machado, A.R.; Silva, A.M.S.; Cardoso, J.E. Diversity of genus Lasiodiplodia associated with perennial tropical fruit plants in northeastern Brazil. Plant Pathol. 2017, 66, 90–104. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Z.; Cai, L.; Hyde, K.; Yu, Z.; McKenzie, E. Colletotrichum anthracnose of Amaryllidaceae. Fungal Divers. 2009, 39, 123–146. [Google Scholar]
- Rossman, A.Y. The Genus Ophionectria (Euascomycetes, Hypocreales). Mycologia 1977, 69, 355–391. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden Fungi, Emergent Properties: Endophytes and Microbiomes. Ann. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega, F.E.; Simpkins, A.; Aime, M.C.; Posada, F.; Peterson, S.W.; Rehner, S.A.; Infante, F.; Castillo, A.; Arnold, A.E. Fungal endophyte diversity in coffee plants from Colombia, Hawai’i, Mexico and Puerto Rico. Fungal Ecol. 2010, 3, 122–138. [Google Scholar] [CrossRef]
- Cilliers, A.J.; Swart, W.J.; Wingfield, M.J. A Review of Lasiodiplodia theobromae with Particular Reference to its Occurrence on Coniferous Seeds. S. Afr. For. J. 1993, 166, 47–52. [Google Scholar] [CrossRef]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Sutherland, E.D.; Lockwood, J.L. Hyperparasitism of oospores of some Peronosporales by Actinoplanes missouriensis and Humicola fuscoatra and other Actinomycetes and fungi. Can. J. Plant Pathol. 1984, 6, 139–145. [Google Scholar] [CrossRef]
- Hirsch, A.; Valdes, M. Micromonospora: An important microbe for biomedicine and potentially for biocontrol and biofuels. Soil Biol. Biochem. 2010, 42, 536–542. [Google Scholar] [CrossRef]
- Bensch, K.; Braun, U.; Groenewald, J.Z.; Crous, P.W. The genus Cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef] [Green Version]
- Baral, H.O.; Weber, E.; Marson, G.; Quijada, L. A new connection between wood saprobism and beetle endosymbiosis: The rarely reported saprobic discomycete Tromeropsis is congeneric with the symbiotic yeast Symbiotaphrina (Symbiotaphrinales, Xylonomycetes) and two asexual morphs misplaced in Hyphozyma. Mycol. Prog. 2018, 17, 215–254. [Google Scholar] [CrossRef]
- Barnes, C.J.; Maldonado, C.; Frøslev, T.G.; Antonelli, A.; Rønsted, N. Unexpectedly High Beta-Diversity of Root-Associated Fungal Communities in the Bolivian Andes. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, S.; van der Lelie, D.; Hoffman, A.; Zhang, Y.-B.; Walla, M.D.; Vangronsveld, J.; Newman, L.; Monchy, S. Genome Sequence of the Plant Growth Promoting Endophytic Bacterium Enterobacter sp. 638. PLoS Genet. 2010, 6, e1000943. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Plantation | Trees | Agroforestry Management | Agricultural Land | Forest Land |
|---|---|---|---|---|---|
| Boumnyébel | Pan Makak 1 | 4 | extensively managed old (cocoa is grown under a dense cover of shade tree species) | 20% cocoa fields, 10% annual crop | 70% pristine forest, with forest reserve |
| Pan Makak 2 | 4 | ||||
| Simanya 1 | 4 | ||||
| Simanya 2 | 4 | ||||
| Talba | Talba 1 | 4 | intensively managed manure (cocoa is grown in larger farms) | 70% cocoa fields, 5% annual field crops | 25% pristine forest, no reserve |
| Talba 2 | 4 | ||||
| Talba 3 | 4 | ||||
| Talba 4 | 4 | ||||
| Bakoa | Bakoa 1 | 2 | extensively managed young (cocoa is grown on modified savannah agroecosystems) | 50% cocoa fields, 25% annual field crops, 5% patchy pasture fields | 20% secondary forest, no reserve |
| Bakoa 2 | 2 | ||||
| Bakoa 3 | 2 | ||||
| Bakoa 4 | 2 | ||||
| Kédia | Kédia 1 | 2 | intensively managed young (cocoa is grown under full sun) | 65% cocoa fields, 25% annual field crops, 5% pasture lands | 5% secondary forest |
| Kédia 2 | 2 | ||||
| Kédia 3 | 2 | ||||
| Kédia 4 | 2 | ||||
| Obala | Ekabita Essélé 1 | 4 | home garden cacao forest (cocoa is grown nearby houses with a high variety of fruit tree species) | 70% cocoa fields, 25% annual crop fields of mixed crops, agroforestry trees | 5% secondary forest, no forest reserve |
| Ekabita Essélé 2 | 4 | ||||
| Nkolobang 1 | 4 | ||||
| Nkolobang 2 | 4 |
| DF | SumOfSqs | F | p Value | ||
|---|---|---|---|---|---|
| Bacteria | |||||
| Entire Model | 6 | 3.89 | 2.30 | 0.001 | *** |
| Cocoa Tree Density | 1 | 0.49 | 1.72 | 0.024 | * |
| Altitude | 1 | 0.70 | 2.47 | 0.002 | ** |
| Fungicides rate/Cropping Season | 1 | 0.51 | 1.82 | 0.022 | * |
| Fungicides | 1 | 0.70 | 2.49 | 0.002 | ** |
| Shade Tree Height | 1 | 0.56 | 1.97 | 0.013 | * |
| Insecticides rate/Cropping Season | 1 | 0.45 | 1.58 | 0.049 | * |
| Residual | 57 | 16.08 | |||
| Fungi | |||||
| Entire Model | 5 | 4.33 | 2.31 | 0.001 | *** |
| Humidity | 1 | 1.05 | 2.78 | 0.001 | *** |
| Shade Tree Height | 1 | 0.70 | 1.87 | 0.006 | ** |
| Fungicides rate/Cropping Season | 1 | 0.71 | 1.90 | 0.005 | ** |
| Fungicides | 1 | 0.58 | 1.54 | 0.019 | * |
| Insecticides rate/Cropping Season | 1 | 0.55 | 1.48 | 0.042 | * |
| Residual | 58 | 21.77 | |||
| Fungi | Bacteria | |||
|---|---|---|---|---|
| Richness | Diversity | Richness | Diversity | |
| Obala | 178 ± 76A | 3.04 ± 0.98A | 1729 ± 218 | 7.02 ± 0.31 |
| - Ekabita Essélé | 174 ± 72a | 2.85 ± 0.98a | 1672 ± 255 | 6.92 ± 0.4 |
| - Nkolobang | 182 ± 85a | 3.24 ± 0.99 | 1785 ± 172 | 7.11 ± 0.18 |
| Boumnyébel | 279 ± 106B | 4.09 ± 1.31B | 1820 ± 85 | 7.17 ± 0.1 |
| - Pan Makak | 250 ± 110b | 3.96 ± 1.47 | 1814 ± 88 | 7.16 ± 0.09 |
| - Simanya | 307 ± 101b | 4.22 ± 1.23 | 1826 ± 87 | 7.19 ± 0.1 |
| Bakoa | 231 ± 82AB | 3.48 ± 1.11AB | 1789 ± 160 | 7.15 ± 0.19 |
| Talba | 292 ± 55Bb | 4.44 ± 0.41Bb | 1804 ± 137 | 7.17 ± 0.13 |
| Kédia | 330 ± 87Bb | 4.42 ± 0.64B | 1782 ± 101 | 7.12 ± 0.11 |
| Tested Variable | Fungal Endophytes | Bacterial Endophytes | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Richness | Diversity | Richness | Diversity | ||||||
| Environmental properties | DF | rho | p | rho | p | rho | p | rho | p |
| Altitude | 62 | −0.11 | 0.37 | −0.10 | 0.43 | 0.14 | 0.27 | 0.08 | 0.56 |
| Temperature | 62 | 0.34 | 0.006 | 0.26 | 0.04 | −0.06 | 0.65 | −0.01 | 0.92 |
| Humidity | 62 | −0.34 | 0.006 | −0.26 | 0.04 | 0.02 | 0.87 | −0.11 | 0.41 |
| Rainfall | 62 | 0.12 | 0.34 | 0.24 | 0.052 | 0.04 | 0.77 | 0.04 | 0.73 |
| Agroforestry management | |||||||||
| Age | 62 | −0.22 | 0.07 | −0.11 | 0.39 | 0.05 | 0.70 | 0.02 | 0.87 |
| Cacao Tree Density | 62 | −0.26 | 0.04 | −0.21 | 0.099 | −0.09 | 0.46 | −0.18 | 0.16 |
| Insecticides rate * | 62 | −0.06 | 0.62 | −0.05 | 0.70 | −0.08 | 0.51 | −0.09 | 0.49 |
| Fungicides rate * | 62 | 0.29 | 0.02 | 0.42 | <0.001 | 0.12 | 0.34 | 0.17 | 0.18 |
| Shade Tree Height | 62 | −0.14 | 0.26 | 0.004 | 0.97 | −0.18 | 0.16 | −0.17 | 0.18 |
| Shade Tree Diversity | 62 | −0.07 | 0.57 | 0.04 | 0.78 | 0.14 | 0.28 | 0.09 | 0.48 |
| Shade Cover | 62 | −0.18 | 0.16 | −0.07 | 0.58 | 0.02 | 0.86 | 0.02 | 0.88 |
| Tree Height | 62 | −0.27 | 0.03 | −0.29 | 0.02 | −0.03 | 0.83 | −0.02 | 0.85 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wemheuer, F.; Berkelmann, D.; Wemheuer, B.; Daniel, R.; Vidal, S.; Bisseleua Daghela, H.B. Agroforestry Management Systems Drive the Composition, Diversity, and Function of Fungal and Bacterial Endophyte Communities in Theobroma Cacao Leaves. Microorganisms 2020, 8, 405. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030405
Wemheuer F, Berkelmann D, Wemheuer B, Daniel R, Vidal S, Bisseleua Daghela HB. Agroforestry Management Systems Drive the Composition, Diversity, and Function of Fungal and Bacterial Endophyte Communities in Theobroma Cacao Leaves. Microorganisms. 2020; 8(3):405. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030405
Chicago/Turabian StyleWemheuer, Franziska, Dirk Berkelmann, Bernd Wemheuer, Rolf Daniel, Stefan Vidal, and Hervé Bertin Bisseleua Daghela. 2020. "Agroforestry Management Systems Drive the Composition, Diversity, and Function of Fungal and Bacterial Endophyte Communities in Theobroma Cacao Leaves" Microorganisms 8, no. 3: 405. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030405