Optimised Production and Extraction of Astaxanthin from the Yeast Xanthophyllomyces dendrorhous

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Culture Media
2.2. Bioreactor Fermentations
2.3. Yeast Cell Preparation for Extraction
2.4. Enzymatic Cell Lysis and Astaxanthin Extraction
2.5. Supercritical CO2 Extraction of Astaxanthin
2.6. Analytical Methods
2.7. Scanning Electron Microscopy
3. Results and Discussion
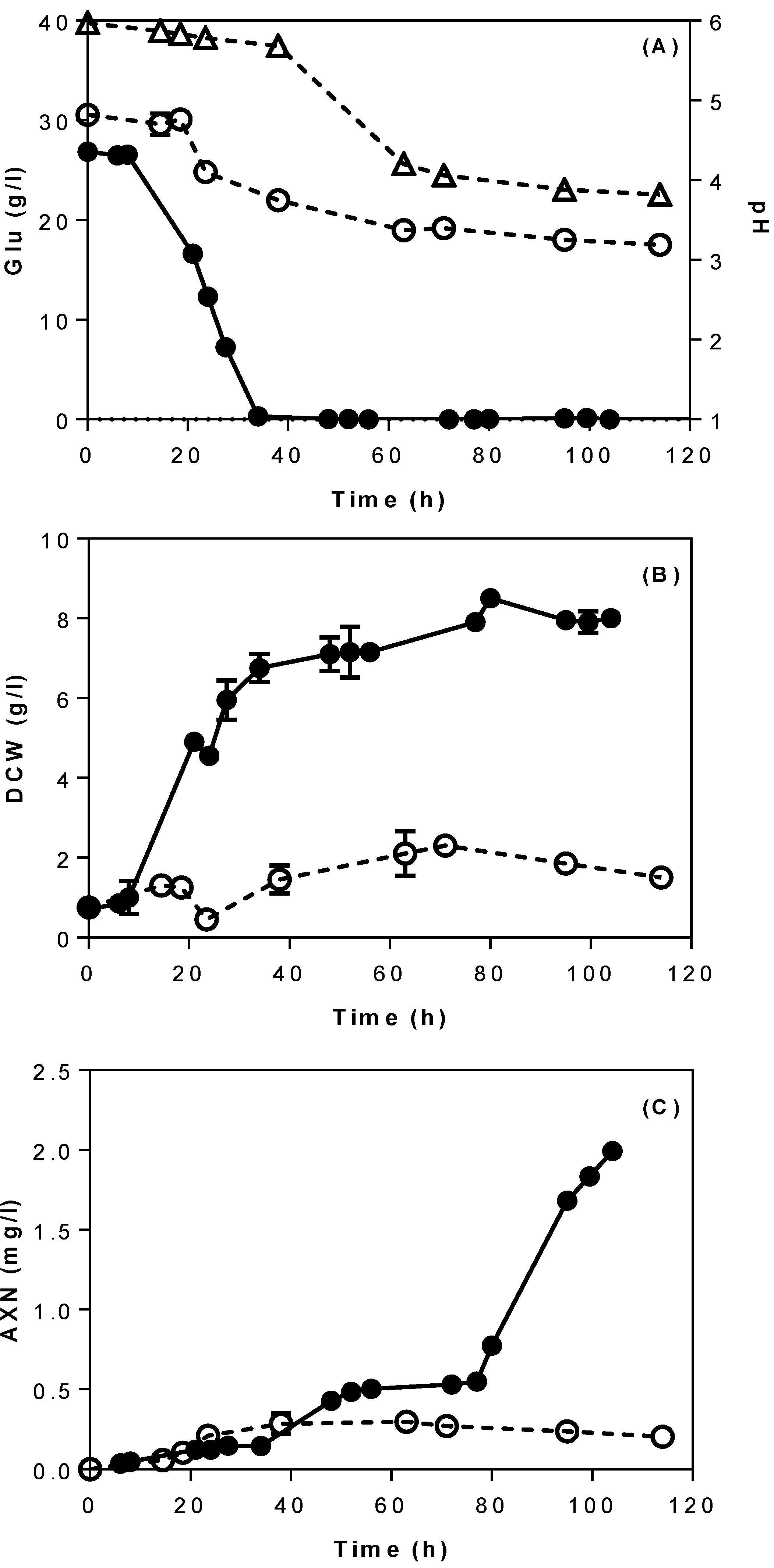
3.1. Batch Bioreactor Fermentations Using Semi-Defined Media
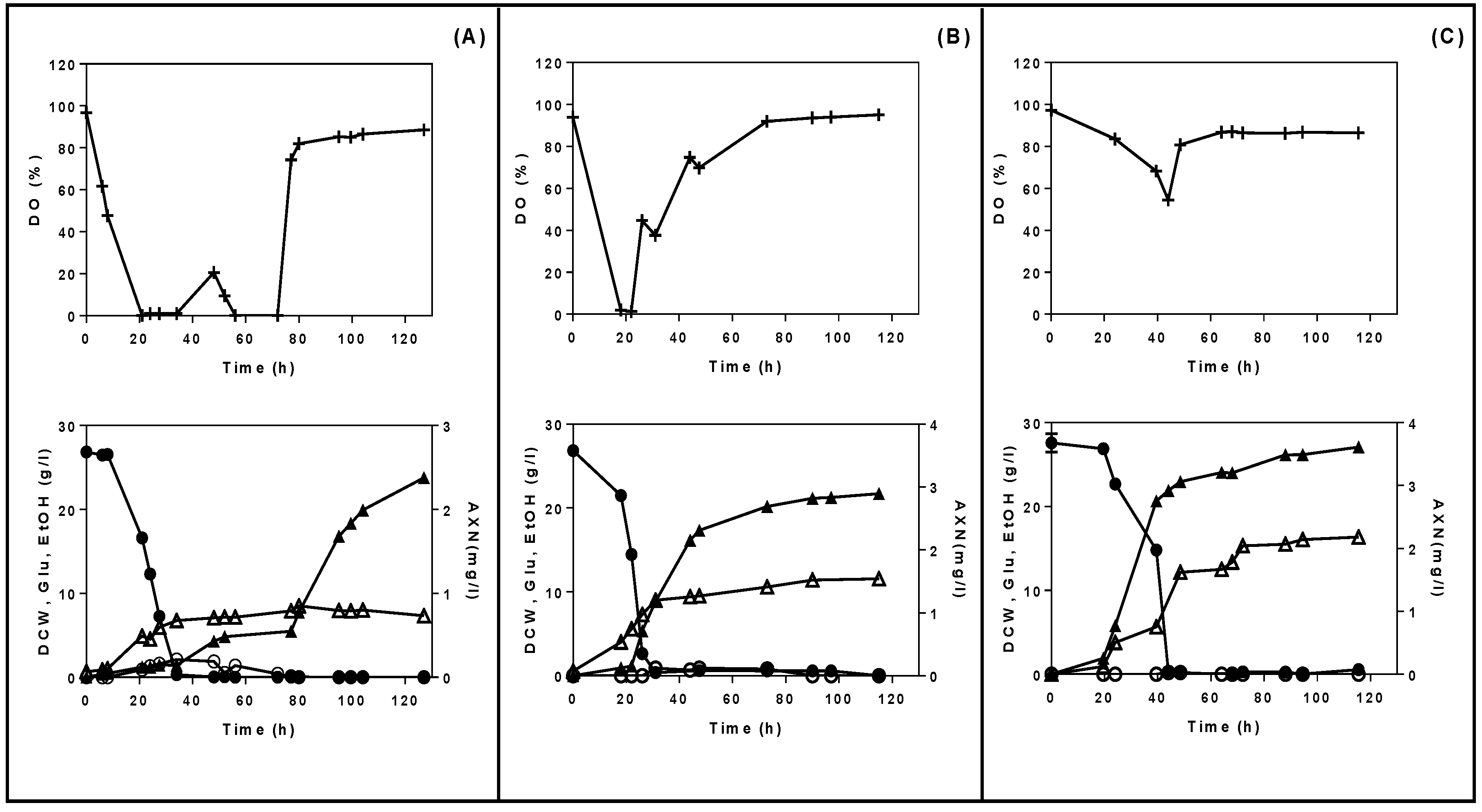
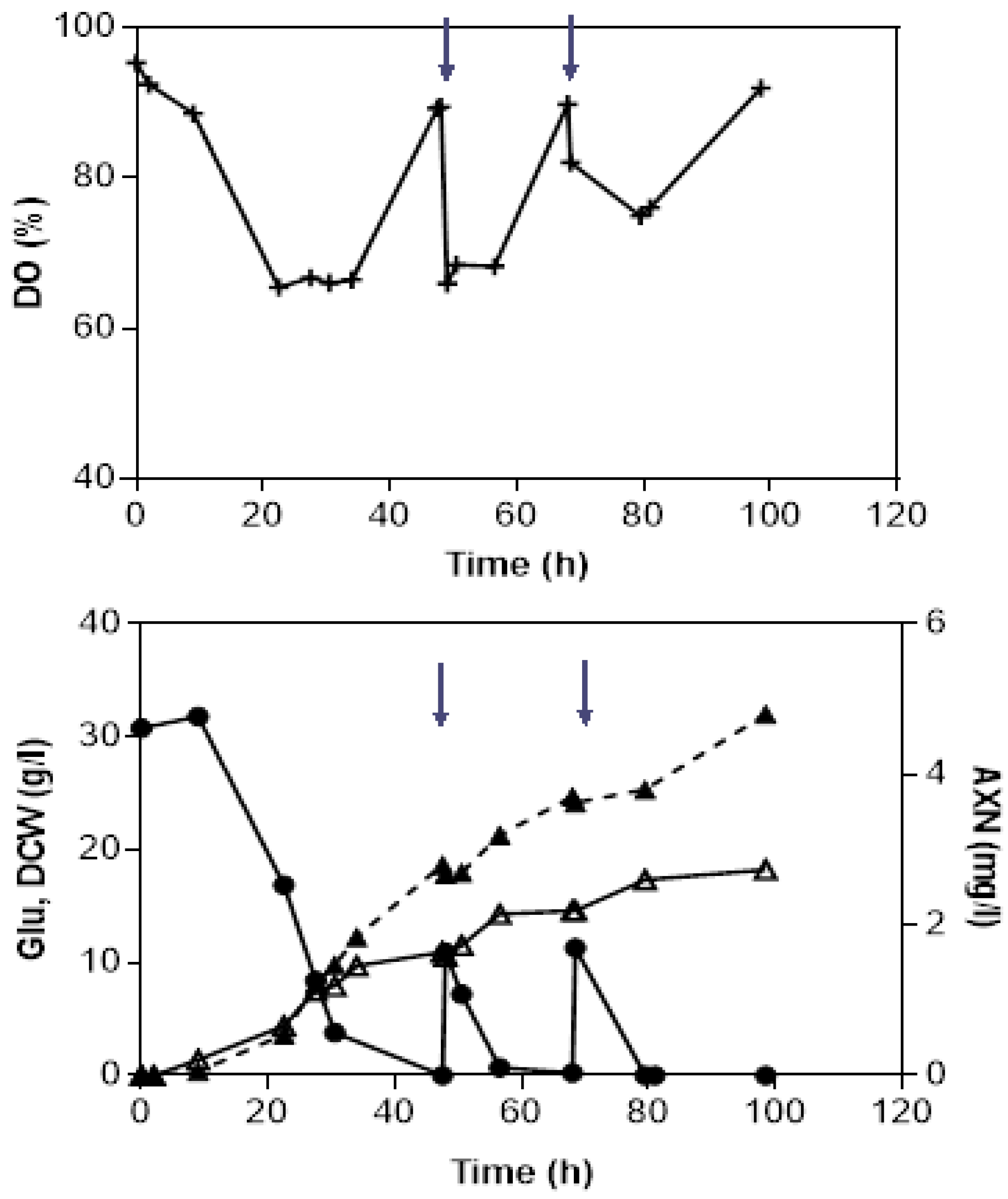
3.2. Fed-Batch Bioreactor Fermentations Using Semi-Defined Media
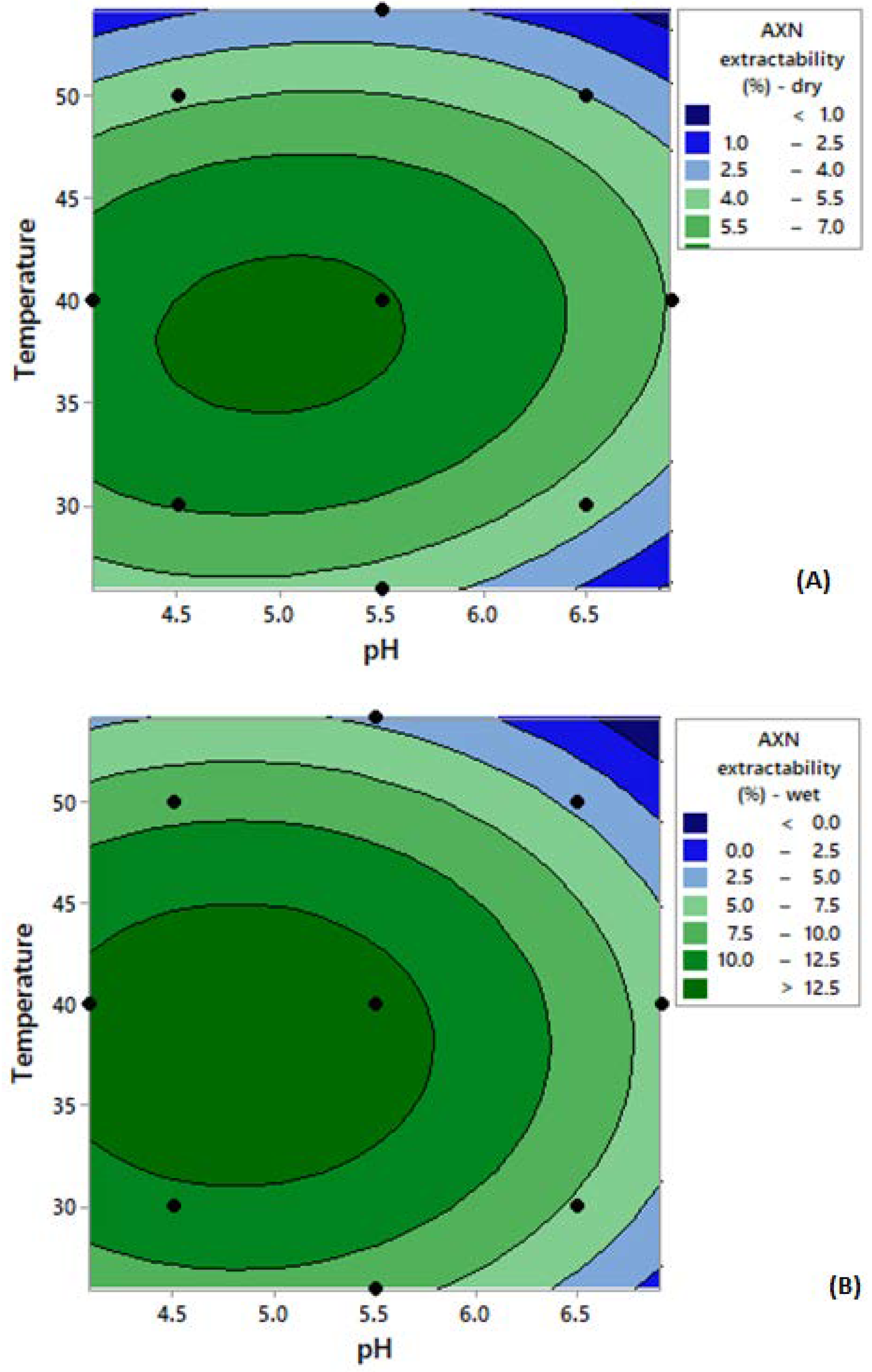
3.3. Enzymatic Cell Disruption and Astaxanthin Extraction
3.4. Supercritical Fluid Extraction as Means of Astaxanthin Extraction
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Molino, A.; Rimauro, J.; Casella, P.; Cerbone, A.; Larocca, V.; Chianese, S.; Karatza, D.; Mehariya, S.; Ferraro, A.; Hristoforou, E.; et al. Extraction of astaxanthin from microalga Haematococcus pluvialis in red phase by using generally recognized as safe solvents and accelerated extraction. J. Biotechnol. 2018, 283, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxantin-producing green microalga haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.C.; Zheng, Y.G.; Wang, Z.; Shen, Y.C. Production of astaxanthin by Xanthophyllomyces dendrorhous ZJUT46 with fed-batch fermentation in 2.0 M3 fermentor. Food Technol. Biotech. 2007, 45, 209–212. [Google Scholar]
- Monks, L.M.; Rigo, A.; Mazutti, M.A.; Vladimir Oliveira, J.; Valduga, E. Use of chemical, enzymatic and ultrasound-assisted methods for cell disruption to obtain carotenoids. Biocatal. Agric. Biotechnol. 2013, 2, 165–169. [Google Scholar] [CrossRef]
- Schmidt, I.; Schewe, H.; Gassel, S.; Jin, C.; Buckingham, J.; Hümbelin, M.; Sandman, G.; Schrader, J. Biotechnological production of astaxanthin with Phaffia rhodozyma/Xanthophyllomyces dendrorhous. Appl. Microbiol. Biotechnol. 2011, 89, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Meyer, P.S.; Du Preez, J.C. Effect of culture conditions on astaxanthin production by a mutant of Phaffia rhodozyma in batch and chemostat culture. Appl. Microbiol. Biotechnol. 1994, 40, 780–785. [Google Scholar] [CrossRef]
- Montanti, J.; Nghiem, N.P.; Johnston, D.B. Production of astaxanthin from cellulosic biomass sugars by mutants of the yeast Phaffia rhodozyma. Appl. Biochem. Biotech. 2011, 164, 655–665. [Google Scholar] [CrossRef]
- Valduga, E.; Rausch Ribeiro, A.H.; Cence, K.; Colet, R.; Tiggemann, L.; Zeni, J.; Toniazzo, G. Carotenoids production from a newly isolated Sporidiobolus pararoseus strain using agroindustrial substrates. Biocatal. Agric. Biotechnol. 2014, 3, 207–213. [Google Scholar] [CrossRef]
- Rodríguez-Sáiz, M.; de la Fuente, J.L.; Barredo, J.L. Xanthophyllomyces dendrorhous for the industrial production of astaxanthin. Appl. Microbiol. Biotechnol. 2010, 88, 645–658. [Google Scholar] [CrossRef]
- Yamane, Y.I.; Higashida, K.; Nakashimada, Y.; Kakizono, T.; Nishio, N. Influence of oxygen and glucose on primary metabolism and astaxanthin production by Phaffia rhodozyma in batch and fed-batch cultures: Kinetic and stoichiometric analysis. Appl. Environ. Microbiol. 1997, 63, 4471–4478. [Google Scholar] [CrossRef] [Green Version]
- Reynders, M.; Rawlings, D.E.; Harrision, S.T.L. Demonstration of the Crabtree effect in Phaffia rhodozyma during continuous and fed-batch cultivation. Biotechnol. Lett. 1997, 19, 549–552. [Google Scholar] [CrossRef]
- Yen, H.W.; Liu, Y.X.; Chang, J.S. The effects of feeding criteria on the growth of oleaginous yeast Rhodotorula glutinis in a pilot-scale airlift bioreactor. J. Taiwan Inst. Chem. Eng. 2015, 49, 67–71. [Google Scholar] [CrossRef]
- Flores-Cotera, L.B.; Martín, R.; Sánchez, S. Citrate, a possible precursor of astaxanthin in Phaffia rhodozyma: Influence of varying levels of ammonium, phosphate and citrate in a chemically defined medium. Appl. Microbiol. Biotechnol. 2001, 55, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, T.; Lababpour, A.; Shimahara, K.; Katoh, S. Astaxanthin production by Haematococcus pluvialis under illumination with LEDs. Enzym. Microb. Technol. 2004, 35, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Ip, P.F.; Wong, K.H.; Chen, F. Enhanced production of astaxanthin by the green microalga Chlorella zofingiensis in mixotrophic culture. Process Biochem. 2004, 39, 1761–1766. [Google Scholar] [CrossRef]
- Bhosale, P. Environmental and cultural stimulants in the production of carotenoids from microorganisms. Appl. Microbiol. Biotechnol. 2004, 63, 351–361. [Google Scholar] [CrossRef]
- Vázquez, M. Effect of the light on carotenoid profiles of Xanthophyllomyces dendrorhous strains (formerly Phaffia rhodozyma). Food Technol. Biotechnol. 2001, 39, 123–128. [Google Scholar]
- Stachowiak, B. Effect of illumination intensities on astaxanthin synthesis by Xanthophyllomyces dendrorhous and its mutants. Food Sci. Biotechnol. 2013, 22, 1033–1038. [Google Scholar] [CrossRef]
- de la Fuente, J.L.; Rodríguez-Sáiz, M.; Schleissner, C.; Díez, B.; Peiro, E.; Barredo, J.L. High-titer production of astaxanthin by the semi-industrial fermentation of Xanthophyllomyces dendrorhous. J. Biotechnol. 2010, 148, 144–146. [Google Scholar] [CrossRef]
- Sena, A.R.; Valasques Junior, G.L.; Baretto, I.K.S.P.; Assis, S.A. Application of Doehlert experimental design in the optimization of experimental variables for the Pseudozyma sp. (CCMB 306) and Pseudozyma sp. (CCMB 300) cell lysis. Ciência Tecnol. Alime 2012, 32, 762–767. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Ding, L.; Sun, J.; Boussetta, N.; Vorobiev, E. Yeast cell disruption strategies for recovery of intracellular bio-active compounds—A review. Innov. Food Sci. Emerg. 2016, 36, 181–192. [Google Scholar] [CrossRef]
- Chen, L.; Wang, J.L.; Ni, H.; Zhu, M.J. Disruption of Phaffia rhodozyma cells and preparation of microencapsulated astaxanthin with high water solubility. Food Sci. Biotechnol. 2019, 28, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Reyes, L.H.; Gomez, J.M.; Kao, K.C. Improving carotenoids production in yeast via adaptive laboratory evolution. Metab. Eng. 2014, 21, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Harith, Z.T.; Charalampopoulos, D.; Chatzifragkou, A. Rapeseed meal hydrolysate as substrate for microbial astaxanthin production. Biochem. Eng. J. 2019, 151, 107330. [Google Scholar] [CrossRef]
- Milic, T.V.; Rakin, M.; Siler-Marinkovic, S. Utilization of baker’s yeast (Saccharomyces cerevisiae) for the production of yeast extract: Effects of different enzymatic treatments on solid, protein and carbohydrate recovery. J. Serb. Chem. Soc. 2007, 5, 451–457. [Google Scholar] [CrossRef]
- Salazar, O. 2D PAGE: Sample Preparation and Fractionation. Methods in Molecular Biology; Posh, A., Ed.; Humana Press: Totowa, NJ, USA, 2008; Volume 424, pp. 23–34. [Google Scholar]
- Krichnavaruk, S.; Shotipruk, A.; Goto, M.; Pavasant, P. Supercritical carbon dioxide extraction of astaxanthin from Haematococcus pluvialis with vegetable oils as co-solvent. Bioresour. Technol. 2008, 99, 5556–5560. [Google Scholar] [CrossRef]
- Sedmak, J.J.; Weerasinghe, D.K.; Jolly, S.O. Extraction and quantification of astaxanthin from Phaffia rhodozyma. Biotechnol. Technol. 1990, 4, 107–112. [Google Scholar] [CrossRef]
- Lopes Brandão, R.; César, J.; Rosa, C.; Nicoli, J.R.; Vinicius, M.; Almeida, S.; Queiros, H.T.; Castro, I.M. Investigating acid stress response in different Saccharomyces strains. J. Mycol. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Yalcin, S.K.; Ozbas, Z.Y. Effects of pH and temperature on growth and glycerol production kinetics of two indigenous wine strains of Saccharomyces cerevisiae from Turkey. Braz. J. Microbiol. 2008, 39, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.-C.; Zheng, Y.-G.; Wang, Z.; Shen, Y.-C. pH control strategy in astaxanthin fermentation bioprocess by Xanthophyllomyces dendrorhous. Enzym. Microb. Technol. 2006, 39, 586–590. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Power, R. Relationship between pH and medium dissolved solids in terms of growth and metabolism of lactobacilli and Saccharomyces cerevisiae during ethanol production. Appl. Environ. Microb. 2005, 71, 2239–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valduga, E.; Schwartz, C.R.M.; Tatsch, P.O.; Tiggemann, L.; Di Luccio, M.; Treichel, H. Evaluation of aeration and substrate concentration on the production of carotenoids by Sporidiobolus salmonicolor (CBS 2636) in bioreactor. Eur. Food Res. Technol. 2011, 232, 453–462. [Google Scholar] [CrossRef]
- Luna-Flores, C.H.H.; Ramírez-Cordova, J.J.J.; Pelayo-Ortiz, C.; Femat, R.; Herrera-López, E.J.J. Batch and fed-batch modeling of carotenoids production by Xanthophyllomyces dendrorhous using Yucca fillifera date juice as substrate. Biochem. Eng. J. 2010, 53, 131–136. [Google Scholar] [CrossRef]
- Zheng, Y.-G.; Hu, Z.-C.; Wang, Z.; Shen, Y.-C. Large-Scale Production of Astaxanthin by Xanthophyllomyces dendrorhous. Food Bioprod. Process. 2006, 84, 164–166. [Google Scholar] [CrossRef]
- Rodrigues, F.; Ludovico, P.; Leão, C. Sugar metabolism in yeasts: An overview of aerobic and anaerobic glucose catabolism. In Biodiversity and Ecophysiology of Yeasts; Springer: Berlin/Heidelberg, Germany, 2006; pp. 101–121. [Google Scholar]
- Liu, Y.S.; Wu, J.Y. Modeling of Xanthophyllomyces dendrorhous growth on glucose and overflow metabolism in batch and fed-batch cultures for astaxanthin production. Biotechnol. Bioeng. 2008, 101, 996–1004. [Google Scholar] [CrossRef]
- Moriel, D.G.; Chociai, M.B.; Machado, I.M.P.; Fontana, J.D.; Bonfim, T.M.B. Effect of feeding methods on the astaxanthin production by Phaffia rhodozyma in fed-batch process. Braz. Arch. Biol. Technol. 2005, 48, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, M.; Martin, A.M. Optimization of continuous culture through response surface methodology. Biotechnol. Bioeng. 1998, 57, 314–320. [Google Scholar] [CrossRef]
- Rezaei, K.; Jenab, E.; Temelli, F. Effects of water on enzyme performance with an emphasis on the reactions in supercritical fluids. Crit. Rev. Biotechnol. 2007, 27, 183–195. [Google Scholar] [CrossRef]
- Michelon, M.; De Matos De Borba, T.; Da, R.; da Silva Rafael, R.; Burkert, C.A.V.; Burkert, J.F.M. Extraction of carotenoids from Phaffia rhodozyma: A comparison between different techniques of cell disruption. Food Sci. Biotechnol. 2012, 21, 1–8. [Google Scholar] [CrossRef]
- Machado, F.R.S.; Trevisol, T.C.; Boschetto, D.L.; Burkert, J.F.M.; Ferreira, S.R.S.; Oliveira, J.V.; Burkert, C.A.V. Technological process for cell disruption, extraction and encapsulation of astaxanthin from Haematococcus pluvialis. J. Biotechnol. 2016, 218, 108–114. [Google Scholar] [CrossRef] [Green Version]
- de Andrade Lima, M.; Charalampopoulos, D.; Chatzifragkou, A. Purification of supercritical-fluid carotenoid-rich extracts by hydrophobic interaction chromatography. Sep. Purif. Technol. 2018, 203, 1–10. [Google Scholar] [CrossRef]
- Lim, G.-B.; Lee, S.-Y.; Lee, E.-K.; Haam, S.-J.; Kim, W.-S. Separation of astaxanthin from red yeast Phaffia rhodozyma by supercritical carbon dioxide extraction. Biochem. Eng. J. 2002, 11, 181–187. [Google Scholar] [CrossRef]
- Duarte, S.H.; dos Santos, P.; Michelon, M.; de Oliveira, S.M.; Martínez, J.; Maugeri, F. Recovery of yeast lipids using different cell disruption techniques and supercritical CO2 extraction. Biochem. Eng. J. 2017, 125, 230–237. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | pH | Temperature (°C) | Astaxanthin Extractability (%, w/w) | |
|---|---|---|---|---|
| Dry Cells | Wet Cells | |||
| 1 | 4.50 (−1) | 30.0 (−1) | 6.33 | 10.59 |
| 2 | 6.50 (+1) | 30.0 (−1) | 4.91 | 6.98 |
| 3 | 4.50 (−1) | 50.0 (+1) | 4.84 | 8.85 |
| 4 | 6.50 (+1) | 50.0 (+1) | 4.78 | 5.30 |
| 5 | 4.09 (−1) | 40.0 (0) | 8.66 | 14.08 |
| 6 | 6.91 (+1) | 40.0 (0) | 4.65 | 5.81 |
| 7 | 5.50 (0) | 25.7 (−1) | 4.97 | 9.30 |
| 8 | 5.50 (0) | 54.1 (+1) | 2.52 | 4.00 |
| 9 | 5.50 (0) | 40.0 (0) | 8.91 | 12.60 |
| 10 | 5.50 (0) | 40.0 (0) | 9.12 | 13.50 |
| 11 | 5.50 (0) | 40.0 (0) | 7.49 | 13.11 |
| 12 | 5.50 (0) | 40.0 (0) | 7.95 | 12.95 |
| 13 | 5.50 (0) | 40.0 (0) | 9.43 | 13.76 |
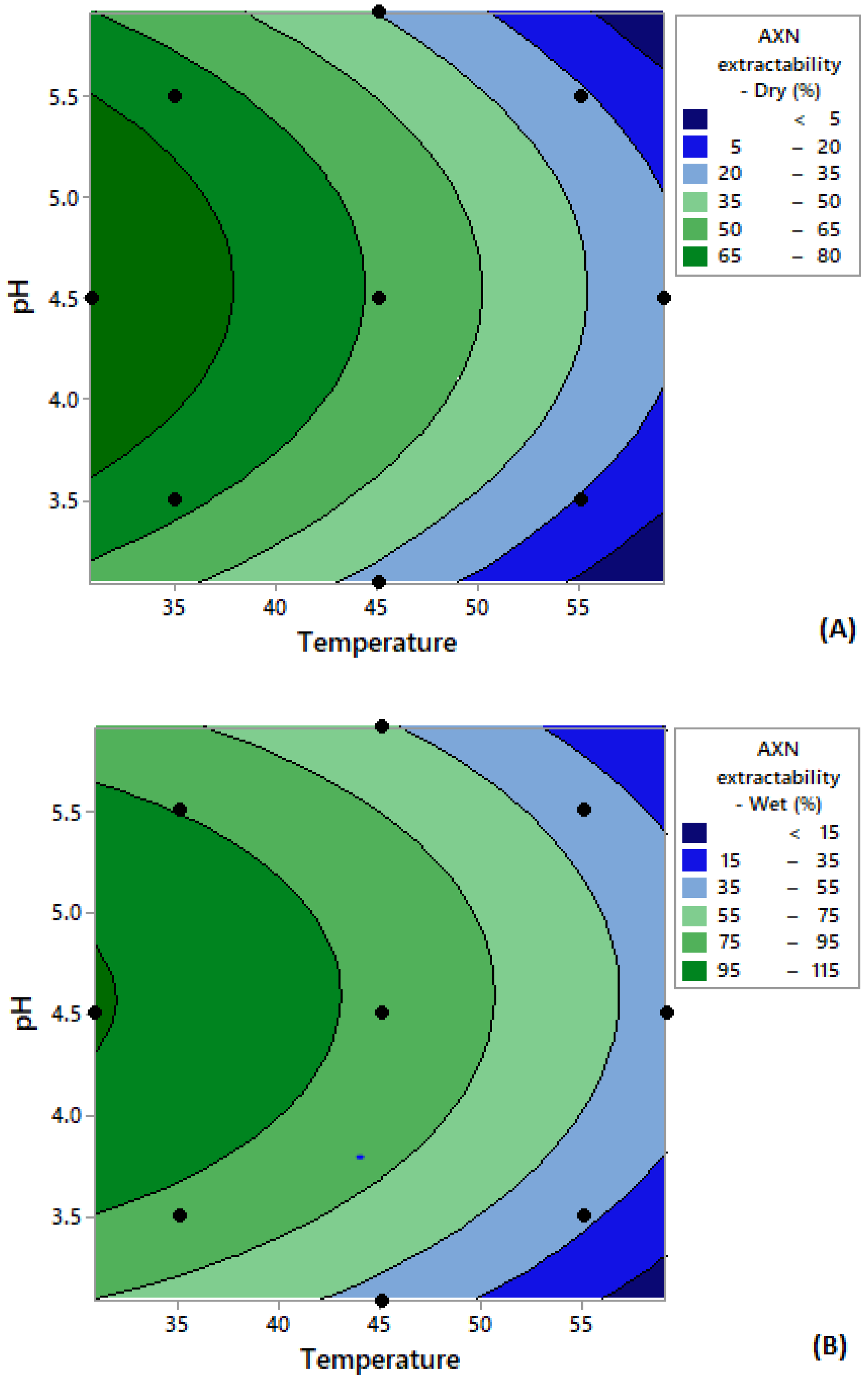
| Run | pH | Temperature (°C) | Astaxanthin Extractability (%, w/w) | |
|---|---|---|---|---|
| Wet Cells | Dry Cells | |||
| 1 | 3.5 (−1) | 35.0 (−1) | 91.80 | 79.80 |
| 2 | 3.5 (−1) | 55.0 (+1) | 39.95 | 22.04 |
| 3 | 5.50 (+1) | 35.0 (−1) | 93.33 | 76.91 |
| 4 | 5.50 (+1) | 55.0 (+1) | 43.00 | 18.09 |
| 5 | 4.5 (0) | 30.9 (−1) | 114.83 | 84.36 |
| 6 | 4.5 (0) | 59.1 (+1) | 47.58 | 25.99 |
| 7 | 3.09 (−1) | 45.0 (0) | 44.83 | 22.04 |
| 8 | 5.91 (+1) | 45.0 (0) | 60.24 | 35.57 |
| 9 | 4.50 (0) | 45.0 (0) | 88.91 | 61.41 |
| 10 | 4.50 (0) | 45.0 (0) | 90.89 | 66.73 |
| 11 | 4.50 (0) | 45.0 (0) | 95.16 | 63.23 |
| 12 | 4.50 (0) | 45.0 (0) | 84.94 | 61.26 |
| 13 | 4.50 (0) | 45.0 (0) | 90.89 | 65.36 |
| Run | Temperature (°C) | Pressure (bar) | EtOH (%) | Astaxanthin Extraction (%, w/w) |
|---|---|---|---|---|
| 01 | 50.0 (−1) | 150.0 (−1) | 5.0 (−1) | 7.78 |
| 02 | 70.0 (+1) | 150.0 (−1) | 5.0 (−1) | 1.62 |
| 03 | 50.0 (−1) | 350.0 (+1) | 5.0 (−1) | 16.37 |
| 04 | 70.0 (+1) | 350.0 (+1) | 5.0 (−1) | 17.94 |
| 05 | 50.0 (−1) | 150.0 (−1) | 15.0 (+1) | 20.33 |
| 06 | 70.0 (+1) | 150.0 (−1) | 15.0 (+1) | 14.50 |
| 07 | 50.0 (−1) | 350.0 (+1) | 15.0 (+1) | 21.70 |
| 08 | 70.0 (+1) | 350.0 (+1) | 15.0 (+1) | 23.01 |
| 09 | 43.2 (−1) | 250.0 (0) | 10.0 (0) | 13.40 |
| 10 | 76.8 (+1) | 250.0 (0) | 10.0 (0) | 14.82 |
| 11 | 60.0 (0) | 81.8 (−1) | 10.0 (0) | 0.04 |
| 12 | 60.0 (0) | 418.2 (+1) | 10.0 (0) | 13.38 |
| 13 | 60.0 (0) | 250.0 (0) | 1.6 (0) | 4.24 |
| 14 | 60.0 (0) | 250.0 (0) | 18.4 (+1) | 19.64 |
| 15 | 60.0 (0) | 250.0 (0) | 10.0 (0) | 14.30 |
| 16 | 60.0 (0) | 250.0 (0) | 10.0 (0) | 14.32 |
| 17 | 60.0 (0) | 250.0 (0) | 10.0 (0) | 16.52 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harith, Z.T.; de Andrade Lima, M.; Charalampopoulos, D.; Chatzifragkou, A. Optimised Production and Extraction of Astaxanthin from the Yeast Xanthophyllomyces dendrorhous. Microorganisms 2020, 8, 430. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030430
Harith ZT, de Andrade Lima M, Charalampopoulos D, Chatzifragkou A. Optimised Production and Extraction of Astaxanthin from the Yeast Xanthophyllomyces dendrorhous. Microorganisms. 2020; 8(3):430. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030430
Chicago/Turabian StyleHarith, Zuharlida Tuan, Micael de Andrade Lima, Dimitris Charalampopoulos, and Afroditi Chatzifragkou. 2020. "Optimised Production and Extraction of Astaxanthin from the Yeast Xanthophyllomyces dendrorhous" Microorganisms 8, no. 3: 430. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms8030430